組織激肽釋放酶家族在病原微生物感染中的作用
2024-01-29 07:53:52尚雪恬潘麗萍
中國防癆雜志 2024年2期
關鍵詞:研究
尚雪恬 潘麗萍
組織激肽釋放酶(tissue kallikrein-related peptidase, KLK)家族廣泛分布于皮膚、胰腺、腎臟、肺臟、心血管、腎上腺和前列腺等組織[1]。一直以來,KLK在心血管、癌癥等慢性病領域中研究較多,而在病原微生物感染領域知之甚少。近年來,逐漸有研究發現KLK可以通過切割病原體表面關鍵蛋白、激活多種宿主受體、調控激肽系統等方式參與機體多種生理活動,在病毒、細菌、真菌感染中發揮重要作用,影響病原微生物感染的進程[2-3]。但是,盡管KLK家族蛋白在各類微生物感染中有一定的研究,而且也發現部分家族蛋白在結核感染和結核病發病中發揮作用,但迄今為止,并無該家族在感染性疾病及結核病領域方面的總結研究。因此,筆者通過梳理KLK家族蛋白在感染性疾病發病中或宿主抗感染過程中的具體作用及功能機制,對KLK家族在感染性疾病領域的研究現狀進行總結,這將為未來在結核病領域更廣泛地開展針對于KLK家族蛋白的功能和機制研究提供一定的研究思路和方向。
一、KLK家族及生物學活性
1.KLK家族:KLK家族是絲氨酸蛋白酶家族中一種具有胰蛋白酶或胰凝乳蛋白酶性質的蛋白水解酶亞群,由15個保守基因(KLK1~KLK15)編碼的分泌絲氨酸蛋白酶組成,編碼基因均位于19q13.4區域,是人類基因組中最大的蛋白酶基因簇[4]。激肽釋放酶這一概念最初用于描述人胰腺中能產生激肽的物質,此后又陸續鑒定出10余種激肽釋放酶相關肽酶[4],目前主要分為血漿激肽釋放酶(plasma kallikrein, PK)和組織激肽釋放酶家族(tissue kallikrein,TK)[3]兩大類。KLK家族不僅廣泛分布于各種組織中,也表達于多種免疫細胞(如中性粒細胞)、肺巨噬細胞,以及血漿、尿液、唾液、淚液和汗液等多種體液中[5-7]。
2.KLK生物學活性:所有的KLK蛋白都是以前肽的形式合成的,分泌后進行蛋白質水解處理,去除前肽,轉化為具有酶活性的成熟形式,并釋放到血液中,其活性可以被抑肽酶和激肽酶相關肽酶抑制劑抑制[4]。近些年發現,KLK可以水解多種病毒和細菌的關鍵蛋白。由于微生物感染機制的不同,在病毒感染中多表現為促進病毒感染,如流感病毒;在細菌感染中則表現為干擾細菌的定植,如奈瑟菌[8-9];在結核分枝桿菌感染中,KLK可以通過調節自噬、凋亡和促炎癥因子釋放發揮其抗菌作用。KLK是激肽釋放酶-激肽系統(kallikrein-kinin system, KKS)的重要成員,可以水解激肽原產生激肽,而激肽是一種具有促炎活性的生物活性肽,可以誘導不同類型的細胞釋放前列腺素,誘導內皮細胞釋放一氧化氮(NO)[10]。人中性粒細胞可以分泌KLK1、KLK6、KLK10、KLK13和KLK14,這些激肽釋放酶可能通過水解組織中的激肽原產生激肽而維持炎癥狀態[6]。KLK除了可以調控激肽釋放酶-激肽系統外,還可以激活肝細胞生長因子(HGF)和尿激酶纖維蛋白溶酶原激活劑受體(urokinase plasminogen activator receptor, uPAR)等分子來促進肺結核患者肺部炎癥[11]。
二、KLK與病原微生物感染
由于KLK的酶活性作用可以水解多種蛋白,因此,能夠參與包括病毒、細菌和真菌在內的多種病原微生物的感染過程。但是,目前的研究提示在不同微生物感染過程中KLK發揮的作用及機制均有所不同(表1),現筆者參照功能分類闡述KLK在不同病原微生物感染中的作用機制并進行總結歸類,為KLK在結核分枝桿菌感染中的機制研究提供思路。
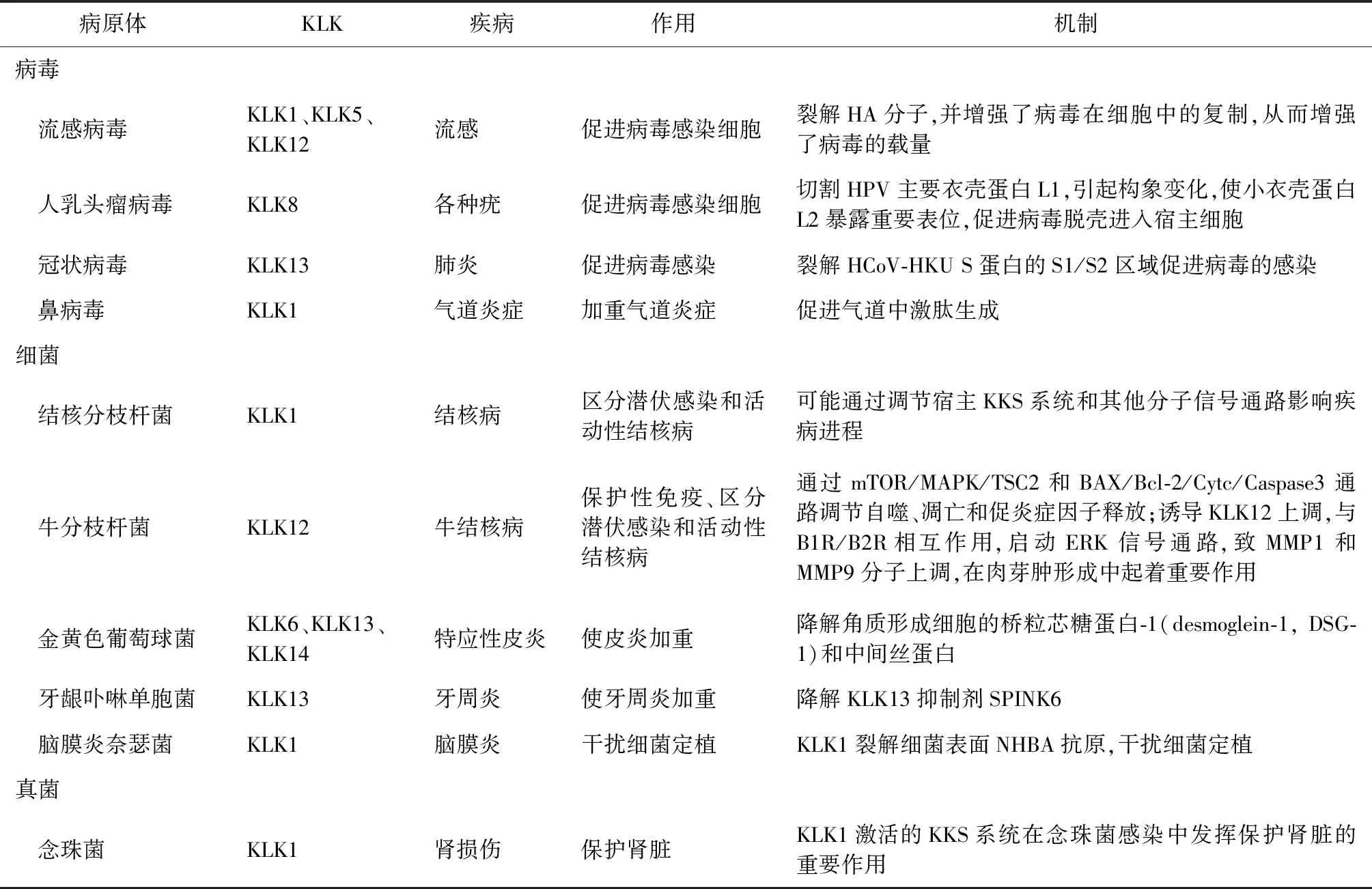
表1 KLK家族基因在不同病原微生物感染中的作用
(一)KLK可水解微生物表面關鍵蛋白
很多病毒需要通過表面關鍵蛋白裂解活化后才能感染宿主細胞。在流感病毒感染宿主細胞過程中血凝素(hemagglutinin, HA)介導的與細胞表面受體結合及膜融合是不可或缺的步驟[12]。流感病毒的血凝素被絲氨酸蛋白酶水解激活后才具有感染性。Magnen等[9]發現流感病毒感染體外重組的人支氣管上皮細胞后,會引發KLK1和KLK5表達增加,因此,流感發病期患者的下呼吸道分泌物中KLK1和KLK5含量增加,其絲氨酸蛋白酶活性能夠水解激活流感病毒血凝素,進一步促進病毒感染;同時還發現,使用KLK5選擇性抑制劑預處理氣道分泌物并與病毒粒子共同孵育,可顯著降低流感病毒的激活,從而抑制病毒侵染細胞。另有研究發現,在流感期間小鼠肺部的KLK1分泌增加,而組織激肽釋放酶結合蛋白水平下降,KLK1裂解血凝素分子,并增強病毒在細胞中的復制,從而增強病毒的載量;相反,酶結合蛋白通過抑制KLK1介導的血凝素裂解可以減少病毒的產生[13]。有研究表明,不同KLK對不同亞型的流感病毒的蛋白切割有選擇性,如KLK5切割流感病毒蛋白H1和H3最有效,而KLK12切割H1和H2最有效;另外,纖溶酶、尿激酶和前激酶不僅是活性胰蛋白酶樣蛋白酶的前體,還可以激活流感病毒;在流感病毒感染后,這些酶原集中在呼吸道組織損傷的部位,也可能被KLK5和KLK12激活,進一步促進流感病毒的感染[14]。人乳頭瘤病毒(human papillomavirus, HPV)衣殼蛋白L1、L2構象改變在病毒入侵細胞的過程中十分關鍵。KLK8可以介導切割HPV主要衣殼蛋白L1,引起構象變化,使小衣殼蛋白L2暴露重要表位,促進病毒脫殼進入宿主細胞[15]。但另有研究發現,親環素和弗林蛋白酶介導的結構修飾可以使L2的N端暴露,導致病毒內化的時間縮短,使結合到細胞表面的病毒快速內化;而KLK8裂解引起的構象變化則對病毒的內化沒有影響,使其在組織細胞感染過程中的切割效率較低,不能使病毒更同步或更快速地進入細胞[16],故KLK8是否促進HPV病毒的感染還需進一步研究。Milewska等[17]使用冠狀病毒HCo-HKU1感染人氣道上皮細胞后,發現KLK13的表達增加,而使用化學方法或短發夾RNA(shRNA)將KLK13阻斷后則可使病毒感染減少;同時,他們還證實KLK13裂解HCoV-HKU S蛋白的S1/S2區域可促進病毒的感染。
KLK的絲氨酸酶活性也可以水解細菌表面抗原,干擾細菌感染。研究發現,唾液中的KLK1可以裂解腦膜炎奈瑟菌表面的奈瑟菌肝素結合抗原(neisserial heparin binding antigen, NHBA),該抗原是一種脂質抗原,不僅可以促進細菌在人血清中的存活,還可能在腦膜炎奈瑟菌定植的黏附步驟中發揮關鍵作用[8]。
雖然目前尚未有KLK裂解結核分枝桿菌表面蛋白或分泌蛋白進而調控結核分枝桿菌入侵的報道,但如同腦膜炎奈瑟菌感染中KLK1可以裂解脂質抗原NHBA一樣,結核分枝桿菌表面也有很多其他的脂質抗原,所以并不能排除KLK也可能通過裂解結核分枝桿菌表面脂質抗原來影響結核分枝桿菌的入侵。除了裂解病原微生物蛋白之外,KLK家族蛋白還可能通過調控宿主其他生理和病理作用來影響結核病的發生和發展。
(二)KLK通過激肽釋放酶-激肽系統影響感染進程
激肽是由激肽原經激肽釋放酶的作用而產生,通過激肽酶水解而失活。這一過程中參與的各種因子組成了激肽釋放酶-激肽系統。激肽釋放酶-激肽系統包括激肽釋放酶、激肽原、激肽(緩激肽和胰激肽)、激肽受體及激肽酶。目前已經發現激肽受體和激肽系統的其他成分參與了諸多感染性疾病,包括新型冠狀病毒感染、念珠菌病和結核病等。
研究發現,下呼吸道速發超敏反應與激肽釋放酶-激肽系統密切相關,緩激肽(bradykinin, BK)及其受體參與了其中一系列的病理改變。在實驗性感染鼻病毒的受試者中,上呼吸道癥狀的嚴重程度與鼻分泌物中的激肽生成水平直接相關,鼻病毒誘導KLK1激活可能導致氣道炎癥和哮喘加重[18-19]。研究表明,新型冠狀病毒感染患者可能存在緩激肽相關通路的失調,緩激肽可能引發新型冠狀病毒感染患者的細胞因子風暴和肺水腫[20-21],阻斷激肽系統可以預防患者出現急性呼吸窘迫綜合征,并且通過阻斷激肽系統中KLK的產生、降解激肽、下調激肽受體等不同環節來減少或降低新型冠狀病毒感染的相關癥狀和疾病惡化[2]。此外,Ramani等[22-23]發現白細胞介素17(IL-17)在白色念珠菌感染的腎臟中誘導KLK1表達,導致腎小管上皮細胞高表達KLK1,通過激肽釋放酶-激肽系統產生緩激肽,并作用于緩激肽受體B1R和B2R,通過限制腎臟駐留細胞的凋亡來保護全身真菌感染期間的腎功能,防止播散性念珠菌感染后免疫活性小鼠的腎損傷;同時在小鼠實驗中,過表達KLK1和使用緩激肽治療可以使IL-17-/-小鼠感染念珠菌后存活時間延長;而使用抑制劑阻斷野生型小鼠的緩激肽受體B1R和B2R后則存活時間縮短。由此可見,KLK1激活的激肽釋放酶-激肽系統在念珠菌感染中發揮著保護腎臟的重要作用;同時基于KLK1受到IL-17的調控,側面說明了KLK1與炎癥反應相關。如前所述,激肽的激活在呼吸道感染中會加重炎癥和呼吸道疾病,在肺結核患者中激肽釋放酶-激肽系統是否也會加重患者的臨床癥狀將有待進一步研究。
雖然目前關于KLK家族蛋白在結核病發生發展中的作用機制研究相對較少,但已有報道發現與KLK家族蛋白功能相關的激肽釋放酶-激肽系統在結核病中發揮重要作用。隨著患者從預處理到治療早期、延長給藥階段和治療結束這一周期內,活動性結核病患者循環水平的緩激肽和脫精氨酸9緩激肽(des-Arg9-BK)的濃度均下降,二者可以誘導炎癥和先天性免疫反應[24]。在結核分枝桿菌感染的小鼠脾臟和肺臟中緩激肽受體 B1R均上調,應用B1R阻斷劑對菌落形成單位(CFU)計數有明顯抑制作用[25]。由此可推測,激肽釋放酶-激肽系統與結核分枝桿菌的感染活動有關。此外,有研究發現KLK1基因可能作為活動性結核病患者與結核分枝桿菌潛伏感染者鑒別診斷的生物標識[26]。這些研究均提示KLK1作為激肽釋放酶-激肽系統的關鍵分子,可能通過該系統調控體內結核分枝桿菌的增殖,但其中涉及的具體分子及機制仍有待進一步研究。
(三)KLK通過激活其他宿主蛋白影響感染進程
KLK除了參與激肽釋放酶-激肽系統外,還可以激活其他宿主蛋白。研究發現,KLK4和KLK5可以激活肝細胞生長因子激活因子(hepatocyte growth factor activator, HGFA)。HGFA是一種絲氨酸蛋白酶和原肝細胞生長因子/分散因子(pro-HGF/SF)的有效激活劑,因此,KLK可以間接激活肝細胞生長因子[27],參與肺部炎癥。有研究發現,活動性肺結核患者血清肝細胞生長因子水平升高,疾病穩定后血清肝細胞生長因子水平降低[28]。He等[29]采用Luminex多因子檢測技術分析活動性結核病、結核分枝桿菌潛伏感染和健康對照組中受早期分泌抗原靶6(ESAT-6)蛋白刺激的外周血單個核細胞分泌的細胞因子,發現肝細胞生長因子可以有效區分結核分枝桿菌潛伏感染與健康對照組。有研究對比分析發現肺軍團菌病、肺結核及其他細菌感染患者的血清肝細胞生長因子水平均高于健康對照組[30]。也有研究表明,KLK可以激活uPAR,而血清可溶性uPAR在活動性結核病患者中的表達水平升高,在患者治療效果較好時降低,因此,認為血清可溶性uPAR水平可以用于指導結核病的治療管理[31]。Araujo等[32]研究也發現,抗肽P-12034 IgG和uPAR聯合檢測可用于鑒別結核病患者。這說明KLK可能在結核病發病過程中通過這些受體發揮作用,但對于KLK是如何通過各種受體調節機體對結核分枝桿菌感染免疫應答的具體過程和機制還需要進一步深入探討。
作為蛋白酶,KLK成員還可以降解其他宿主蛋白。在金黃色葡萄球菌引發的特應性皮炎中,金黃色葡萄球菌能夠刺激人膠質細胞,增加其內源性蛋白酶的活性,同時能夠誘導KLK6、KLK13和KLK14表達,降解角質形成細胞的橋粒芯糖蛋白-1(desmoglein-1, DSG-1)和中間絲蛋白,使皮炎加重[33]。橋粒形成受損會導致肺上皮屏障功能受損[34],因此,KLK是否通過降解各種連接蛋白破壞肺上皮細胞的屏障作用來加重肺結核患者肺部病理變化值得進一步研究。此外,病原蛋白與宿主蛋白相互作用是感染的重要介質。有研究發現,牙齦卟啉單胞菌分泌的牙齦菌蛋白酶(gingipains)既能降解KLK13抑制劑SPINK6,又能激活KLK13蛋白,加重對組織的損傷,加重牙周炎[35]。因此,基于結核分枝桿菌也可分泌多種蛋白和酶類[36],也可認為其有可能通過與宿主蛋白之間的相互作用起到激活或抑制KLK活性的作用,進而影響感染進程。
(四)KLK通過調控各類信號通路影響感染進程
Sabir等[37]研究發現,KLK12在牛分枝桿菌感染的小鼠原代巨噬細胞中上調,可通過mTOR/MAPK/TSC2和BAX/Bcl-2/Cytc/Caspase3通路調節自噬、凋亡和促炎癥因子的釋放,在保護性免疫應答調節中發揮重要作用;同時,牛分枝桿菌特異性抗原ManLAM也可誘導感染后的巨噬細胞內的KLK12上調,并與緩激肽受體B1R/B2R相互作用,啟動ERK信號通路,導致基質金屬蛋白酶MMP1和MMP9分子的表達上調,在肉芽腫形成中起著重要作用。另外,KLK12在牛結核病和牛結核分枝桿菌潛伏感染模型中差異表達,可用作區分潛伏性和活動性牛結核病的血清學標記物[38]。研究發現,KLK家族蛋白參與多種信號通路[39],包括KLK4可以調控ERK/AKT信號通路[40]、KLK8可以調控MAPK信號通路[41]、KLK3可以調控Wnt信號通路等[42]。盡管目前尚未有報道關于上述KLK家族蛋白與結核病發生發展的相關研究,但這些信號通路均與結核病的發生發展密切相關[43],因此,KLK家族蛋白在結核病的發生發展過程中是否通過上述信號,抑或通過其他信號通路影響宿主內結核分枝桿菌的存活及疾病進程,仍需要進一步研究。
三、總結和展望
盡管目前的研究已經發現KLK可以通過切割病毒蛋白促進病毒感染、調控經典的激肽釋放酶-激肽系統,或調控宿主特定蛋白、影響信號通路等方式參與到病毒、細菌、真菌等微生物感染后的疾病進程中,但具體的分子、機制、調控通路等并未能完全清晰闡釋。此外,現有數據表明,在不同微生物感染中KLK家族蛋白發揮的作用可能有所差別,但具體差別如何尚需要更深入和廣泛的研究。另外,既往研究發現的這些KLK家族基因,是否可以作為微生物感染的生物標志物,通過靶向這些KLK是否能夠發揮治療作用也未能確定。再者,KLK并非獨立存在,可能是作為復雜蛋白酶網絡的一部分在發揮作用,而我們對于調控這些KLK的復雜的蛋白網絡的理解并不完整。特別是在結核感染和結核病研究領域,目前僅發現KLK1和KLK12具有鑒別診斷結核病和結核分枝桿菌潛伏感染的潛力,且針對上述兩個分子在結核病發病中的作用和機制解析尚不夠深入,對KLK家族其他分子是否具備作為結核病診斷或治療靶標的潛力,其作用機制如何,也需要進一步廣泛研究。
筆者通過對KLK家族蛋白既有研究的梳理和總結,展現了KLK作用機制的多樣性與復雜性,同時也提示該家族在病原微生物感染領域中可能存在較好的研究潛質,對KLK家族蛋白功能及作用機制的深入研究,可能為后續研發靶向KLK的新型抗感染藥物提供較好的理論基礎。因此,未來應在感染領域加深對KLK家族基因功能和機制的相關研究,以明確其在病原微生物感染中的具體作用并指導臨床應用。
利益沖突所有作者均聲明不存在利益沖突
作者貢獻尚雪恬:查閱文獻、撰寫初稿;潘麗萍:文獻審核、修改文稿、基金支持
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
