寄生蟲抗藥性及其對水產動物寄生蟲病藥物防治的啟示
2024-02-07 07:30:00李文祥習丙文王桂堂
水生生物學報 2024年2期
關鍵詞:機制
李文祥 王 浩 習丙文 李 明 聶 品, 4 王桂堂
(1.中國科學院水生生物研究所淡水生態與生物技術國家重點實驗室, 武漢 430072; 2.上海海洋大學, 上海 201306;3.中國水產科學研究院淡水漁業研究中心, 無錫 214081; 4.青島農業大學, 青島 266109)
為減少細菌性疾病的發生, 抗生素被大量用于陸生動物[1]和水生動物[2,3] 的養殖生產, 從而引起細菌對抗生素產生了抗性(Antimicrobial resistance),這不僅影響養殖動物疾病的藥物防控效果, 而且還會使人類相關疾病的病原菌產生抗藥性[4]。細菌的抗藥性(Drug resistance)已成為全球關注的熱點問題[5]。
抗藥性原本是指微生物能經受住藥物毒性的一種能力, 少數個體在藥物作用下發生遺傳變異,并在持續的藥物選擇壓力下存活, 從而形成對藥物具有耐受力的種群[6,7]; 且抗藥能力取決于抗性個體在種群中的比例和抗藥機制[8]。目前陸生動物寄生蟲病的防治仍然依靠化學藥物, 由于驅殺蟲藥物(Anthelmintics)的長期大量使用, 寄生蟲產生了嚴重的抗藥性[9]。抗寄生蟲藥物的種類本來就很有限, 隨著抗藥性的出現, 可用的藥物越來越少, 且短期內有效的疫苗難以研制出來, 加之新藥研制是個漫長的過程, 造成了當前寄生蟲病的藥物防治越來越困難[10], 寄生蟲抗藥性受到廣泛關注[11—15]。
與陸生動物相比, 水生動物寄生蟲的抗藥性研究較少[16], 雖然在20世紀90年代發現了歐洲鰻鱺(Anguilla anguilla)的擬指環蟲(Pseudodactylogyrusspp.)的抗藥性問題[17], 但針對抗藥性的系統研究目前僅限于鮭鱒類寄生的鮭瘡痂魚虱(Lepeophtheirus salmonis)[18—20]。高密度養殖中殺蟲藥物的反復使用是產生抗藥性的前提條件[16], 隨著我國水產養殖集約化程度逐漸提高和養殖規模逐漸擴大, 寄生蟲病的發生越來越嚴重, 殺蟲藥物被大量使用, 有些寄生蟲病的防治效果已呈現明顯下降趨勢, 表明寄生蟲可能已經產生抗藥性, 然而缺乏相關研究。
本文將通過對寄生蟲病主要防治藥物、寄生蟲抗藥性機制、抗藥性測定方法、減緩抗藥性措施及水產養殖動物寄生蟲抗藥性的研究現狀進行綜述, 為我國水產養殖中寄生蟲抗藥性研究提供思路, 同時為寄生蟲病的合理用藥提供策略, 避免或減緩抗藥性的產生。
1 寄生蟲病主要防治藥物及驅殺蟲的作用機制
世界上幾乎所有的殺蟲藥物都是為了防治農作物害蟲(主要是昆蟲和螨)、家禽家畜的寄生蟲(球蟲、吸蟲、絳蟲和線蟲等)和人體寄生蟲而研發的。目前使用的抗寄生蟲藥物主要涵蓋以下幾類:有機磷類(Organophosphates)、擬除蟲菊酯類(Pyrethroids)、大環內酯類(Macrocyclic lactone)、苯甲酰脲類(Benzoyl ureas)、新煙堿類(Neonicotinoid)、咪唑并噻唑類(Imidazothiazole)、苯并咪唑類(Benzimidazoles)和異喹啉類(Isoquinoline), 還有一些化學合成抗球蟲藥, 以及驅蟲殺蟲的無機化合物、消毒劑和中草藥等。
寄生蟲的抗藥性與藥物種類和作用方式(Mode of action)密切相關, 以下是幾類主要殺蟲藥物的作用方式或作用機制。
1.1 有機磷類
有機磷類是應用最為廣泛的一類殺蟲劑, 以胃毒、觸殺、熏蒸或內吸等多種方式作用于蟲害[21]。該類藥物的主要靶標為乙酰膽堿酯酶, 可使傳導神經沖動的乙酰膽堿無法水解, 在突觸處大量積累,干擾神經沖動的正常傳導, 使蟲體因過度興奮死亡[22]。常用的有敵百蟲(Trichlorfon)、甲基吡啶磷(Azamethiphos)、辛硫磷(Phoxim)和馬拉硫磷(Malathion)[23]等, 可用于寄生蠕蟲(吸蟲、絳蟲、線蟲、棘頭蟲和魚蛭)和甲殼類寄生蟲病的防治。
1.2 擬除蟲菊酯類
該類殺蟲劑包括從除蟲菊屬植物中提取的天然除蟲菊酯(Pyrethrin)及經合成衍生而成, 對昆蟲有強烈的觸殺、胃毒和熏蒸作用, 而對哺乳動物和禽類毒性低, 主要作用于軸突膜上的電壓門控鈉離子通道, 引起去極化過程, 擾亂蟲體神經的正常生理活動[24]。常用的有溴氰菊酯(Deltamethrin)、氰戊菊酯(Fenvalerate)、氯氰菊酯(Cypermethrin)、高效氯氰菊酯(Beta-cypermethrin)和高效氟氯氰菊酯(Lambada-cyhalothrin)[25]等, 主要用于甲殼類寄生蟲病的防治。
1.3 大環內酯類
大環內酯類抗寄生蟲藥是一類天然的或是半合成的具有十六元大環內酯環化合物, 該類化合物由兩大類組成, 即阿維菌素類(Avermectins)和美貝霉素類(Milbemycins)。阿維菌素類有阿維菌素(Abamectin)、伊維菌素(Ivermectin)、埃瑪菌素(Emamectin benzoate, 也叫甲維鹽)[26]等。該類藥物是一種神經毒劑, 主要激活存在于無脊椎動物中的谷氨酸門控氯離子通道(GluCls), 同時可以激活其他具有半胱氨酸環的離子通道, 如γ-氨基丁酸(GABA)受體、甘氨酸受體和煙堿型乙酰膽堿受體等, 促進相應神經遞質與次級神經元細胞膜或效應器細胞膜的結合, 產生抑制效應, 使蟲體麻痹死亡, 具有觸殺和胃毒作用[27,28], 常用于甲殼類寄生蟲病和線蟲病的防治。
1.4 苯甲酰脲類
苯甲酰脲類化合物是一種昆蟲幾丁質抑制劑,抑制幾丁質合成酶的活性, 進而抑制幾丁質合成和沉積, 從而阻止脫皮過程, 使昆蟲變態受阻, 導致不育及直接殺卵等而殺死害蟲[29,30]。常用的有二氟脲(Diflubenzuron, 除蟲脲)、氟苯脲(Teflubenzuron)和八氟脲(Lufenuron, 虱螨脲)[18]等, 可用于甲殼類寄生蟲病的防治。
1.5 新煙堿類
該類殺蟲劑來源于煙葉中提取的活性成分煙堿經修飾而成的化合物, 主要作用于煙堿型乙酰膽堿受體, 引起過度興奮而導致麻痹和死亡, 具有觸殺和胃毒作用, 高效低毒[31]。主要包括吡蟲啉(Imidacloprid)、啶蟲脒(Acetaniprid)、噻蟲啉(Thiacloprid)、噻蟲胺(Clothianidin)和呋蟲胺(Dinotefuran)等, 常用于甲殼類寄生蟲病的防治。
1.6 咪唑并噻唑類
該藥物是一類化學合成的廣譜驅線蟲藥, 能選擇性地打開線蟲神經和肌肉中乙酰膽堿受體(AChR)離子通道產生去極化, 鈣大量進入后導致肌漿鈣增加, 產生痙攣性肌肉收縮, 然后蟲體被機體清除[32]。主要有四咪唑(Tetramisole, 噻咪唑)和左旋咪唑(Levamisole)等, 常用于線蟲病的防治。
1.7 苯并咪唑類
目前認為此類藥物的作用機理是抑制蠕蟲的延胡索酸還原酶復合體, 影響其糖酵解途徑使得蟲體代謝障礙; 也有觀點認為該類藥物是細胞微管蛋白抑制劑以及能量代謝抑制劑, 阻礙蟲體在微管裝配過程中微管蛋白的聚合[33,34]。常用的有甲苯咪唑(Mebendazole)、芬苯達唑(Fenbendazole)、三氯苯達唑(Triclabendazole)和阿苯達唑(Albendazole)[35]等, 可用于扁形動物寄生蟲、線蟲和棘頭蟲引起的寄生蟲病。
1.8 異喹啉類
該類藥物中常用于驅殺寄生蟲的是吡喹酮(Praziquantel), 是一種含有吡嗪(Pyrazino-)和異喹啉(Isoquinolin-)雜環的衍生物[36], 通過改變電壓門控的鈣離子通道活性, 提高蟲體膜通透性, 導致鈣離子流入, 引起肌肉收縮和外皮出現空泡[37], 但可能有多種作用方式[38,39], 是一種高效的驅殺吸蟲和絳蟲的藥物[40,41]。
2 寄生蟲的抗藥性機制
藥代動力學描述的是藥物進入蟲體后的過程,包括吸收、分布、代謝和排出。藥物發生作用就是藥物分子與分子靶點(作用位點)發生相互作用的過程, 一方面是藥物分子對機體細胞產生毒性, 引起蟲體死亡; 另一方面蟲體會發生相應的生理變化和遺傳變異, 改變藥物吸收、分布、代謝和排出過程, 降低蟲體內的藥物濃度, 減少藥物的有害影響,從而保證機體存活[12]。使這個藥物動力學發生改變的生化和生理過程就是抗藥性機制(Mechanisms of resistance)[8]。
2.1 抗藥性機制類型
抗藥性機制主要包括3類(圖1): 通過改變穿透性(Penetration)或藥物吸收系統減少藥物進入和增加藥物外排(Efflux); 通過改變作用位點(Modified site of action)或靶標改變(Target modification)減少與殺蟲藥物結合; 通過增加新陳代謝(Enhanced metabolism)提高蟲體對殺蟲藥物的降解能力, 使藥物失活[8,42]。
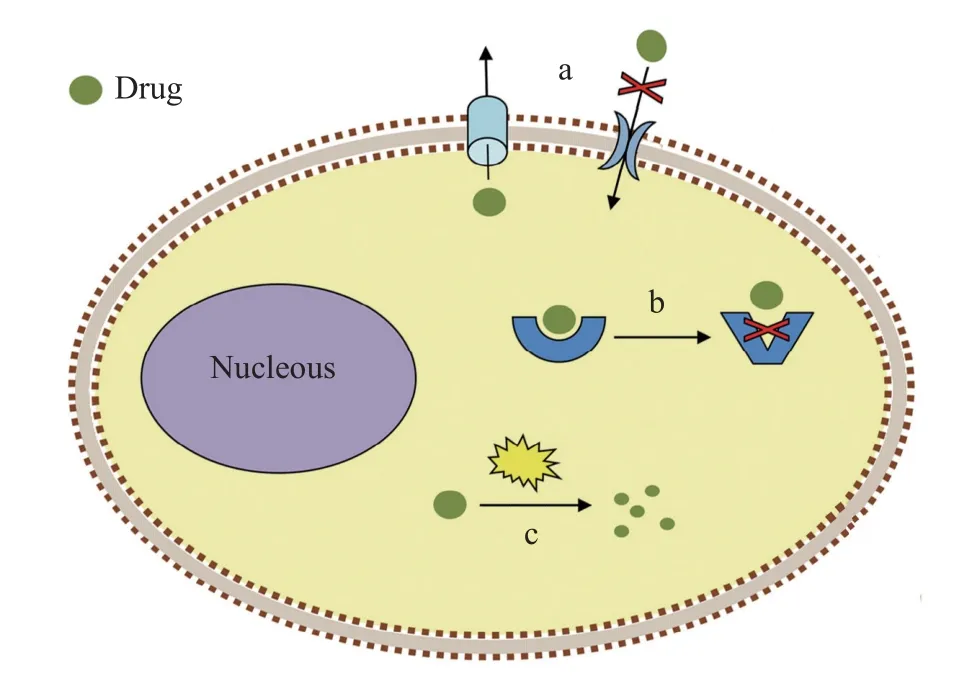
圖1 寄生蟲常見的抗藥性機制(修改自Garcia-Salcedo等[44])Fig.1 Common mechanisms for drug resistance of parasites(Modified from Garcia-Salcedo, et al.[44])
研究寄生蟲抗藥性產生機制不僅可開發檢測抗藥性的分子標記, 進行早期識別, 避免一些無效、有害的藥物治療, 而且有助于制定更合理的聯合用藥方案, 減少抗藥性產生, 延長藥物的有效性[12];另外, 還有助于準確錨定藥物靶標, 開發藥物類似物(Drug analogue)[43]。
穿透抗藥性機制通過蟲體的滲透屏障(表皮、消化道外膜和血腦屏障)結構和滲透性的改變,減少殺蟲藥物在體內的滲透量和殘留量, 主要包括減少表皮的藥物滲透 , 以及增加藥物的運輸和排出[42],如通過上調表皮沉積蛋白基因和幾丁質合成酶基因來增厚表皮[45,46], 以及通過P-糖蛋白(ABC轉運蛋白中的一種)增加藥物外排[47]。
靶標改變抗藥性機制指與殺蟲劑作用的靶標分子發生突變或者表達水平的改變, 減少了與殺蟲藥物結合, 從而降低蟲體對藥物的敏感性[43]。離子通道由一類膜蛋白組成, 起神經系統傳導信號和胞內信息交流作用, 是殺蟲藥物的主要靶標, 包括電壓門控的(Voltage-gated)離子通道, 如電壓門控鈉離子通道、鈣離子通道和鉀離子通道等, 以及配體門控的(Ligand-gated)離子通道, 也稱離子型受體; 配體結合導致通道蛋白結構的構象變化而打開通道, 離子流通過質膜, 如煙堿型乙酰膽堿受體(nAChRs) 、乙酰膽堿門控的氯離子通道(ACCs)或乙酰膽堿受體(AChRs)、谷氨酸鹽門控的氯離子通道(GluCls)、γ-氨基丁酸(GABA)門控的氯離子通道等[48]; 另外, 可水解神經遞質乙酰膽堿的乙酰膽堿酯酶(AChE)[49]和組成細胞骨架的β-微管蛋白也是重要的藥物靶標[50], 它們的基因突變會增加蟲體對藥物的耐受性。
增加新陳代謝抗藥機制通過體內解毒酶系表達量或活性的增加來提高蟲體對殺蟲劑的降解能力, 使藥物失活(Inactivation), 如將藥物由疏水型轉化為更易排出的親水型的細胞色素P450酶系(CYP450), 對藥物具有解毒作用的谷胱甘肽轉移酶(GST) 和羧酸酯酶(CarE)[11,51]。
2.2 主要寄生蟲類群的抗藥性機制
在通常情況下, 同種寄生蟲對同一類藥物的抗藥性機制是相同的。當寄生蟲對某種藥物產生抗藥性后, 對于結構近似或作用方式相同的藥物也會產生抗藥性, 這種現象被稱為交叉抗藥性(Cross-resistance)[10]。而多藥抗性(Multidrug resistance)則是寄生蟲對某種藥物產生抗藥性后, 對具有不同化學結構或不同作用方式的多種藥物產生抗藥性, 如ABC(ATP binding cassette)轉運蛋白中的P-糖蛋白過量表達就是較常見的多藥抗性機制[47,52]。
不同類別的寄生蟲具有不同的身體結構和生理機能, 對不同的殺蟲藥物產生的抗藥機制不盡相同, 以下是幾種主要寄生蟲類群的抗藥性機制。
寄生原蟲的抗藥性機制寄生原蟲的抗藥性機制研究主要集中在人類的瘧原蟲、非洲錐體蟲和利什曼原蟲[43,53]。氯喹(Chloroquine)曾是治療瘧疾的特效藥, 但現在世界各地的瘧原蟲對氯喹產生了普遍的抗藥性。氯喹的主要作用是抑制蟲體食物泡內血紅蛋白酶活性和降低pH, 從而使瘧原蟲不能消化所攝取的血紅蛋白而致使核破裂。因此,瘧原蟲對氯喹的抗藥機制是位于食物泡膜的一種ATP結合盒轉運蛋白(ATP-Binding-Cassettes, ABC)Pgh1基因發生點突變, 表達上調, 增加了對氯喹的泵出[54]。另外, 編碼食物泡另一種膜蛋白PfCRT基因的突變, 通過調節食物泡內的pH增加對氯喹的抗性[55]。后來隨著葉酸拮抗劑(Antifolates)大量使用, 瘧原蟲很快也產生了抗藥性, 主要通過二氫葉酸還原酶(DHFR)和二氫蝶酸合成酶(DHPS)的點突變, 改變葉酸拮抗劑藥物與作用靶點葉酸的結合[56]。現在盡管青蒿素(Artemisinin)及其衍生物是治療瘧疾的常用藥物, 但是瘧原蟲也產生了抗藥性, 由于蟲體食物泡附近編碼Kelch13蛋白的基因發生點突變, 失活的蛋白減少了血紅蛋白的內吞作用, 進而減少了對青蒿素及其衍生物的激活, 導致瘧原蟲對青蒿素及其衍生物產生抗藥性[57]。
雞的球蟲病非常普遍和嚴重, 莫能菌素(Monensin)等聚醚類離子載體(Polyether ionophores)是常用藥物, 主要作用于孢子體(Sporozoites)和裂殖體(Merozoites)的細胞膜, 大量的陽離子進入細胞內使細胞內外形成滲透壓差, 大量水分進入蟲體, 最終導致蟲體膨脹破裂[15], 艾美球蟲(Eimeria tenella)對這類藥物產生了抗藥性, 但其復雜的抗藥性機制仍不清楚[58]。
寄生蠕蟲的抗藥性機制寄生線蟲的抗藥性機制是研究最多的。線蟲常用的殺蟲藥物有苯并咪唑類、大環內酯類、左旋咪唑和噻嘧啶[13]。線蟲對苯并咪唑類藥物的抗藥性機制是蟲體β-微管蛋白基因點突變, 引起微管蛋白與藥物結合減少而導致抗藥性產生[59—61]。而線蟲對大環內酯類藥物的抗藥性則是由于Pgp基因點突變, 導致P糖蛋白上調, 而引起藥物外排增加[62], 以及多種離子通道或受體改變, 如煙堿型乙酰膽堿受體、乙酰膽堿門控的氯離子通道、谷氨酸鹽門控的氯離子通道和γ-氨基丁酸門控的氯離子通道等, 從而減少藥物與作用靶點的結合[48]。
其次, 抗藥性研究較多的是人類的肝吸蟲(包括片吸蟲、華支睪吸蟲和后睪吸蟲等)和血吸蟲,吡喹酮和苯并咪唑類是扁形動物寄生蟲的常用殺蟲藥物[63]。多種片吸蟲對三氯苯咪唑(Triclabendazole)都產生了抗藥性, 抗藥性機制包括P-糖蛋白增加藥物外排和谷胱甘肽轉移酶增加藥物代謝作用[11,64];在一些地區, 血吸蟲對吡喹酮也產生了抗藥性, 主要通過點突變減少藥物與電壓門控的鈣離子通道β亞基(一種胞漿蛋白)的結合[65]。
絳蟲的抗藥性機制也有少量報道, 電壓門控的鈣離子通道同樣也是吡喹酮在豬肉絳蟲(Taenia solium)上的關鍵作用靶點[66], 犬復孔絳蟲(Dipylidium caninum)在對吡喹酮出現明顯抗藥性后檢測到β亞基類似物的[67]。鈣離子通道β亞基的突變, 降低了吡喹酮與蟲體鈣離子通道的結合, 不會引起肌肉收縮和外皮出現空泡[38,39]。
甲殼類寄生蟲的抗藥性機制甲殼類和昆蟲都屬于節肢動物, 昆蟲的殺蟲藥物也常用于甲殼類寄生蟲, 因此, 甲殼類寄生蟲的抗藥性機制與昆蟲的類似[18]。Heumann等[68]在鮭瘡痂魚虱克隆到一種新的P-糖蛋白基因SL-PGY1, 后被證實鮭瘡痂魚虱對甲維鹽的抗藥性是通過增加P糖蛋白表達量從而增加了藥物的外排[69]; 另外, 還在抗甲維鹽的魚虱檢測到乙酰膽堿受體和GABA-gated的氯離子通道的mRNA 轉錄水平顯著降低, 推測是因為點突變引起這兩類離子型受體表達量降低, 從而減少作用靶點與藥物結合[70]; 鮭瘡痂魚虱對甲基吡啶磷的抗藥性機制是乙酰膽堿酯酶基因(ace1a)上一個等位基因(362Tyr)突變而引起[71]; 它對擬除蟲菊酯類藥物的抗藥性機制則是單氧化酶介導的解毒作用增強[72]。
不管是陸生動物的寄生蟲, 還是水生動物的寄生蟲, 相同類群的寄生蟲產生的抗藥性機制應該是相同或類似的。因此, 上述陸生動物和人類的寄生蟲抗藥性研究可為水產養殖中原生動物寄生蟲、寄生蠕蟲和甲殼類寄生蟲的抗藥性機制研究提供參考, 以及為新型殺蟲藥物開發提供靶標。
3 抗藥性控制策略
抗藥性產生的遺傳機制是在藥物選擇壓力下蟲體發生基因突變, 因此, 若要減少抗藥性的產生,就必須從治療策略(Treatment strategies)和保護敏感蟲體種群兩方面來采取措施。合理的治療策略可以阻止和減少抗藥性產生, 保護敏感蟲體種群則可促進抗藥性逆轉。
3.1 治療策略
在陸生系統中, 主要通過在時間和空間上改變化學藥物使用模式, 通過增加治療的異質性來減少抗藥性產生, 這是因為多種藥物聯用或交替使用可延緩某一種藥物帶來強大選擇壓力, 通常治療的異質性越高, 抗藥性進化越慢[10]。這些用藥策略主要包括以下四種: 響應換藥(Responsive alternation),當蟲體對一種藥物產生抗藥性后, 換用另一種藥物,這種策略一般在不易產生抗藥性的寄生蟲中使用;定期使用或輪換用藥(Periodic application, sequential use, rotation), 即兩種或多種藥物定期輪換使用,換藥時間可長于蟲體世代, 也可短于世代; 錯區用藥(Mosaic), 兩種或多種藥物同時在不同區域使用,即一個區域分別用一種不同的藥物; 聯用或混用(Combination, mixture), 兩種或多種藥物同時在一次治療中使用, 聯用藥物可以是全劑量, 也可是半劑量[10, 16]。
盡管聯合用藥比其他幾種用藥策略更優, 但在實際應用中到底使用哪一種策略, 主要取決于治療對象, 在人類的瘧疾和農作物害蟲防治中, 常采用聯合用藥策略, 但在細菌病的抗生素治療中, 特別是群體中, 聯用策略并不合適[10]。基于海虱病原模型, 模擬了7種用藥場景并同時檢測海虱種群產生抗藥性水平, 結果發現聯合用藥是最有效的策略[16]。雞球蟲病的用藥策略是比較成熟的, 在某一生長階段采用聯合用藥策略, 而在整個養殖過程中采用定期輪換用藥計劃[15]。
3.2 敏感蟲體種群保護
理論上講, 建立未用藥保護區或殘遺種保護區(Refugia), 保護部分敏感蟲體種群可增加敏感等位基因頻率, 延緩或逆轉抗藥性[73]。在家畜寄生線蟲的抗藥性管理中, 基于殘遺種保護區的控制策略就是進行選擇性治療, 即只有一部分家畜種群用藥。但是很多因素會影響該策略的使用效果, 如保護區寄生蟲種群的抗藥性水平和遺傳多樣性、治療寄生蟲種群的比例和治療效果以及寄生蟲生長、繁殖的生活史特征等[73]。
已經產生的抗藥性一般難以逆轉。吸蟲和線蟲對苯并咪唑類藥物產生了抗藥性, 停止用藥后表型抗性仍能保持多年[74, 75]; 將具有甲維鹽抗性的海虱(Caligus rogercresseyi)在實驗室連續養殖7代后,對甲維鹽的敏感性仍然沒有恢復[76]; 對有機磷和擬除蟲菊酯具有抗藥性的鮭瘡痂魚虱在第十代子代中也沒有發現抗藥性降低[18]。但是通過從保護區內引入敏感種群來增加敏感等位基因頻率, 可恢復抗藥蟲體的敏感性或逆轉抗藥性[77,78]。
3.3 對水產動物寄生蟲抗藥性控制的啟示
從降低寄生蟲抗藥性產生的策略來分析, 抗藥性產生速度與養殖方式和寄生蟲種類密切相關。我國大部分是室外的池塘養殖, 而這種養殖方式通常只在生長季節用藥, 藥物壓力是不持續的; 而且池塘一般呈現斑塊分布, 具有空間異質性, 不同養殖區域的養殖品種不同, 用藥種類和時間也不同,可看作錯區用藥策略; 另外, 大多數寄生蟲的宿主特異性不強, 如原生動物寄生蟲、甲殼類寄生蟲、復殖吸蟲和線蟲, 那么鄰近區域其他魚類上的相同寄生蟲可起到殘遺種保護區作用。因此, 我國池塘養殖模式中大多數魚類寄生蟲產生的抗藥性相對較慢。
從抗藥性產生的遺傳機制來看, 生活世代短、具有有性繁殖的寄生蟲更容易產生抗藥性[14]。單殖吸蟲不僅具有直接的生活史, 世代短, 有性生殖,且具有較強的宿主特異性[79], 因此, 單殖吸蟲可能是最容易產生抗藥性的寄生蟲類群之一。寄生于鰻鱺的擬指環蟲在低劑量甲苯咪唑的作用下, 經過7個世代后開始出現明顯的抗藥性[80], 在短短的十年內, 甲苯咪唑對歐洲鰻鱺的指環蟲病幾乎沒有治療效果[17]; 在我國的鰻鱺養殖中, 也發現了擬指環蟲的抗藥性問題[81]。
不管是室外的池塘養殖和設施養殖, 還是室內的循環水養殖, 高密度的集約化養殖是我國水產養殖的一種趨勢, 針對頻頻發生的寄生蟲病使用藥物防治不可避免。根據陸生動物寄生蟲抗藥性的控制策略, 首先要布局好養殖品種和養殖方式, 如多品種混養, 不同品種的錯區養殖; 其次是在用藥方式上, 不能長期只用一種藥物, 要多種藥物聯用。
4 抗藥性的主要檢測方法
抗藥性蟲體表型特征變化是最直接的抗藥性識別方法。生物體在產生抗藥性過程中, 會消耗較高能量代價, 或者產生一些不利因素來降低適合度,稱為適合度代價(Fitness costs), 如繁殖力降低、擴散力削弱等種群生物學性狀的表型變化[82]。鑒定適合度代價在識別和限制抗藥蟲體種群的綜合管理中具有明顯優勢, 種群相對適合度的表征有兩種,即種群抗藥性基因突變頻率的變化, 以及抗藥性種群生物學性狀的變化[83]。這種抗藥性引起的適合度代價在昆蟲中比較常見[82], 在抗藥瘧原蟲中也存在[84,85], 但在抗甲維鹽魚虱的第四代子代中沒有發現適合度代價[86]。
4.1 生物測定法(Bioassays)
利用生物體對殺蟲藥物的反應來定性或定量測定抗藥性強弱, 這些反應包括死亡或停止運動等,衡量參數常用半致死濃度或90%致死濃度等, 這是傳統的抗藥性檢測方法。該方法的優點是在抗藥性機制未知情況下仍然可以檢測, 缺點是測試敏感性低, 在比較不同地區生物體的抗藥性水平時受到限制[18]。在鮭瘡痂魚虱的抗藥性測定中, 50%的有效濃度(EC50)的估計需要多個藥物濃度[87]; 因此, 后來又有人建立了一種基于劑量-殺蟲效果模型的單劑量快速測定方法[88]。
4.2 分子檢測法(Molecular assays)
在抗藥性機制已知情況下, 利用分子生物學技術檢測殺蟲藥物作用靶標的抗性點突變或解毒代謝酶基因的增強表達, 其中基因突變檢測技術可應用于檢測殺蟲藥物作用靶標的基因突變, 而基因差異表達技術則應用于檢測解毒代謝酶基因的增強表達[89]。有時候具體的抗藥性機制可通過抗性個體展示的表型特征進行鑒別, 但通常情況下需要高通量測序, 并對大量的候選基因進行功能鑒定[18]。
5 水產動物寄生蟲抗藥性研究現狀
寄生原蟲、扁形動物寄生蟲和甲殼類寄生蟲是水產養殖動物重要的寄生蟲類群。在寄生原蟲引起的寄生蟲病中, 小瓜蟲病危害最嚴重, 但由于缺少真正的有效藥物, 沒有一種藥物是長期使用的,因此還沒有抗藥性的研究報道[90]。
在扁形動物寄生蟲病中, 吡喹酮是常用藥物,盡管人類的血吸蟲、肝吸蟲和絳蟲都對該藥物產生了抗藥性, 但魚類寄生的絳蟲和復殖吸蟲仍然保持較高的敏感性, 可能是吡喹酮在水產中的使用時間較短, 而且也沒有反復使用[40]。在挪威的大西洋鮭養殖中, 吡喹酮常用于真溝絳蟲(Eubothriumspp.)的治療, 該絳蟲對吡喹酮已經產生了抗藥性[91,92]。擬指環蟲是歐洲鰻鱺養殖中危害最嚴重的單殖吸蟲, 甲苯咪唑是最有效的藥物[93], 最開始1 mg/L的甲苯咪唑在24h內可100%殺滅擬指環蟲[94,95], 由于循環水養殖模式和長期只使用這一種殺蟲藥物, 甲苯咪唑的治療效果越來越差, 使用濃度每年要提高6—12倍, 而且已經很難有效控制指環蟲病的發生[17]。
在甲殼類寄生蟲中, 海虱(L.salmonis,Caligusspp.)是影響大西洋鮭養殖最重要的寄生蟲, 盡管每個國家采取的防治方案不同, 但由于殺蟲藥物的長期使用, 對海虱的治效果已明顯下降[18]。挪威從2014年開始對鮭瘡痂魚虱的抗藥性進行監測, 發現對溴氰菊酯、甲基吡啶磷和甲維鹽一直保持較高的抗藥性, 對過氧化氫和淡水的耐受性也在增加[96]。2008年蘇格蘭報道鮭瘡痂魚虱對甲維鹽的敏感性降低了7倍[68]。加拿大在2010—2012年對22個大西洋鮭養殖場的調查中, 發現魚虱對甲維鹽敏感性降低[97]。在智利, 甲維鹽和溴氰菊酯對海虱(C.rogercresseyi)的治療效果也明顯降低[76,98]。
綜上所述, 單殖吸蟲和甲殼類寄生蟲的抗藥性都是在歐洲鰻鱺和大西洋鮭的集約化養殖中, 長期反復使用殺蟲藥物而產生的。在我國, 水產養殖歷史更悠久, 在一些常見、頻發的寄生蟲病防治中,藥物使用年限更長, 不規范用藥也更嚴重, 盡管抗藥性的研究報道較少, 但在一些水產養殖區域, 寄生蟲病的防治效果明顯變差, 殺蟲藥物的使用劑量越來越高, 這已經成為擺在水產養殖生產面前的重要問題。而且隨著水產養殖規模和集約化程度逐漸提高, 寄生蟲抗藥性問題將會更加嚴重和突出。
6 水產動物寄生蟲抗藥性研究展望
陸生動物寄生蟲和鮭鱒魚類寄生海虱的抗藥性研究提供了成功案例和經驗, 但我國水產養殖動物寄生蟲的抗藥性研究幾乎還是空白。因此, 選擇合適的寄生蟲類群, 開展寄生蟲抗藥性研究, 是亟待開展的基礎研究工作。
我國水產養殖動物寄生蟲抗藥性可從以下幾個方面開展研究: (1)建立抗藥性生物檢測方法, 在重點養殖區域調查寄生蟲對藥物的敏感性, 看哪些寄生蟲類群產生了抗藥性。(2)針對抗藥寄生蟲, 通過對敏感和抗性蟲株進行轉錄組測序, 找出差異表達的候選基因, 再利用生物信息學分析、RNAi 技術或CRISPR-Cas9基因編輯技術鑒定出抗藥性基因, 闡明抗藥性分子機制[18]。(3)利用抗藥性基因,建立抗藥性的分子檢測方法, 對全國范圍內主要水產養殖區進行寄生蟲抗藥性普查。(4)研究寄生蟲抗藥性產生規律, 建立合理的藥物輪用和組合策略,以及殘遺種保護區等措施, 避免或減緩抗藥性產生。(5)將抗藥性基因作為抗藥性治療靶點, 利用RNAi 技術抑制抗藥性基因的過量表達[48,99], 研發P-gp轉運蛋白和解毒酶系的抑制劑[100]等, 以減少寄生蟲的抗藥性產生。
通過上述研究, 最終形成一個完整的寄生蟲抗藥性監測和防治體系, 保證現有殺蟲藥物的有效性,保障水產養殖的綠色健康和可持續發展。
猜你喜歡
四川勞動保障(2021年9期)2022-01-18 05:11:08
文苑(2018年21期)2018-11-09 01:23:06
當代陜西(2018年9期)2018-08-29 01:21:00
當代陜西(2017年12期)2018-01-19 01:42:33
暨南學報(哲學社會科學版)(2016年9期)2017-01-15 13:52:00
中國衛生(2016年9期)2016-11-12 13:28:08
中國衛生(2015年9期)2015-11-10 03:11:12
醫學研究雜志(2015年12期)2015-06-10 06:57:46
中國衛生(2014年3期)2014-11-12 13:18:12
中國火炬(2014年4期)2014-07-24 14:22:19
