肝腸胞蟲對凡納濱對蝦腸道和肝胰腺菌群的影響
2024-02-07 07:29:56李舢舢趙晨曦鄭俠飛戴文芳林志華柳敏海
水生生物學報 2024年2期
李舢舢 趙晨曦 吳 波 鄭俠飛 戴文芳 林志華 柳敏海
(浙江萬里學院寧海海洋生物種業研究院, 浙江省水產種質資源高效利用技術研究重點實驗室, 寧波 315000)
水產養殖為不斷增長的全球人口提供食物來源方面發揮著關鍵作用。據聯合國糧食及農業組織(Food and Agriculture Organization of the United Nations, FAO)發布的《2022年世界漁業和水產養殖狀況》報告[1], 水產品對糧食安全和營養貢獻之大前所未有。全國水產品總產量6690.29萬噸, 養殖產量5394.41萬噸, 甲殼類動物位居養殖產量第二,其中約70%的甲殼類動物來自蝦和蟹[2]。凡納濱對蝦(Litopenaeus vannamei)是南美洲太平洋沿岸、秘魯北部及亞洲等地區主要養殖物種之一, 其養殖過程中也面臨一些挑戰[3], 如頻繁暴發的疾病: 白斑綜合征(White Spot disease, WSD)[4]、肝胰腺微孢子蟲病(Hepatopancreatic microsporidiosis,HPM)[5]、急性肝胰腺壞死病(Acute hepatopancreatic necrosis disease, AHPND)[6]、Taura綜合征(Taura syndrome,TS)[7]和黃頭病(Yellow head disease, YHD)[8], 其中HPM已給凡納濱對蝦養殖造成了巨大的損失[9]。
蝦肝腸胞蟲(Enterocytozoon hepatopenaei,EHP)是引起對蝦HPM的主要病原[10], 已成為對蝦養殖業最致命的威脅之一, 肝胰腺是易受EHP感染的組織,在腸道和胃中也檢測到EHP感染[11,12]。在EHP感染早期, 對蝦通常不會死亡, 攝食正常且無明顯疾病癥狀; 感染后期, 對蝦生長緩慢或停滯生長, 產量嚴重下降、經濟損失嚴重。最近的研究表明由EHP感染引起的肝胰腺微孢子蟲病已在不同地區報告, 包括越南、印度、文萊、中國、印度尼西亞、馬來西亞、委內瑞拉和澳大利亞等國家[13,14—18]。自2013年以來, 我國各地養殖的凡納濱對蝦先后出現EHP感染, 且感染率居高不下[19]。肝胰腺作為蝦的主要消化腺, 分泌消化酶, EHP感染肝胰腺導致蝦生長不良和免疫力受損。Ha等[20]報道越南養殖的斑節對蝦感染EHP后生長緩慢, 且伴有白便癥狀;Flegel等[21]發現了與 Ha 等[20]報道相同現象, 認為泰國和越南養殖的斑節對蝦及凡納濱對蝦的白便癥狀與EHP有一定的關系, EHP已成為制約凡納濱對蝦養殖業健康發展的最主要病害之一[22]。
目前, 有關EHP感染是否影響凡納濱對蝦腸道及肝胰腺中細菌群落的研究鮮有報道。因此, 本研究比較了凡納濱對蝦健康樣本和感染EHP的發病樣本中其腸道和肝胰腺微生物的結構、多樣性和組成差異, 進一步探究了其腸道及肝胰腺菌群介導的功能。本研究旨在確定健康對蝦、發病對蝦之間的細菌組成差異, 篩選腸道和肝胰腺中的優勢菌屬用于區分健康對蝦和發病對蝦。
1 材料與方法
1.1 實驗材料
養殖池塘投放同一批次凡納濱對蝦苗種, 養殖48d后, 表現出明顯的體長和體重差異, 2022年5月31日從養殖池中隨機采集生長差異的對蝦, 大蝦(D)平均體重(8.11±0.36) g, 平均體長(86.42±1.18) mm;小蝦(X)平均體重(1.73±0.12) g, 平均體長(50.04±1.21) mm; D和X的平均體重和體長均存在極顯著差異(P<0.01)。在無菌操作下采集對蝦腸道和肝胰腺, 分別設3組平行且保存無菌凍存管中, 并置于-80℃冰箱備用。D和X進行PCR檢測驗證(圖1),X與肝腸胞蟲病蝦序列的相似度高達99.51%, 因此標記D為健康對蝦, X為發病對蝦。
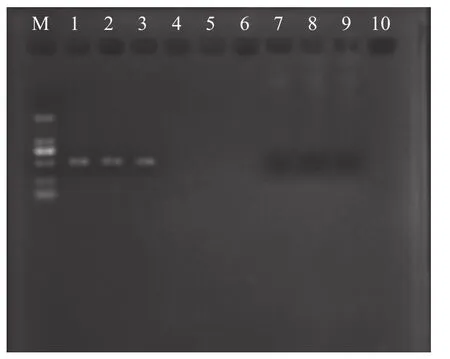
圖1 PCR擴增結果圖Fig.1 PCR amplification results graph
1.2 細菌DNA提取及高通量測序
從-80℃冰箱中取出樣品, 冰上融化后, 采用SDS法提取樣本總基因組DNA。在1%的瓊脂糖凝膠上檢測DNA的濃度和純度。根據濃度, 用無菌水稀釋樣品DNA至l μg/μL。利用正向引物(5′-GTGCC AGCMGCCGCGGTAA-3′)和反向引物(5′-CGCC AG CGCGGGTAA-3′)對16S rRNA基因V3—V4可變區進行擴增。以10 ng DNA樣本為模板進行PCR擴增, 反應系統: 98℃預變性1min, 98℃變性10s, 50℃退火30s, 72℃延長30s, 進行30次循環, 最后72℃延長5min, 并在4℃保存。使用QIAquick PCR純化試劑盒(Qiagen, GmbH, Hilden, 德國), 對每個樣品的PCR產物進行合并及純化。將健康對蝦腸道(D.C)、健康對蝦肝胰腺(D.G)、發病對蝦腸道(X.C)、發病對蝦肝胰腺(X.G)及水體菌群(S)這5組樣品送測, 最后在Illumina NovaSeq平臺上進行測序。
1.3 高通量測序原始數據處理
將配對末端讀數與FLASH (V1.2.7, http://ccb.jhu.edu/software/FLASH/)連接, 然后使用Quantitative Insights Into Microbial Ecology (QIIME v1.9.0,http://qiime.org/)通道進行處理。通過 QIIME中的質量分數、序列長度、模糊序列及引物錯位閾值等信息來篩選出嵌合體, 再利用 UCHIME (V11, http://www.drive5.com/usearch/manual/uchime_algo.html)技術將嵌合體序列剔除, 從而得到有效的序列。采用USEARCH的UPARSE (V7.0.1001, http://www.drive5.com)算法進行樣品運算分類單元(OTU)聚類分析, 相似度大于97%的序列聚為一類, 篩選每個OTU的代表性序列。對于每個具有代表性的序列,使用基于Mothur算法的SILVA123 (http:// www.arbsilva.de/)數據庫對分類信息進行注釋。采用MUSCLE軟件(v5.1, https://www.drive5.com/muscle/)對多個基因序列進行了比對, 研究不同OTUs的系統發育關系, 并比較不同樣本(組)中優勢種間的差異。對α多樣性(Chao1指數、Ace指數、Simpson指數和Shannon指數)進行分析; 在生物學分類的門和屬水平上對樣本的微生物的群落組成及相對豐度進行統計; 利用Tax4Fun對SILVA數據庫中16S序列的注解進行 KEGG功能預測。
1.4 數據統計
利用SPSS 18.0開展單因素方差分析, 比較各組間水體、肝胰腺、腸道之間的微生物群落多樣性的顯著差異, 用基于binary_jaccard距離的主坐標分析PCoA和群落相似度分析ANOSIM比較各組間水體、肝胰腺、腸道之間的微生物群落組成結構的顯著差異。
2 結果
2.1 樣品測序數據質量評估及復雜度分析
D.C、D.G、X.C、X.G和S五組樣品得到的原始序列1214292條, 通過過濾和雙端拼接最終獲得了1014269個tags數, 其中有效數據占83.53%。在相似性超過97%的水平上共發現了6449個OTU數。
統計和比較所有樣品中的 OTU數, 繪制花瓣圖(圖2)。從圖2中可見, 5組樣品中共有的OTU數為80, 其中D中的特有OTU數目與S中特有的OTU數目較為接近, 且S中特有的OTU數目最多, 說明水體菌群的豐富度和多樣性最高、D菌群次之。
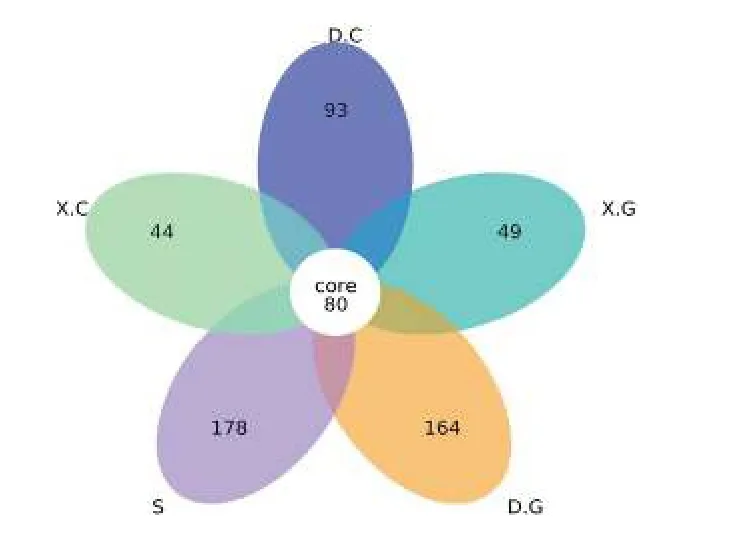
圖2 各樣本間OTU的差異性Fig.2 Variability of OTU among samples
稀釋性曲線間接地反映出樣品中的物種豐富度, 5個樣本的物種數據構建稀釋性曲線見圖3。隨著測序序列的增加, 5組樣品所能聚類的物種數量逐漸趨于飽和, 表明隨著測序量增加, 新的菌系已經不再顯著增加, 菌群分析結果具有較高的可靠性和充分性。
2.2 菌群多樣性分析
Alpha多樣性分析結果對各樣品在97%一致性閾值下的Alpha Diversity分析指數進行統計分析(表1)。D和X中腸道細菌和肝胰腺細菌的群落的豐富度及多樣性各不相同, S、D.C、X.C、D.G、X.G樣品中Shannon指數、Simpson指數、Chao 1指數、ACE指數依次減少。從Shannon 指數及Simpson指數來看, D.C與X.C、D.G與X.G及S與X.C Shannon 指數及Simpson指數之間無顯著差異(P>0.05),而D.C中細菌群落與X.G細菌菌落存在差異(P<0.05), S細菌群落與D.G中細菌菌群存在差異(P<0.05)。從Chao1和ACE可知, D.C與X.C、S與X.C及S與X.G細菌群落之間豐度差異顯著(P<0.05), 而D.G與X.G、D.G與X.C及S與D.C細菌群落之間豐度無明顯差異(P>0.05)。

表1 各樣品α-多樣性指數差異表Tab.1 Differences in alpha-diversity indices among samples (mean±SD, n=3)
Bate多樣性分析結果非定向β多樣性是對某些環境或實驗因素響應的群落結構變化的度量。使用OTU水平的細菌群落的PCoA來可視化D和X中腸道和肝胰腺細菌群落的差異(圖4),D.C和X.C微生物群落、D.G和X.G微生物群落的組成是均勻的且高度相似性, 而S的細菌群落較蝦腸道及蝦肝胰腺的細菌群落相似性較低。
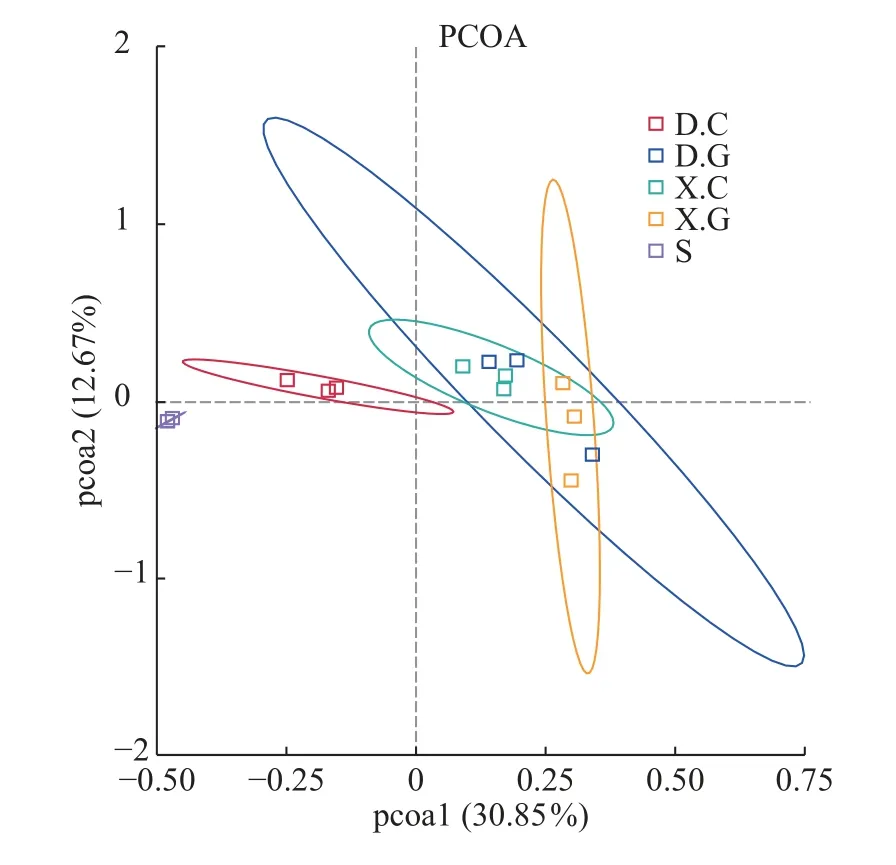
圖4 不同樣本之間細菌群落的β多樣性Fig.4 Beta diversity of bacterial communities among different samples
ANOSIM檢驗結果進一步證實了水體、肝胰腺和腸道之間的微生物群落差異。由表2可知,D.G中細菌群落與S中細菌群落存在極顯著差異(P<0.001), D.G中細菌群落與D.C細菌群落、X.C細菌群落與D.G細菌群落、X.G群落與S中的細菌群落均顯著差異(P<0.05)。
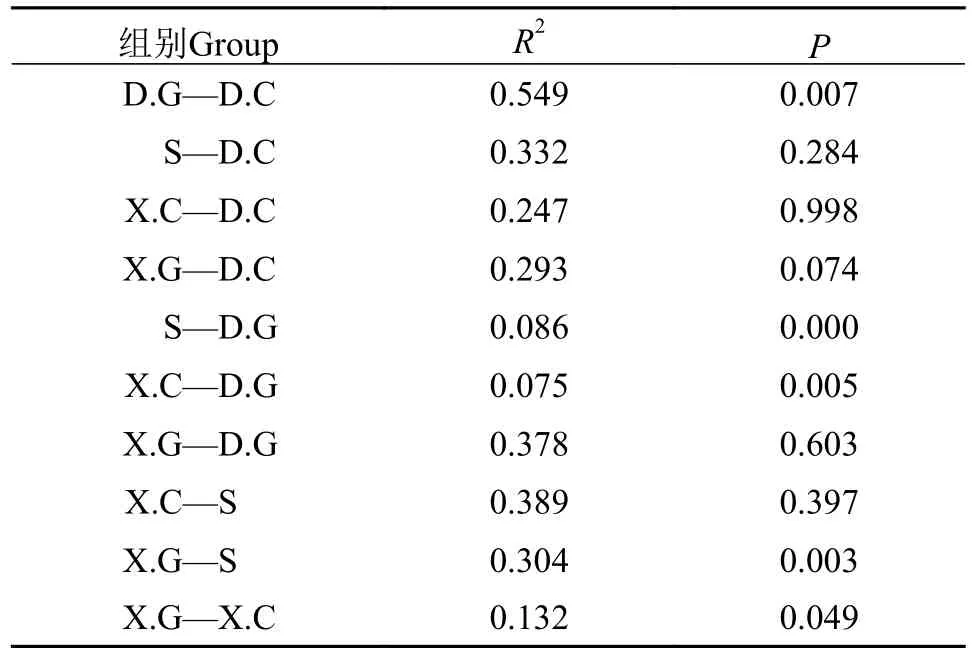
表2 不同樣本組之間方差的置換多變性分析Tab.2 Displacement multivariance analysis of variance between different sample groups
2.3 細菌群落的組成
基于門分類水平上的物種注釋及分析基于門(Phylum)分類水平構建相對豐度累加圖(圖5)。D.C主要是擬桿菌門(Bacteroidetes)、變形菌門(Pro-teobacteria)及厚壁菌門(Firmicutes), 以上三門微生物占菌群總量的94.42%; 變形菌門是D.G、X.C、X.G及S優勢菌群, 占比分別為86.19%、38.18%、87.74%、40.68%。在D.G及X.G中未檢測到其他共有的優勢菌門, 而在X.C及S中還檢測到擬桿菌門為共有優勢菌門。D.C與X.C共有擬桿菌門、變形菌門及厚壁菌門3個優勢菌門。D.C與S有擬桿菌門、變形菌門2個優勢菌門。五組樣品的細菌種群在門水平上的組成和分布說明在同一養殖池塘中D.C、X.C、S、D.G、X.G中細菌菌群組成有密切的關聯。
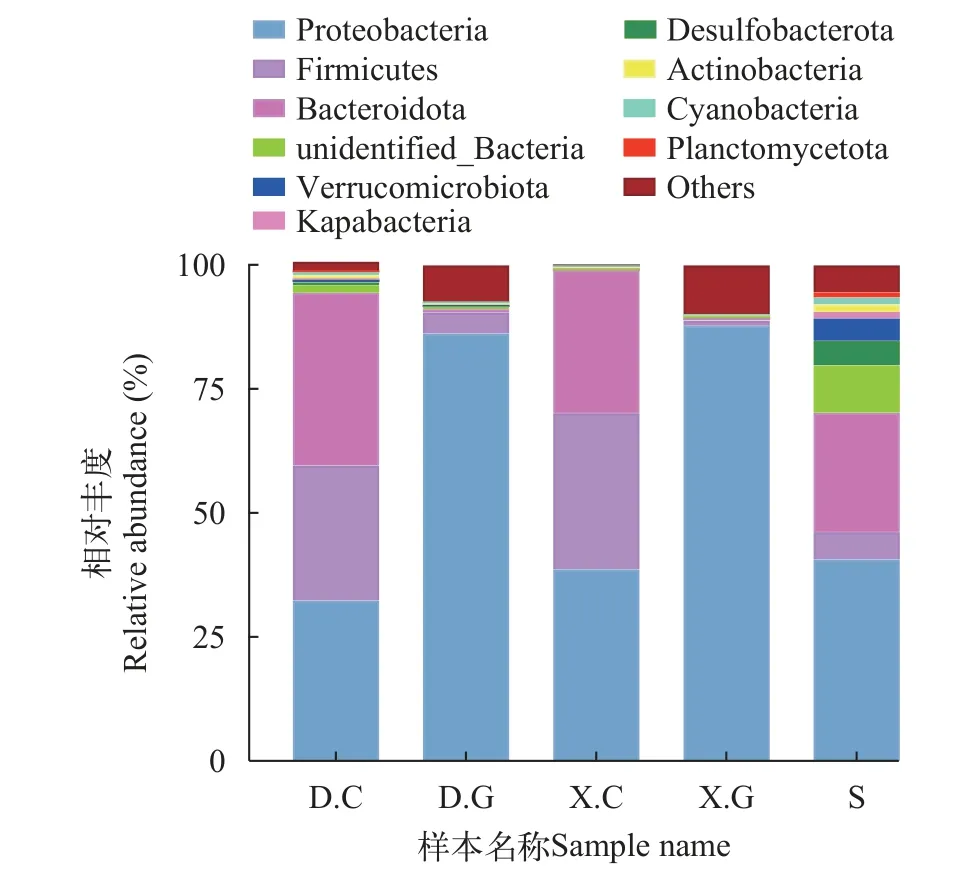
圖5 各組樣本中的優勢細菌門的平均相對豐度分布情況Fig.5 Mean relative abundance distribution of the dominant bacterial phylum in each group of samples
基于屬分類水平上樣本菌群組成分析在屬(Genus)水平上, D和X的腸道及肝胰腺內優勢細菌的組成及豐度存在明顯不同(圖6)。D.C優勢菌屬為埃希桿菌-志賀菌屬(Escherichia-Shigella)、海綿菌屬(Spongiimonas), 而X.C優勢菌屬為希瓦式菌屬(Shewanella)、巨球型菌屬(Megasphaera)、克雷伯氏菌屬(Klebsiella)。D.G優勢菌屬是假單胞菌屬(Pseudomonas)、漢氏鹽單胞菌屬(Halomonas), 而X.G優勢菌屬為貪銅菌屬(Cupriavidus)。由于S中的菌群的多樣性較高, 因此具有優勢豐度的菌群數量居多。

圖6 菌群在屬水平豐度聚類熱圖Fig.6 Abundance clustering heat map of the bacteriophage at the genus level
2.4 菌群與預測功能特征的相關性
利用Tax4Fun2預測菌群中的潛在功能并繪制功能豐度聚類圖(圖7)。結果顯示: D.C與X.C、D.G與X.G存在明顯差異, 其中D.C菌群的主要功能為氨基酸代謝、X.C菌群的主要功能為轉運RNA;D.G菌群的主要功能為二分組系統, 而X.G菌群的主要功能為肽酶。
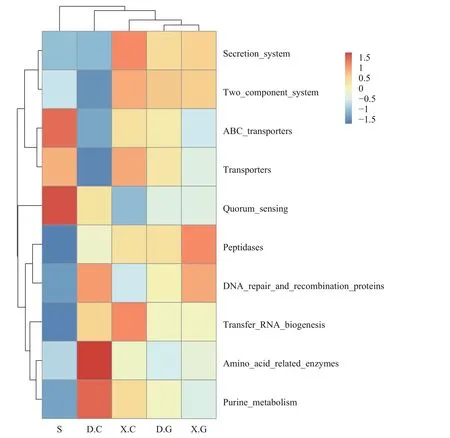
圖7 Tax4Fun2功能豐度聚類圖Fig.7 Tax4Fun2 functional abundance clustering map
3 討論
3.1 腸道及肝胰腺菌群生物多樣性分析
凡納濱對蝦作為產量最高的經濟蝦之一, 受到了養殖戶的廣泛關注和選擇[23]。與其他蝦相比, 微生物菌群與健康、疾病、生長速度有關[24], 與其他種類蝦相比, 凡納濱對蝦的生長速度和繁殖速度非常迅速, 其所需的高蛋白飼料可以實現大規模的養殖。腸道菌群為宿主提供有益的功能, 如營養素的吸收、消化酶的產生、通過激活對病原體的保護機制產生宿主代謝必需元素和免疫反應, 以及與致病菌產生競爭關系[25]。雖然已有研究揭示對蝦腸道內的細菌組成[26], 最近研究發現[27], 正常水生生物和患病水生生物具有不同功能的消化系統, 它們產生的微生物群落也具有多樣性。肝胰腺是甲殼類動物消化、吸收和儲存營養物質的重要器官, 在調節宿主先天免疫方面也發揮著關鍵作用。自2009年泰國研究者Peng和Pu[12]首次發現一種未知的微孢子蟲, 經過后續的研究命名為EHP。EHP主要在凡納濱對蝦的腸道表皮表層、肝臟、胰臟及附著在肝胰腺小管上皮的細胞上, 被感染的對蝦其病變位置出現腫大現象, 并且有大量的嗜堿性包涵體存在于肝胰臟小管上皮細胞的細胞質內。研究表明凡納濱對蝦生長緩慢跟EHP的感染有直接關系[28]。本研究表明: 健康蝦的腸道和肝胰腺菌群的豐度及多樣性比發病蝦的腸道和肝胰腺的豐度及多樣性更高, 表明受EHP侵染對凡納濱蝦腸道及肝胰腺菌群多樣性造成了一定的影響, 這一結果與吳金鳳等[29]、段健誠等[30]研究的結果相似。
3.2 腸道及肝胰腺菌群結構分析
腸道是水生動物共生微生物定殖和增殖的適宜環境[31]。腸道也是消化吸收營養物質的主要器官, 是機體最大的“免疫器官”。腸道菌群是構建抵御病原體的第一道屏障, 促進腸道健康并確保腸道持續正常的生理功能。在門水平, 變形菌門、擬桿菌門、放線菌門和厚壁菌門在不同發育階段的蝦腸道中均占優勢。Tzeng等[32]比較了不同棲息環境日本沼蝦(Macrobrachium nipponense)腸道細菌組成, 發現主要為變形菌門, 其次是厚壁菌門和放線菌門。Rungrassamee等[33]研究發現不同發育階段的斑節對蝦腸道優勢菌都是變形菌門。郁維娜等[34]發現健康和患病的凡納濱對蝦腸道主要菌群均為變形菌門 (51.2%) 和放線菌門 (27.2%) 。Mente等[35]研究處于蛻殼不同階段羅氏沼蝦(Macrobrachiumrosenbergii)的腸道優勢菌主要為變形菌門, 其次為厚壁菌門。段健誠等[30]研究肝腸胞蟲病蝦及健康蝦腸道菌群, 優勢菌群也為變形菌門和厚壁菌門。本實驗研究得到健康對蝦與發病對蝦中腸道及肝胰腺中的優勢菌組成類似, 其中健康蝦與發病對蝦腸道中的優勢細菌均屬于變形菌門、厚壁菌門和擬桿菌門, 而其肝胰腺中的優勢菌門均為變形菌門。這一結果與其他蝦腸道優勢菌亦類似, 表明健康對蝦與發病對蝦中腸道與肝胰腺細菌群落具有高度的相似性。
而在屬水平上, 健康對蝦和發病對蝦的腸道及肝胰腺內優勢細菌的組成及豐度存在明顯不同, 健康對蝦腸道中的豐度最大的優勢菌屬為埃希桿菌-志賀菌屬、海綿菌屬; 而發病對蝦腸道中的則為希瓦式菌屬、巨球型菌屬和克雷伯氏菌屬。健康對蝦肝胰腺中最具優勢豐度的是假單胞菌屬、漢氏鹽單胞菌, 而發病對蝦肝胰腺中的則為貪銅菌屬。蔣葛等[36]研究凡納濱對蝦急性肝胰腺壞死綜合癥病蝦與健康蝦腸道優勢菌群, 發現不同生長階段病蝦腸道中的優勢菌屬均為弧菌屬(vibrio), 而健康蝦幼蝦腸道優勢菌屬為假單胞菌屬, 養殖中期的優勢菌屬為發光桿菌屬(Photobacterium), 養殖后期的優勢菌屬為弧菌屬; 而Jesús AntonioLópez-Carvallo等[37]研究發現蝦感染EHP后期, 肝胰腺豐度最高的菌屬為假單孢菌屬。同時研究表明[38], 肝腸胞蟲病會引起對蝦飲食及個體大小不同, 這些差異可能會影響蝦腸道微生物的變化。李彥芹等[39]曾進行了一項初步研究, 比較了攝食不同餌料對日本沼蝦腸道菌群的影響, 研究結果表明: 攝食人工飼料及小黃魚的日本沼蝦的腸道的優勢菌群均為鄰單胞菌屬(Plesiomonas)和假單胞菌屬, 且研究還發現攝食飼料后的腸道細菌數量與攝食飼料前相比呈先增后減的趨勢; 另外, 楊慧贊等[40]研究發現大蝦和小蝦在菌屬水平存在顯著差異, 大蝦的主要優勢菌屬為乳桿菌屬(Lactobacillus)和片球菌屬(Pediococcus)、蒼白桿菌屬(Ochrobactrum)、軍團桿菌屬(Legionella), 而小蝦的主要優勢菌屬為紅小梨形菌屬(Rhodopirellula)、布拉格菌屬(Pragia)和不動桿菌屬(Acinetobacter)。
3.3 腸道及肝胰腺菌群功能預測分析
蝦腸道內富含許多微生物, 其中包括一些能夠代謝蛋白質的細菌和酵母菌, 將蛋白質分解成小分子的氨基酸, 提供給對蝦進行吸收利用。本研究中Tax4Fun2預測結果顯示, 健康對蝦腸道菌群的基因功能與新陳代謝功能有關, 主要為氨基酸代謝。研究表明健康蝦腸道菌群代謝氨基酸可能通過合成蛋白質、酶和激素等功能來維持健康狀態并提供能量[41], 同時Li等[42]也做了相關研究, 證實了這一觀點。而發病對蝦的腸道菌群和肝胰腺菌群的基因功能主要與遺傳信息處理功能有關, 王金星等[43]研究發現相關基因和遺傳信息可以影響肝胰腺腸道菌群的組成和數量, 從而影響蝦類的免疫系統和代謝。健康對蝦腸道及肝胰腺菌群的基因功能和發病對蝦腸道及肝胰腺菌群之間的基因功能不同,導致它們對病原體的不同反應, 這為研究蝦類疾病的病理機制提供啟示, 并幫助開發更有效的防治策略。
綜上所述, 健康對蝦與發病對蝦間的腸道和肝胰腺中的優勢菌門具有高度相似性, 但優勢菌屬組成及豐度存在明顯不同。健康對蝦腸道和肝胰腺的菌群的主要功能與發病對蝦腸道和肝胰腺的主要功能存在明顯差異。本研究有助于了解肝腸胞蟲對凡納濱對蝦腸道及肝胰腺細菌群落的影響, 并為凡納濱對蝦的疾病管理和健康養殖提供依據。
猜你喜歡
我愛學·笑話與口才(2025年3期)2025-02-24 00:00:00
音樂探索(2022年2期)2022-05-30 21:01:37
趣味(語文)(2020年3期)2020-07-27 01:42:46
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
中國衛生(2015年2期)2015-11-12 13:13:54
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
中國火炬(2014年11期)2014-07-25 10:31:58
