
芒果AP2/ERF轉(zhuǎn)錄因子MiERF2基因的克隆及表達(dá)分析
2024-02-21 19:48:52魏玲寇明睿李雯
熱帶作物學(xué)報(bào) 2024年1期
魏玲 寇明睿 李雯



關(guān)鍵詞:芒果;ERF轉(zhuǎn)錄因子;生物信息學(xué)分析;亞細(xì)胞定位;表達(dá)分析
AP2/ERF轉(zhuǎn)錄因子超家族存在于眾多植物中,在植物生長發(fā)育方面具有重要功能,該家族蛋白特征明顯,含有典型的AP2/ERF結(jié)構(gòu)域,具有結(jié)合DNA的功能[1]。ERF最早是從煙草中分離發(fā)現(xiàn)的,功能域主要由4個(gè)部分構(gòu)成,依次為轉(zhuǎn)錄調(diào)控功能域、DNA結(jié)合功能域、寡聚化位點(diǎn)功能域和核定位信號(hào)功能域[1]。AP2/ERF轉(zhuǎn)錄因子與其他轉(zhuǎn)錄因子區(qū)分的主要依據(jù)是該家族成員均包含有1個(gè)或多個(gè)由60~70個(gè)高度保守的氨基酸組成的AP2/ERF結(jié)構(gòu)域,該結(jié)構(gòu)域是ERF行使轉(zhuǎn)錄調(diào)控的關(guān)鍵[2]。AP2/ERF家族轉(zhuǎn)錄因子根據(jù)其含有的結(jié)構(gòu)域數(shù)量和是否含有其他類型結(jié)構(gòu)域這2個(gè)依據(jù),將其分為4個(gè)亞族(AP2、DREB、ERF、RAV)和單獨(dú)成員(Soloist)[3-5]。AP2亞家族劃分的依據(jù)是氨基酸序列中含有2個(gè)串聯(lián)的相似度很高的AP2/ERF結(jié)構(gòu)域;DREB和ERF亞家族相似度很高,序列中均只含有1個(gè)AP2/ERF保守結(jié)構(gòu)域,主要區(qū)分依據(jù)是ERF亞家族第14位是丙氨酸,第19位是天冬氨酸,而DREB亞家族則分別是纈氨酸和谷氨酸;RAV亞家族中含有2個(gè)不同的結(jié)構(gòu)域,分別是AP2/ERF結(jié)構(gòu)域和B3結(jié)構(gòu)域,可以結(jié)合不同元件發(fā)揮多種調(diào)控作用;而Soloist亞家族中雖然也包含1個(gè)AP2/ERF結(jié)構(gòu)域,但與其他幾類亞家族的結(jié)構(gòu)域差異很大,對(duì)Soloist亞家族的研究報(bào)道也很少[3,6-7]。AP2亞家族成員的基因主要參與植物側(cè)根形成和開花等過程,ZHANG等[8]研究發(fā)現(xiàn)油菜BrAP2a和BrAP2b能夠修復(fù)擬南芥ap2-5突變體花冠缺陷。DREB亞家族可以參與調(diào)控植物干旱及低溫等非生物脅迫響應(yīng),田文等[9]的研究表明小麥TaDREB27-A-2等5個(gè)基因參與了小麥的熱脅迫反應(yīng);王雷立等[10]研究發(fā)現(xiàn)基于玉米轉(zhuǎn)錄水平分析結(jié)果顯示與脅迫響應(yīng)相關(guān)的DREB亞家族基因多數(shù)下調(diào)表達(dá)。ERF亞家族參與調(diào)控乙烯應(yīng)答機(jī)制,也參與植物非生物脅迫響應(yīng)。研究表明香蕉MaERF9與乙烯合成呈正相關(guān),并且可以通過調(diào)控乙烯合成促進(jìn)香蕉果實(shí)成熟[11],而獼猴桃AcERF1則與其果實(shí)成熟呈負(fù)相關(guān)[12]。RAV亞家族在植物開花、種子發(fā)育等方面起重要調(diào)控作用,研究表明擬南芥RAV1與種子發(fā)育和開花時(shí)間呈負(fù)相關(guān),此外,過表達(dá)AtRAV1還能延緩擬南芥?zhèn)雀蜕徸~的發(fā)育[13-14],而在擬南芥中過表達(dá)白菜NGA1則可以顯著抑制擬南芥根、花、子葉和葉片的發(fā)育[15]。
芒果是漆樹科芒果屬植物[16],果香沁人心脾,果皮色澤艷麗,果形飽滿,果肉細(xì)膩香甜,營養(yǎng)物質(zhì)豐富,維生素含量高,具有較高的營養(yǎng)價(jià)值,被譽(yù)為“熱帶果王”,深受人們喜愛。世界糧農(nóng)組織(FAO,2020年)統(tǒng)計(jì)數(shù)據(jù)顯示,全球芒果種植面積約為6.543×106hm2,位居世界第五,芒果產(chǎn)量約為5.502×107t,僅次于柑橘、香蕉、葡萄、蘋果,是熱帶地區(qū)主要的經(jīng)濟(jì)作物之一。芒果作為典型的呼吸躍變型水果,在貯藏及銷售過程中會(huì)釋放大量乙烯,使得果實(shí)容易腐爛,導(dǎo)致貯藏期短,不易保存,對(duì)芒果的商品價(jià)值造成影響,制約了芒果產(chǎn)業(yè)的發(fā)展。挖掘與芒果成熟衰老相關(guān)的AP2/ERF轉(zhuǎn)錄因子家族基因并對(duì)其功能進(jìn)行解析,為研究果實(shí)采后成熟衰老的調(diào)控機(jī)制提供理論依據(jù)。因此,本研究通過PCR擴(kuò)增技術(shù),以采后貴妃芒果為試材,克隆獲得1個(gè)AP2/ERF轉(zhuǎn)錄因子MiERF2基因,對(duì)其進(jìn)行生物信息學(xué)分析和亞細(xì)胞定位研究,并對(duì)其在不同采收成熟度下乙烯利處理后的基因表達(dá)量進(jìn)行檢測,為進(jìn)一步分析MiERF2轉(zhuǎn)錄因子在芒果采后成熟過程中的功能以及芒果貯藏保鮮研究提供理論依據(jù)。
1材料與方法
1.1材料
供試芒果品種為貴妃芒,采摘于海南省陵水黎族自治縣英州鎮(zhèn)萬豐村景鵬萬豐基地種植園。分2次采摘6成熟芒果(花后100d)和8成熟芒果(花后130d)。選取大小均勻一致、無機(jī)械損傷、無病蟲害的芒果,立即運(yùn)回實(shí)驗(yàn)室。芒果剪留0.5cm果柄,用0.1%的氯消凈清洗并浸泡20min,晾干,用400μg/mL乙烯利(ETH)溶液浸泡5min,報(bào)紙包裹密封于塑料桶中24h后取出晾干。以不做任何處理的芒果為對(duì)照,隨機(jī)選取10個(gè)芒果為1組,裝入保鮮袋中,貯藏在(251)℃,相對(duì)濕度85%~90%的培養(yǎng)箱中,每3d取1次樣,用液氮將所取樣品速凍后保存在?80℃超低溫冰箱備用。
1.2方法
1.2.1RNA提取與MiERF2基因全長克隆以芒果果肉為材料,提取RNA并逆轉(zhuǎn)錄成cDNA,果肉RNA提取采用改良的CTAB法[17],cDNA的合成使用逆轉(zhuǎn)錄試劑盒(MonScriptTMRTIIIAll-in-OneMixWithdsDNase)。利用DNAMAN6.0軟件設(shè)計(jì)MiERF2的特異性引物(MiERF2-F:5?ATGCAGAAGTCCTCCAAGCG-3,MiERF2-R:5?TTAAGAACTATGACAATTTCCAGTTG-3)。參照TOYOBO高保真酶說明書,利用PCR擴(kuò)增得到MiERF2的ORF序列,擴(kuò)增產(chǎn)物用1%瓊脂糖凝膠電泳檢測,并用膠回收試劑盒(中科瑞泰瓊脂糖凝膠DNA回收試劑盒)回收目標(biāo)條帶,得到純化產(chǎn)物。純化產(chǎn)物連接pMD19-T載體(TaKaRapMD19-TVectorCloningKit)并轉(zhuǎn)化大腸桿菌感受態(tài)(吐露港,E.coliDH5α),挑選陽性菌落送中美泰和生物技術(shù)(北京)有限公司測序。
1.2.2MiERF2生物信息學(xué)分析利用ExPASyProtParam、NetPhos3.1Server、SignalP-4.1、TMHMMServerv.2.0等軟件分析蛋白理化性質(zhì)、磷酸化位點(diǎn)、信號(hào)肽及跨膜結(jié)構(gòu);利用SMART、SOPMA、SWISS-MODEL軟件分析MiERF2蛋白的結(jié)構(gòu);利用DNAMAN6.0軟件進(jìn)行多序列比對(duì);利用MEME5.5.0分析motif結(jié)構(gòu),motif數(shù)設(shè)為10;利用MEGA11.0.8軟件采用鄰接法構(gòu)建系統(tǒng)進(jìn)化樹。
1.2.3亞細(xì)胞定位采用洋蔥內(nèi)表皮轉(zhuǎn)化法進(jìn)行亞細(xì)胞定位分析[18],略有修改。采用雙酶切法構(gòu)建到PCAMBIA1302-GFP(35S::GFP)載體上,生成35S::MiERF2-GFP融合表達(dá)載體,酶切位點(diǎn)選用NcoI和BglII,將重組質(zhì)粒轉(zhuǎn)化GV3101農(nóng)桿菌制備OD600為0.8~1.0的懸浮液,將洋蔥內(nèi)表皮(1.5cm1.5cm)浸泡在懸浮液中減壓10min,用濾紙吸去洋蔥內(nèi)表皮上多余的菌液,將其轉(zhuǎn)移到鋪有濾紙的MS固體培養(yǎng)基上,放置在25℃培養(yǎng)箱中避光培養(yǎng)2d后制成玻片,在熒光生物攝像顯微鏡下觀察熒光信號(hào)。
1.2.4MiERF2的表達(dá)分析利用DNAMAN6.0軟件設(shè)計(jì)定量引物(qMiERF2-F:5-CGCAACAACATTAACGAGTCA-3,qMiERF2-R:5-CCACTTTCCCTCCCAAATTCA-3),內(nèi)參基因?yàn)锳ctin(Actin-F:5-ATCTGCTGGAAGGTGCTGAG-3,Actin-R:5-CCAAGCAGCATGAAGATCAA-3),使用試劑盒(MonAmpTMChemoHSqPCRMix)進(jìn)行RT-qPCR擴(kuò)增,利用2?ΔΔCT法計(jì)算基因的相對(duì)表達(dá)量。
1.3數(shù)據(jù)處理
使用WPSOffice2022軟件進(jìn)行數(shù)據(jù)統(tǒng)計(jì)與計(jì)算,使用SPSS26軟件進(jìn)行差異顯著性分析,使用GraphPadPrism8進(jìn)行相對(duì)表達(dá)量作圖。
2結(jié)果與分析
2.1MiERF2基因的克隆及其蛋白的理化性質(zhì)分析
克隆得到1個(gè)開放閱讀框(ORF)長度為915bp的基因,該基因編碼305個(gè)氨基酸,命名為MiERF2。對(duì)MiERF2蛋白理化性質(zhì)進(jìn)行預(yù)測,結(jié)果顯示,MiERF2蛋白分子式為C1449H2259N433O470S11,分子量為33618.16Da,總原子數(shù)為4622,理論等電點(diǎn)為7.66。由氨基酸組成可以得知,帶負(fù)電殘基數(shù)量(Asp+Glu)為29個(gè),帶正電殘基數(shù)量(Arg+Lys)為30個(gè),脂肪族氨基酸指數(shù)為52.66;親水性總平均值(GRAVY)為?0.796,為親水蛋白;不穩(wěn)定性指數(shù)為47.13,為不穩(wěn)定蛋白。
2.2MiERF2蛋白結(jié)構(gòu)分析
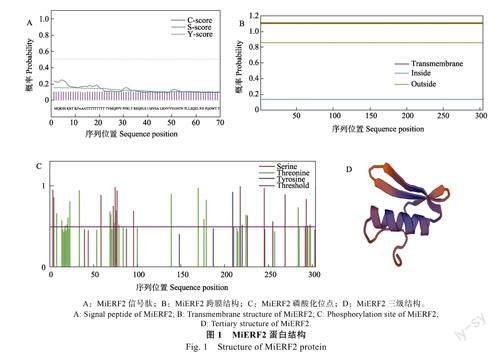
預(yù)測結(jié)果顯示,MiERF2蛋白中不存在信號(hào)肽(圖1A),也并未發(fā)現(xiàn)跨膜區(qū)域(圖1B)。MiERF2蛋白的磷酸化可能發(fā)生在絲氨酸(Ser)、蘇氨酸(Thr)和酪氨酸(Tyr)位點(diǎn)上(圖1C),主要為蘇氨酸位點(diǎn),說明此蛋白發(fā)揮調(diào)控功能可能是以蘇氨酸位點(diǎn)的磷酸化修飾為主。MiERF2包含1個(gè)AP2保守結(jié)構(gòu)域,屬于AP2/ERF家族轉(zhuǎn)錄因子,其二級(jí)結(jié)構(gòu)以無規(guī)則卷曲為主,占比為66.12%,α-螺旋占比為24.67%,延長鏈占比為7.89%,β-折疊占比為1.32%。蛋白的三級(jí)結(jié)構(gòu)模型中包含有3個(gè)β折疊和1個(gè)α-螺旋(圖1D),此為AP2結(jié)構(gòu)域的典型三維結(jié)構(gòu)。
2.3MiERF2蛋白motif分析及同源序列比對(duì)
選取9個(gè)與MiERF2同源性較高的ERF轉(zhuǎn)錄因子進(jìn)行motif分析,結(jié)果顯示,所有motif結(jié)構(gòu)相似,有4種motif在所有蛋白中都含有,與MiERF2所含的motif數(shù)目和種類一致的有3種,分別為阿月渾子、木油桐、皴皮婆羅雙(圖2A)。氨基酸同源序列比對(duì)結(jié)果顯示,其氨基酸序列中僅含有1個(gè)AP2保守結(jié)構(gòu)域,且在該結(jié)構(gòu)域中的第14位和第19位氨基酸為丙氨酸(A)和天冬氨酸(D)(圖2B)。上述結(jié)果表明MiERF2蛋白屬于ERF亞家族成員。
2.4MiERF2蛋白質(zhì)的系統(tǒng)進(jìn)化分析
選取漆樹科植物、模式植物、熱帶植物等不同物種ERF基因構(gòu)建進(jìn)化樹,結(jié)果顯示,芒果MiERF2與同為漆樹科的阿月渾子ERF109-like聚在一起,親緣關(guān)系最近,其次是橄欖、枳和甜橙,而熱帶植物番木瓜、菠蘿以及模式植物番茄則與芒果MiERF2親緣關(guān)系較遠(yuǎn)(圖3)。
2.5亞細(xì)胞定位結(jié)果分析
35S::GFP對(duì)照洋蔥內(nèi)表皮細(xì)胞中的細(xì)胞核和細(xì)胞壁均檢測到了綠色熒光信號(hào),而35S::MiERF2-GFP融合蛋白的綠色熒光信號(hào)僅出現(xiàn)在細(xì)胞核中(圖4),這表明芒果MiERF2蛋白在洋蔥內(nèi)表皮細(xì)胞中定位于細(xì)胞核,符合轉(zhuǎn)錄因子所具有的核定位特征。
2.6MiERF2基因的表達(dá)特征分析
在乙烯利處理下不同采收成熟度中的MiERF2基因表達(dá)結(jié)果顯示,在6成熟芒果中MiERF2基因的表達(dá)量總體呈現(xiàn)先上調(diào)后下調(diào)的趨勢,對(duì)照組在15d達(dá)到最大值,而ETH處理組在9d時(shí)達(dá)到最大值后迅速下降,且處理組中MiERF2基因的表達(dá)量只在6d時(shí)顯著高于對(duì)照組,在12d后均顯著低于對(duì)照組;在8成熟芒果中對(duì)照組MiERF2基因的表達(dá)量呈現(xiàn)先上調(diào)后下調(diào)的趨勢,而ETH處理組呈波動(dòng)趨勢,對(duì)照組在3d達(dá)到最大值,處理組則不明顯,相較于對(duì)照組,除18d外,處理組MiERF2基因的表達(dá)水平在整個(gè)貯藏期中始終顯著低于對(duì)照組(圖5)。MiERF2基因在不同成熟度中表達(dá)模式不同,在對(duì)照和處理中存在顯著差異,表明MiERF2基因可能在芒果采后成熟衰老過程中發(fā)揮負(fù)調(diào)控作用。
3討論
新的研究發(fā)現(xiàn)藍(lán)藻、線蟲和病毒中也存在著AP2結(jié)構(gòu)域[19],但AP2/ERF轉(zhuǎn)錄因子在植物中的研究最為廣泛。AP2/ERF轉(zhuǎn)錄因子能夠識(shí)別和結(jié)合多種不同的順式作用元件,從而發(fā)揮調(diào)控作用。ZHAO等[20]研究發(fā)現(xiàn)了1個(gè)水稻ERF3基因,OsERF3基因能夠促進(jìn)水稻根冠的形成;騰飛[21]研究發(fā)現(xiàn)影響擬南芥合子胚發(fā)育的OsERF055基因能導(dǎo)致胚胎發(fā)育滯后,從而無法形成正常的合子胚。AP2/ERF轉(zhuǎn)錄因子還能加速葉片衰老[22]、加速果實(shí)褪綠[23]、防止果實(shí)提前軟化[24]以及果實(shí)木質(zhì)化[25]等。目前已經(jīng)在許多物種中鑒定到AP2/ERF轉(zhuǎn)錄因子,如在蘋果中鑒定了259個(gè)ERFs[26];桃中鑒定了131個(gè)ERFs[27];柑橘中鑒定了126個(gè)[28]。ERF亞家族是AP2/ERF超家族中最大的一個(gè)家族,在乙烯信號(hào)轉(zhuǎn)導(dǎo)的最后一步作為反式作用因子發(fā)揮著重要調(diào)控作用,它可以結(jié)合啟動(dòng)子中多種順式元件,如DRE/CRT、GCC-box、ERE和ATCTA元件[29-31]。易萍等[32]在臺(tái)農(nóng)1號(hào)芒果中鑒定了1個(gè)MiERF113基因,該基因?qū)儆贓RF亞家族成員,其蛋白不存在信號(hào)肽和跨膜結(jié)構(gòu),與阿月渾子親緣關(guān)系最近,該研究與本研究結(jié)果相似。本研究鑒定的MiERF2基因也屬于ERF亞家族,蛋白具有親水性,性質(zhì)不穩(wěn)定,不存在信號(hào)肽,也沒有跨膜結(jié)構(gòu);MiERF2蛋白的二級(jí)和三級(jí)結(jié)構(gòu)預(yù)測其包含α-螺旋、β-折疊、延長鏈及無規(guī)則卷曲,以無規(guī)則卷曲為主,并且具有AP2結(jié)構(gòu)域的典型三維結(jié)構(gòu),系統(tǒng)進(jìn)化分析發(fā)現(xiàn)其與同為漆樹科的阿月渾子ERF109-like親緣關(guān)系最近。
果實(shí)采收成熟度是影響果實(shí)品質(zhì)的主要因素之一,采收偏早,易造成果實(shí)品質(zhì)和風(fēng)味欠佳;采收偏晚,易使果實(shí)硬度下降,運(yùn)輸效果和耐貯性變差。高豪杰等[33]研究結(jié)果顯示1-MCP處理同一品種芒果后,相比采收成熟度低的果實(shí),采收成熟度高的果實(shí)病情指數(shù)高、色度值高、果實(shí)硬度低,不耐貯藏。生產(chǎn)上往往采用乙烯利進(jìn)行催熟,乙烯利處理會(huì)促進(jìn)果實(shí)成熟,加速果實(shí)變軟,使果實(shí)硬度降低,如甜瓜[34]等,因此本研究選取了6成熟和8成熟2種采收成熟度芒果,分別用乙烯利進(jìn)行處理,以研究MiERF2基因的表達(dá)模式。而陳明敏等[35]檢測了6個(gè)芒果ERF轉(zhuǎn)錄因子在不同濃度乙烯利處理下的表達(dá)情況,發(fā)現(xiàn)處理后有5個(gè)基因表達(dá)量高于對(duì)照,1個(gè)基因表達(dá)量低于對(duì)照;陳永萍等[36]通過番木瓜轉(zhuǎn)錄組分析發(fā)現(xiàn),差異表達(dá)的ERF家族成員基因中,乙烯利處理后有5個(gè)ERF轉(zhuǎn)錄因子表達(dá)量高于對(duì)照,2個(gè)ERF轉(zhuǎn)錄因子的表達(dá)量低于對(duì)照,與本研究結(jié)果相似。本研究鑒定的MiERF2基因定量結(jié)果表明,在不同成熟度中表達(dá)模式并不相同,且MiERF2的表達(dá)量在乙烯利處理后與對(duì)照相比差異顯著。在6成熟芒果貯藏后期,對(duì)照組MiERF2基因表達(dá)量顯著高于處理組,在8成熟芒果采后貯藏過程中,對(duì)照組MiERF2的表達(dá)量相對(duì)較高,處理組MiERF2的表達(dá)量則一直處于較低水平。上述結(jié)果表明MiERF2可能在芒果成熟過程中發(fā)揮著負(fù)調(diào)控作用,而關(guān)于MiERF2基因后續(xù)的功能驗(yàn)證還有待更深入的研究。
