
TCS1等位基因的差異表達參與弄島野茶低咖啡堿的形成
2024-02-21 19:48:52劉玉飛龐丹丹陳春林田易萍孫云南李友勇陳林波
熱帶作物學報 2024年1期
劉玉飛 龐丹丹 陳春林 田易萍 孫云南 李友勇 陳林波


關(guān)鍵詞:弄島野茶;特異資源;TCS1;生物合成
茶樹富含咖啡堿、可可堿、茶葉堿等多種嘌呤生物堿,其中咖啡堿(2.5%~4.5%)是其含量較高并且較常見的嘌呤生物堿[1]。咖啡堿具有興奮、提高大腦記憶力和識別能力等多種對人體有利的生理功能[2],但過量的攝入會使敏感人群出現(xiàn)失眠[3],并且有可能引起骨質(zhì)疏松和流產(chǎn)等不利反應(yīng)[4-5]。因此,低(無)咖啡堿茶以及相關(guān)的茶制品(咖啡堿含量低于1%)受到了有特殊需求消費群體的喜愛[6]。目前常用的脫咖啡堿技術(shù)主要有溶劑萃取、物理吸附法、生物降解法以及培育低咖啡因茶樹品種等[7-8],其中低咖啡堿茶樹育種是實現(xiàn)茶葉低(無)咖啡堿最經(jīng)濟、安全和有效的方法[8]。咖啡堿代謝途徑的解析以及低咖啡堿特異茶樹資源的鑒定有利于茶樹低咖啡堿育種工作的開展。
咖啡堿的生物合成途徑已經(jīng)明確,其可以通過從頭合成嘌呤環(huán),然后再合成咖啡堿,也可以利用已有的嘌呤類化合物為基礎(chǔ)進行合成。NEGISHI等[9]根據(jù)同位素示蹤研究,提出了茶樹咖啡堿生物合成的核心途徑,即以黃嘌呤核苷(xanthosine,XR)為底物,經(jīng)7-甲基黃嘌呤核苷、7-甲基黃嘌呤和可可堿等中間產(chǎn)物而合成,合成途徑涉及N-甲基轉(zhuǎn)移酶類(N-methyltransferases,NMTs)、N-甲基核苷酶和S-腺苷-L-甲硫氨酸合成酶(S-adenosyl-L-methioninesynthetase,SAMS)。咖啡堿的合成涉及到多種酶類基因,如SAMS(AB041534)[10]、TIDH(inosine-5'-monophosphatedehydrogenase,EU106658)[11]和咖啡堿合成酶TCS(caffeinesynthase,AB031280)[12]等。
TCS是茶樹咖啡堿代謝最為關(guān)鍵的基因,屬于NMT家族,KATO等[12]通過克隆獲得了茶樹第一個NMT,茶樹咖啡堿合成酶基因TCS1,接著YONEYAMA等[13]在茶樹中克隆與TCS1序列相似大于89%的TCS2(AB031281),隨后金基強等[14]根據(jù)TCS1序列從茶樹基因組中克隆得到4種TCS基因(TCS3~TCS6)。前期的功能鑒定與進化分析結(jié)果表明,TCS1是咖啡堿合成途徑中的關(guān)鍵基因,對茶樹合成咖啡堿最為重要[12-14]。多項研究表明TCS1具有多種等位變異,現(xiàn)已報道8種TCS1的等位基因,分別命名為TCS1a~TCS1g和TCS1i,其中TCS1a、TCS1b、TCS1c與之前克隆的TCS1、ICS1、PCS1序列相同[15-17]。前期的進化分析以及酶學實驗表明TCS1的不同等位變異可以分為2類,一類僅具有可可堿合成酶活性(TCS1b、TCS1c、TCS1g和TCS1i);另一類同時具有可可堿和咖啡堿合成酶活性(TCS1a、TCS1d、TCS1e和TCS1f)[15,17]。
本課題組前期從國家大葉茶樹資源圃(勐海)篩選獲得1份低咖啡堿、高可可堿茶樹種質(zhì)——弄島野茶(MHLC),然而,MHLC低咖啡堿、高可可堿的形成原因尚不清楚。因此,本研究通過對MHLC和云抗10號(YK10,對照品種)不同季節(jié)以及MHLC不同部位嘌呤生物堿的含量進行分析,明確MHLC中的化學成分,并通過基因型鑒定和基因表達量分析初步解析MHLC低咖啡堿的形成機制。
1材料與方法
1.1材料
采摘弄島野茶(MHLC)、云抗10號(YK10)(圖1)春、夏、秋的一芽二葉(春季:3月27日;夏季:7月10日;秋季:10月27日),并收集MHLC新梢不同成熟度葉片和莖的樣品(采于10月27日),微波固樣2min,然后80℃烘至足干,一部分用于生化成分的測定,另一部分液氮固樣,并置于–80℃冰箱用于后期RNA的提取。
1.2方法
1.2.1化學組分提取與測定準確稱取0.1g(精確到0.001)樣品,加入10mL70%的甲醇溶液,搖勻,然后70℃水浴10min,期間搖勻2次,提取結(jié)束后7000r/min離心1min,取上清液過0.45μm有機濾膜,用HPLC測定樣品中的生物堿,具體方法參見JIN等[15]的研究。
1.2.2實時熒光定量PCR以茶樹GAPDH基因作為內(nèi)參基因測定MHLC和YK10中嘌呤生物堿關(guān)鍵基因SAMS、TIDH和TCS的表達量,其中SAMS、TIDH分別是N-甲基核苷酶和S-腺苷-L-甲硫氨酸合成酶基因、肌苷-5′-單磷酸脫氫酶基因;TCS1-CS、TCS1-TS、TCS1-Total分別是表達蛋白同時具有可可堿、咖啡堿合成酶(caffeinesynthase,CS)活性的等位變異、表達蛋白僅具有可可堿合成酶(theobrominesynthase,TS)活性的等位變異以及TCS1整體表達水平的引物。引物序列來源于JIN等[15]的研究(表1),RT-qPCR具體方法參照PANG等[18]的研究。
1.2.3TCS1等位變異cDNA全長的克隆利用TCS1(TCS1a,AB031280)的序列設(shè)計引物克隆不同樣品中的TCS1基因,具體的基因克隆方法見PANG等[18]的研究。利用ClustalW(https://www.genome.jp/tools-bin/clustalw)在線軟件進行序列比對,并利用EXPASY中的BoxShade(https://embnet.vital-it.ch/software/BOX_form.html)輸出同源比對的結(jié)果。
2結(jié)果與分析
2.1不同季節(jié)化學組分含量
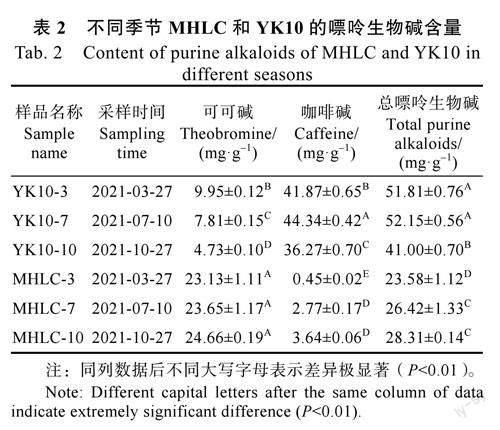
MHLC和對照品種YK10三個季節(jié)(春、夏、秋)的可可堿與咖啡堿測定結(jié)果顯示,MHLC與YK10的嘌呤生物堿的總量和各組分含量均存在顯著差異(表2)。MHLC生物堿總量和咖啡堿含量均顯著低于YK10,其中咖啡堿含量不足YK10的九分之一,但是其可可堿含量要高于YK10;MHLC中含量最高嘌呤生物堿是可可堿(大于23.00mg/g),其次是咖啡堿(小于4.00mg/g),而YK10與之相反。另外,春季到秋季MHLC中的可可堿和咖啡堿含量均有增加的趨勢,但是除春季MHLC的咖啡堿極顯著低外,其他差異均未達到極顯著水平;而YK10中的可可堿則一直降低,咖啡堿先升高后降低。
2.2MHLC不同組織化學組分含量
為了明確MHLC嘌呤生物堿不同發(fā)育階段的變化規(guī)律,本研究測定了MHLC芽(MHLC-B)、不同發(fā)育階段的葉片(芽下第一片葉子用MHLC-1L表示,其它依次類推)和莖(芽至第一片葉子之間的莖用MHLC-1S表示,第一片葉子至第二片葉子之間的莖用MHLC-2S表示,其它依次類推)的嘌呤生物堿。結(jié)果顯示,MHLC芽中具有最高的可可堿(32.16mg/g)和咖啡堿(6.14mg/g)含量,同一發(fā)育階段葉片中可可堿和咖啡堿含量高于莖,另外,隨著葉片和莖成熟度的增加,可可堿、咖啡堿以及嘌呤生物堿總量均逐漸下降(圖2)。
2.3咖啡堿合成酶基因的克隆
根據(jù)TCS1(TCS1a,AB031280)的基因序列設(shè)計引物,成功從MHLC和YK10中克隆得到TCS1基因的編碼區(qū)全長。在YK10中克隆到1個TCS1的等位變異,命名為YK10_TCS1,其與TCS1a(具有可可堿和咖啡堿合成酶活性)有3個核苷酸的差異,雖然它們引起了編碼氨基酸殘基的變化,但是均不影響TCS的保守基序和關(guān)鍵活性位點。在MHLC中克隆得到2種TCS1的等位變異,分別命名為MHLC_TCS1b和MHLC_TCS1d。其中MHLC_TCS1b與已經(jīng)報道的僅具有可可堿合成酶活性的TCS1b(AB056108)雖然有3個核苷酸的差異,但是編碼蛋白序列完全相同;MHLC_TCS1d與已經(jīng)報道的具有可可堿和咖啡堿合成酶活性的TCS1d(KT215399),有1個核苷酸的差異,并且引起編碼氨基酸殘基的變化,但并不影響TCS的保守基序和關(guān)鍵活性位點(圖3)。
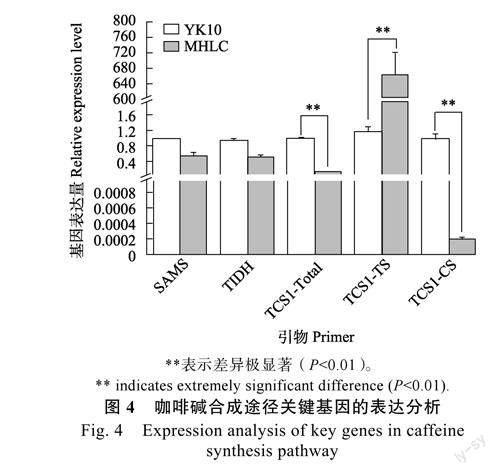
2.4咖啡堿合成途徑關(guān)鍵基因的表達分析
3討論
我國具有豐富的茶樹資源,低咖啡堿茶樹資源已在廣東[19]、福建[16,20]、四川[6]、云南[21-22]等多個省份發(fā)現(xiàn)。已報道的云南省低咖啡堿茶樹來源于文山壯族苗族自治州[21]、西雙版納傣族自治州[22-23]以及昭通市[24],本研究報道的低咖啡堿(不足4.00mg/g)和高可可堿(大于23.00mg/g)的弄島野茶(MHLC)來源于云南省德宏傣族景頗族自治州瑞麗市,是該地區(qū)首次報道的低咖啡堿茶樹資源。瑞麗市MHLC為低(無)咖啡堿茶樹育種提供了可利用的親本材料,并擴大了篩選低咖啡堿茶樹資源的區(qū)域。
MHLC中可可堿和咖啡堿由春季到秋季均有增加的趨勢,但除春季咖啡堿顯著低外,可可堿或咖啡堿不同季節(jié)間的差異均未達到極顯著水平;而YK10中可可堿從春季到秋季則一直降低,咖啡堿先升高后降低。這說明不同茶樹種質(zhì)資源,咖啡堿和可可堿不同季節(jié)間的變化趨勢有所差異,李金[25]對25個茶樹品種的研究結(jié)果也指出,不同品種在不同季節(jié)咖啡堿和可可堿的變化趨勢具有一定的差異,這可能與茶樹品種自身的調(diào)控和代謝機制相關(guān)。茶樹中咖啡堿和可可堿主要分布在幼嫩部位,其中在新梢部分的分布最為集中,越老化的葉片含量越少[16,26-27]。MHLC中咖啡堿和可可堿在不同成熟度葉片和莖中的分布規(guī)律與之前的結(jié)果一致,也是在幼嫩的部位含量較高。
本研究發(fā)現(xiàn),MHLC中含有2種TCS1等位變異基因:MHLC_TCS1b和MHLC_TCS1d。其中MHLC_TCS1b與已經(jīng)報道的TCS1b[15](AB056108,僅具有可可堿合成酶活性)編碼蛋白序列基本完全相同;雖然MHLC_TCS1d與TCS1d[15-16](KT215399,同時具有可可堿和咖啡堿合成酶活性)的編碼蛋白序列有1個氨基酸殘基的差異,但是并不影響NMT保守基序和TCS的關(guān)鍵活性位點,推測MHLC_TCS1d與TCS1d具有相似的功能。以上分析可知,MHLC同時具有TS活性和CS活性的TCS1等位變異。基因表達量分析發(fā)現(xiàn),MHLC_TCS1b在MHLC中的表達量很高,這與MHLC相比YK10在芽葉中積累較多的可可堿相符;而MHLC_TCS1d在MHLC中表達量極低(不足YK10中TCS1d基因表達量的1/4800),很可能是MHLC低咖啡堿含量的一個主要原因。JIN等[15]和劉玉飛等[28]的研究均表明TCS1不同等位變異的啟動子序列存在顯著差異,但MHLC_TCS1d的低表達是否由啟動子序列變異引起還需進一步研究。
