谷氨酰胺代謝相關蛋白在腫瘤轉移中的作用研究進展
2024-02-28 10:23:00劉雪柔楊玉梅榮翔宇鄭瑞潔龐金龍李姍姍
中國癌癥雜志 2024年1期
關鍵詞:肝癌
劉雪柔,楊玉梅,趙 倩,榮翔宇,劉 偉,鄭瑞潔,龐金龍,李 嫻,李姍姍
1.蚌埠醫科大學藥學院,安徽 蚌埠 233030;
2.安徽省生化藥物工程技術研究中心,安徽 蚌埠 233030
腫瘤細胞的轉移是由基因和蛋白表達或功能的改變引起的分子事件[1],包含一系列復雜的動態過程,高侵襲性的腫瘤具有浸潤性,能夠向周圍間質擴散,經過血液循環尋找遠處轉移的部位,且在轉移過程中還需適應營養供給和氧化應激的動態變化[2],之后在轉移部位進一步地增殖。癌組織與癌旁組織相比,代謝發生了顯著變化,且代謝變化成為驅動腫瘤轉移的主要因素。谷氨酰胺是人體血清中含量最豐富的非必需氨基酸,并在腫瘤發生、發展過程中為癌細胞提供碳源和氮源,用于癌細胞的生物合成,為癌細胞提供能量,促進腫瘤的生長。有研究[3]表明,谷氨酰胺代謝在腫瘤細胞轉移過程中發揮重要作用。本文綜述近年來谷氨酰胺在腫瘤轉移中作用及其機制的研究進展,并提出潛在的治療靶點,以期為臨床治療腫瘤轉移提供新的策略。
1 谷氨酰胺代謝概述
谷氨酰胺通過谷氨酰胺酶(glutaminase,GLS)生成谷氨酸和游離氨,接著谷氨酸經谷氨酸脫氫酶(glutamine dehydrogenase,GDH)催化生成α-酮戊二酸(α-ketoglutaric acid,α-KG),隨后α-KG可進入三羧酸循環(tricarboxylic acid,TCA)參與糖代謝[4]。因此,谷氨酰胺可作為碳源補充TCA。此外,谷氨酰胺可經完全氧化生成腺苷三磷酸(adenosine triphosphate,ATP),作為氮源合成核苷酸和其他非必需氨基酸[5-6]。其代謝產物谷氨酸可作為合成谷胱甘肽的原料之一,與NADPH一起作為還原劑參與維持細胞內氧化還原穩態,利于細胞正常增殖(圖1)。谷氨酰胺代謝重編程可作為腫瘤的代謝特征之一,在惡性增殖的腫瘤細胞中,谷氨酰胺代謝具有成癮性,在沒有外源性谷氨酰胺存在時無法存活[7]。谷氨酰胺代謝參與腫瘤細胞的生物前體合成,維持腫瘤細胞氧化還原穩態,并為腫瘤細胞提供能量,從而促進細胞增殖、遷移和侵襲。與谷氨酰胺代謝相關的酶,如GLS1/2[8]、谷氨酰胺合成酶(glutamine synthetase,GS)[9]、GDH[10]和谷氨酰胺轉氨酶(glutamine transaminase,TG)[11],以及參與谷氨酰胺轉運的蛋白質轉運體,如SLC1A5[12]、SLC38A2[13]和SLC38A3[14]等,在腫瘤細胞內部或腫瘤微環境(tumor microenvironment,TME)中發揮調控腫瘤轉移的作用。
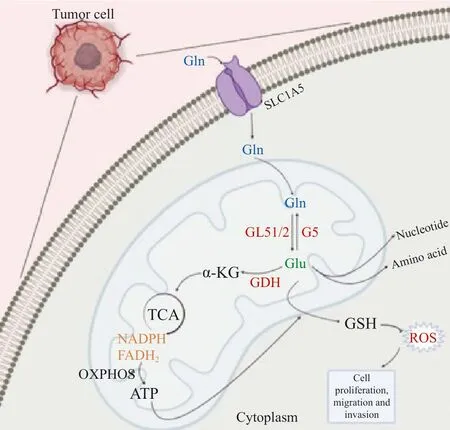
圖1 谷氨酰胺代謝的主要途徑Fig.1 The main pathway of glutamine metabolism
2 谷氨酰胺代謝影響腫瘤轉移的作用及機制
2.1 GLS與腫瘤轉移
GLS是一種線粒體酶,分為GLS1和GLS2兩種亞型,GLS1是一種線粒體基質蛋白,在轉錄階段通過不同的剪切方式又可分為GAC(肝型)和KAG(腎型)兩種亞型,GAC只存在于線粒體中,通過對腫瘤代謝物水平的調控參與腫瘤轉移,而KAG主要在細胞質中發揮作用,KAG主要是通過介導蛋白質-蛋白質的相互作用,可能以非酶促的方式參與調控,因此GLS1在參與腫瘤細胞的代謝調控時,GAC亞型的表達抑制或促進在腫瘤代謝功能改變中占據主要地位。GLS1和GLS2雖然都能催化谷氨酰胺生成谷氨酸和游離的氨,但兩者在腫瘤中的表達活性有明顯差別。
GLS1在腫瘤中表現出高活性,該蛋白表達與腫瘤進展呈正相關[15]。GLS1在肝癌、胃癌、結直腸癌(colorectal cancer,CRC)、食管癌及肺癌等多種癌癥中表達上調,促進腫瘤細胞增殖、侵襲和遷移,使用GLS1抑制劑BPTES、CB-893或經siRAN沉默形成siGLS1后,GLS1在腫瘤細胞中的表達活性減弱,可明顯抑制腫瘤細胞遷移。最近研究[16-17]發現,在甲狀腺乳頭狀癌中,GLS1高表達,雷帕霉素復合物1介導的信號通路被激活,ULK1被磷酸化失活,從而抑制細胞自噬,促進腫瘤細胞轉移。多數情況下,細胞自噬水平增加不利于腫瘤發展,但是在腫瘤細胞后期,細胞自噬也可以通過清除破損細胞器,維持細胞內環境穩定來應對氧化應激,有利于腫瘤細胞生存,例如,在CRC中,GLS1高表達導致腫瘤細胞自噬水平增加,促進腫瘤進展,而GLS1缺失導致細胞自噬水平下降,與此同時細胞核中Nrf2蛋白表達降低,共同導致細胞內活性氧(reactive oxygen species,ROS)水平升高,從而抑制腫瘤轉移,Nrf2蛋白作為一種轉錄因子通過調節下游的抗氧化基因來維持氧化還原平衡[18]。此外,在CRC中,GLS1可以作為缺氧誘導因子-1(hypoxia-inducible factor-1,HIF-1)的一個靶基因,HIF-1可與GLS1基因組中的一個位點結合,激活GLS1的表達來調節谷氨酰胺代謝,當GLS1缺乏可抑制缺氧誘導的肺定植和淋巴結轉移。還有研究[19]顯示,GLS1可與乳腺癌中的circHECTD1相互作用,促進腫瘤生長和轉移。GLS1還可以通過增加ROS積累和抑制Wnt/β-catenin信號通路減弱干細胞特性,從而減少癌癥干細胞(cancer stem cell,CSC)表型,CSC是一種能夠自我更新并分化為其他不同類型的腫瘤細胞,具有干細胞的特性,致瘤潛能大[20]。因此癌癥中存在CSC被認為是導致腫瘤復發、轉移和獲得化療耐藥性的重要原因[21],阻礙CSC表型形成能抑制腫瘤細胞增殖和遷移。
與GLS1不同的是,GLS2在多數情況下被認為是一種腫瘤抑制因子,且在腫瘤中的表達活性較低[22],其表達水平與腫瘤進展呈負相關。如在肝癌轉移研究[23-24]中發現,GLS2的敲除導致肝癌細胞系發生肺轉移的概率顯著增加,GLS2主要是通過抑制小鳥苷三磷酸酶(guanosine triphosphatase,GTPase)Rac1的活性進而抑制肝癌細胞遷移和侵襲。同樣有研究[25]報道,在肝癌中,GLS2表達通過穩定Dicer活性,誘導miR-34a的表達,接著降低snail的表達從而抑制上皮-間充質轉化(epithelial-mesenchymal transition,EMT),進而抑制腫瘤轉移。另外,在乳腺癌中,GLS2過表達可通過EGF-ERK-ZEB1-vimentin軸抑制EMT,從而抑制腫瘤進展[26]。
EMT與腫瘤的侵襲、轉移及耐藥密切相關,是腫瘤轉移的起始過程也是關鍵步驟[27]。在此階段,腫瘤上皮細胞喪失極性,成為間充質細胞,且更具有運動性,加速腫瘤細胞向遠處轉移,并使腫瘤獲得CSC的特性[28]。EMT受到多種轉錄因子的調控,包括snail[29]、ZEB[30]、Slug[31]和TWIST[32]等。已有研究[33]證明,在胰腺癌細胞中,谷氨酰胺缺失引起的MEK/ERK信號通路和ATF4轉錄因子的激活能誘導Slug蛋白表達水平的增加,促進EMT,增強侵襲性,而抑制Slug蛋白表達,可以降低胰腺癌細胞侵襲和轉移。
同樣也有研究[34]證明,GLS2在乳腺癌、食管鱗狀細胞癌(esophageal squamous cell carcinoma,ESCC)中作為致癌基因而存在,例如,甲基轉移酶樣因子3(methyltransferase like 3,METTL3)通過增強GLS2的表達促進ESCC的轉移,下調GLS2可以減弱METTL3介導的腫瘤侵襲和遷移。與GLS1在多種癌癥中的作用水平類似,GLS2的高水平表達與乳腺癌患者的不良預后相關。
2.2 GS與腫瘤轉移
GS催化細胞內谷氨酸和游離氨生成谷氨酰胺,因此在氮代謝、氨解毒方面發揮作用,GS可催化谷氨酰胺分解代謝逆過程,作為唯一從頭合成谷氨酰胺的酶,可以在細胞內通過合成谷氨酰胺來滿足腫瘤細胞快速增殖的需求,因此在癌癥轉移中至關重要[35]。GS在膠質瘤[36]、卵巢癌[37]和乳腺癌[38]等癌癥中表達上調,促進多種腫瘤的生長、增殖過 程。
GS在TME的調控中發揮重要作用,TME包括存在于腫瘤細胞外的細胞外基質、成纖維細胞、巨噬細胞、脂肪細胞及T淋巴細胞等,還包括血管的生成[39]。有研究[40]顯示,腫瘤相關巨噬細胞的表型轉化在多種類型腫瘤轉移中發揮重要作用,并表現出EMT表型。肝癌細胞產生的抗炎因子促進巨噬細胞的M2型極化,促進腫瘤轉移[41]。CRC細胞中,巨噬細胞同時存在M1和M2兩種表型,其中M2表型發揮主要作用,導致CRC細胞侵襲、遷移和轉移能力增強[42]。最近的研究[43]表明,GS活性下降能夠介導腫瘤相關巨噬細胞表型轉化,抑制腫瘤轉移。
當巨噬細胞處于M0表型時,在炎癥因子干擾素和其他激動劑的作用下轉變為M1表型,M1表型表現出炎癥反應,抑制血管生成,妨礙腫瘤轉移過程,而GS的高表達促使M1表型巨噬細胞在白細胞介素(interleukin,IL)-4、IL-13和IL-10等作用下成為具有抗炎作用的M2表型[44]。GS增加細胞中琥珀酸水平,激活HIF-1使腫瘤細胞維持在M2表型,促進血管生成,增加T淋巴細胞的毒性作用,進而促進腫瘤轉移(圖2)。此外在腫瘤細胞中,癌基因c-myc被證明與谷氨酰胺成癮有關,在口腔癌中,c-myc作為上游調節因子通過上調GLS和GS活性促進EMT,抑制c-myc的表達后明顯減弱口腔癌細胞增殖和遷移能力[45]。
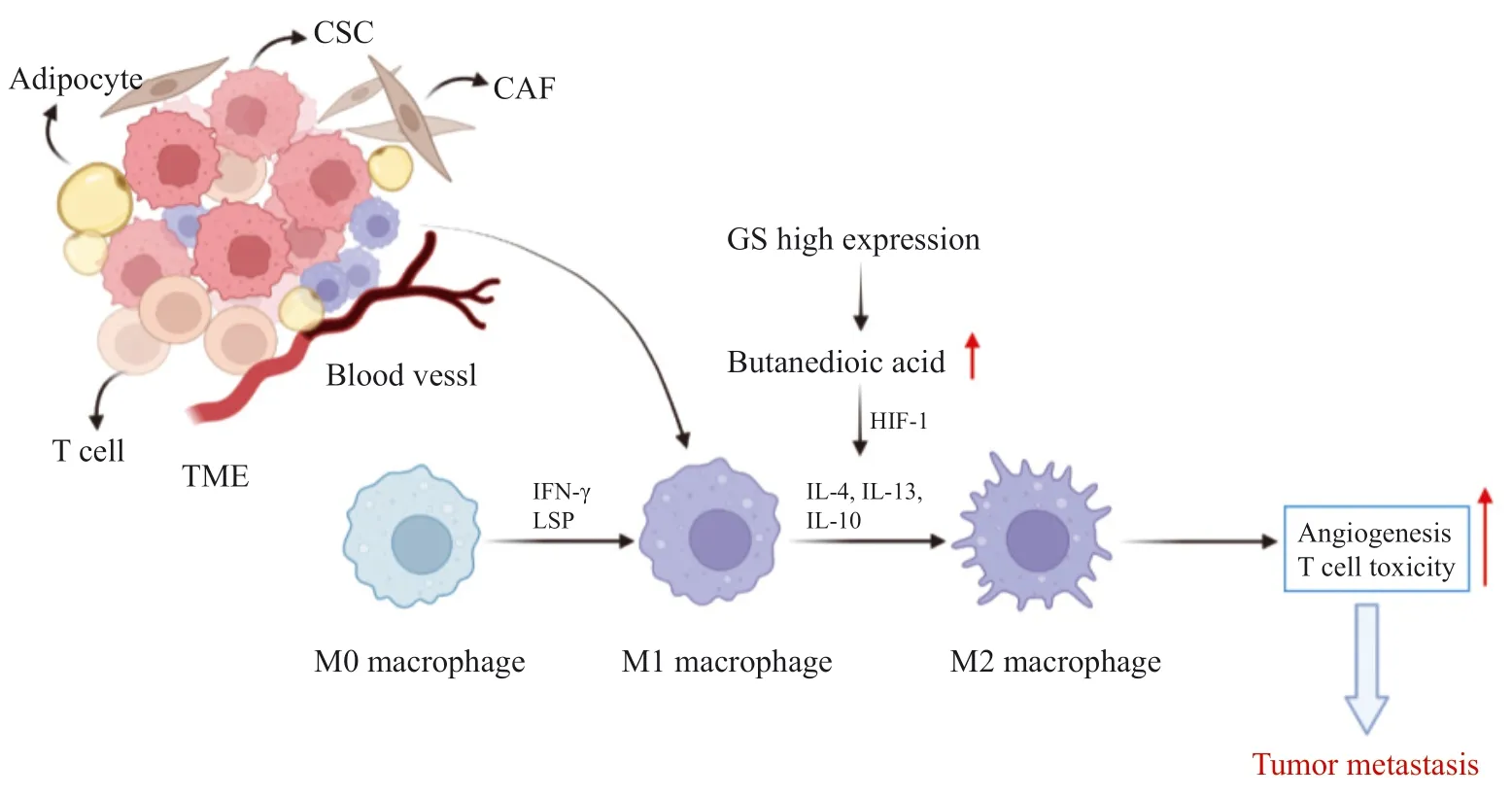
圖2 GS在TME的巨噬細胞活化中的作用Fig.2 The role of GS in macrophage activation in TME
2.3 GDH與腫瘤轉移
GDH用于催化谷氨酸生成α-KG,α-KG進入TCA參與能量供應和生物能合成。已有研究[46]表明,在腫瘤抑制因子LKB1缺失的肺癌細胞中,GDH1及下游產物α-KG通過CamKK2介導促進AMP活化蛋白激酶(AMP-activated protein kinase,AMPK)的磷酸化。AMPK被激活,觸發能量代謝,抑制下游mTOR信號通路,從而增強凋亡抵抗,促進肺癌細胞轉移[47]。GDH1通過酪氨酸135的磷酸化被表皮生長因子受體(epidermal growth factor receptor,EGFR)激活,通過CaMKIV信號轉導與RSK2一起增強cAMP響應性元件結合蛋白活性,從而促進肺癌轉移[48]。GDH還通過誘導STAT3介導的EMT來促進CRC細胞遷移。鑒于GDH的高表達與較差的臨床結果有關,GDH可作為CRC轉移的預后標志物之一[49]。有研究[50]顯示,具有腺苷二磷酸(adenosine diphosphate,ADP)-核糖基化活性的線粒體Sirtuin 4通過促進GDH1的糖基化來抑制前列腺癌進展中的谷氨酰胺代謝途徑。
2.4 TG與腫瘤轉移
TG能催化蛋白質或多肽鏈內谷氨酰胺殘基的水解和與其他氨基進行酰胺基的交換。TG蛋白家族包括TG1、TG2、TG3、TG4、TG5和TG6,其中TG2在參與并調控腫瘤轉移中發揮關鍵作用。TG2在多種癌癥中表達上調,如乳腺癌、CRC、肺癌、胰腺癌及卵巢癌等。
TG2同時具有酶促作用和非酶促作用,一方面TG2能催化Ca2+家族依賴蛋白的轉酰胺化,發揮轉氨酶的活性;另一方面,TG2具有GTPase、蛋白激酶和蛋白二硫化物異構酶活性[51],該活性在腫瘤細胞增殖、侵襲和轉移的信號轉導過程中發揮主要作用。此外,TG2在細胞外部和細胞內部都具有促進腫瘤轉移的作用[52]。TG2與細胞膜表面的整聯蛋白及細胞基質中的纖維連接蛋白相結合,誘導整聯蛋白介導的細胞黏附的產生,從而促進腫瘤細胞轉移,然而大部分TG2在細胞內部發揮作用,TG2可以與細胞內的HIF-1結合,進而刺激snail、Twist、ZEB等EMT調節因子的表達從而促進EMT,導致腫瘤細胞轉移能力增強。然而這些作用都依賴于TG2的GTPase活性,很少需要轉氨酶活性的參與。
在CRC中,TG直接調控EMT(snail和E-cadherin)和CSC(CD133和SOX2)相關蛋白的表達進而影響干細胞特性和腫瘤遷移及侵襲[53]。在肝癌中,癌癥相關成纖維細胞(cancer-associated fibroblast,CAF)通過TG2誘導IL-6-IL6R-STAT3軸促進EMT,加速肝癌細胞侵襲和轉移[54]。此外,也有研究[55]報道,真核延伸因子-2激酶通過上調TG2表達活性及整聯蛋白介導的通路來增強胰腺癌細胞的EMT和遷移。在胃癌中,TG2高表達,通過介導ERK1/2活性參與胃癌細胞增殖和遷移[56]。TG2也可作為口腔鱗狀細胞癌的生物標志物,在原發性腫瘤和轉移性病灶中TG2的mRNA水平明顯升高[57]。
2.5 谷氨酰胺轉運蛋白與腫瘤轉移
腫瘤細胞在轉移過程中,需要消耗大量的氨基酸進入細胞,而其中包括許多外源性的非必需氨基酸,為滿足外源性氨基酸進入腫瘤細胞的要求,氨基酸轉運體發揮著關鍵作用。谷氨酰胺進入細胞需要谷氨酰胺轉運蛋白的參與,如SLC1A5/ASCT2、SLC38A3等,其中SLC1A5是一種依賴Na+的跨膜轉運蛋白,被認為在多種腫瘤細胞中擔任主要的谷氨酰胺轉運體角色,在胃癌、CRC、乳腺癌、非小細胞肺癌(nonsmall cell lung cancer,NSCLC)和肝癌等癌癥中表達上調,能促進腫瘤細胞生長、增殖、遷移和EMT。在NSCLC中,SLC1A5可作為腫瘤淋巴結轉移的不良預后標志,其表達水平與在原位瘤及轉移性腫瘤中的表達呈正相關[58]。此外,在黏液表皮樣癌所分離出來的一種具有高轉移性的腫瘤組織MC3中,腫瘤抑制因子NDRG2通過抑制蛋白激酶B(protein kinase B,AKT)表達,減少對GSK3β的磷酸化,保持其活性,GSK3β通過誘導依賴Fbw7的c-myc的降解,導致c-myc活性降低,即從轉錄水平上抑制SCL1A5的表達,使腫瘤細胞依賴的谷氨酰胺分解減少,從而抑制EMT和腫瘤轉移[59]。SLC38A3同樣可通過激活PDK1/AKT信號通路減少EMT的上皮標志物E-cadherin的表達,增加間充質標志物N-cadherin的表達,從而促進EMT,導致NSCLC的細胞轉移[60]。與原發性腫瘤相比,SLC38A2在轉移性CRC組織中呈現高表達狀態,SLC38A2的缺失有利于抑制腫瘤進展[61],并且與臨床上三陰性乳腺癌患者的預后不良有關[62]。氨基酸逆向轉運蛋白SLC7A5也對晚期CRC中腫瘤的轉移和發展至關重要,在建立的轉移性實驗模型中,SLC7A5的缺失能顯著延長模型小鼠生存期且減少腫瘤轉移[63]。
3 靶向谷氨酰胺代謝干預腫瘤轉移的治療前景
目前處于臨床前研究的靶向GLS抑制劑包括CB-839、6-重氮-5-氧-L-正亮氨酸、968和BPTES等,但是由于腫瘤細胞內代謝的復雜性,不只有谷氨酰胺代謝的存在,各種信號轉導通路之間有重疊性,存在多個不同的靶點,因此谷氨酰胺代謝可以與其他臨床藥物聯合使用,通過對多個代謝通路的抑制,增加抑瘤作用。已有研究[64]表明,其他靶向藥物與GLS抑制劑聯用具有良好的抗腫瘤效果。在CRC的治療中,二甲雙胍與利巴韋林、奧沙利鉑和GLS抑制劑968聯用具有較好的療效[65]。968還可與自噬抑制劑氯喹聯用,氯喹抑制968引起的細胞自噬,增強968的靶向抗腫瘤作用[66]。因此,實際應用中,可將谷氨酰胺代謝抑制劑與其他靶向抗腫瘤藥物聯合應用,增加治療的可行性,為臨床研究提供理論和實驗基礎,有希望在未來開發更多的抗腫瘤藥物。
利益沖突聲明:所有作者均聲明不存在利益沖突。
作者貢獻聲明:
劉雪柔:前期文獻調研,文章初稿撰寫;
楊玉梅:文章框架整理;
趙倩:參考文獻收集和插入;
榮翔宇:文章校樣修改;
劉偉:文章返修內容核對,文獻整理;
鄭瑞潔:文章返修內容核對;
龐金龍:提供研究思路;
李嫻:提供基金支持;
李姍姍:提供基金支持,文章審閱和修改。
猜你喜歡
天津醫科大學學報(2019年3期)2019-08-13 06:53:08
中成藥(2016年8期)2016-05-17 06:08:14
癌癥進展(2016年12期)2016-03-20 13:16:17
罕少疾病雜志(2016年5期)2016-03-11 16:34:44
吉林大學學報(醫學版)(2015年1期)2015-12-17 07:47:28
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
中國當代醫藥(2015年16期)2015-03-01 02:03:11
中國醫藥導報(2015年26期)2015-02-28 22:07:59
肝膽胰外科雜志(2015年4期)2015-02-27 11:12:34
肝膽胰外科雜志(2015年4期)2015-02-27 11:12:24
