基于純培養和超高深宏基因組測序技術分析茅臺鎮兩企業高溫大曲微生物多樣性差異
2024-03-10 11:24:50王玉榮侯強川田龍新劉菊珍周加平
食品科學 2024年4期
關鍵詞:差異
王玉榮,侯強川,田龍新,劉菊珍,周加平,郭 壯,
(1.湖北文理學院 湖北省食品配料工程技術研究中心,湖北 襄陽 441053;2.襄陽市醬香型白酒固態發酵企校聯合創新中心,湖北 襄陽 441053;3.醬香型白酒固態發酵襄陽市重點實驗室,湖北 襄陽 441614)
白酒作為我國一款傳統的酒精飲品,不僅歷史悠久,還因其以酯類為主體的復合香味而深受各族人民的喜愛[1-2]。中國白酒最早確立的香型有清香型、醬香型、濃香型和鳳香型,隨著研究的深入,白酒香型已經擴展至12 種[3]。相較于其他香型,醬香型白酒具有醬香細膩、醇厚柔和、酒體豐滿和空杯留香持久等特點[4]。相關研究發現,白酒作為一種微生物發酵制品,酒曲不僅為其發酵過程提供了豐富的微生物和酶類物質[5],并直接或間接地影響著酒體的品質[6-7]。因此,在我國也有“曲為酒之骨”的說法。
根據生產原料差異大致可將酒曲分為麥曲、大曲和小曲等類型,而根據發酵工藝又可分為高溫曲、中高溫曲和中溫曲等。傳統酒曲的制作環境較為開放,容易受到制作環境和原料的影響,導致各曲樣間存在較大差異[8-9]。科研人員針對酒曲展開了大量卓有成效的研究,Hou Qiangchuan等[10]發現不同類型酒曲中微生物群落結構和功能之間存在明顯的差異;Wang Yurong等[11]進一步分析發現不同類型高溫大曲顯著影響著醬香型白酒的發酵過程,如黑色高溫大曲中主要以Thermoactinomyces和Staphylococcus為主,其不僅可以分泌淀粉酶、酯酶和纖維素酶等,還能產生具有醬香風味的吡嗪類物質和其他風味物質,對于醬香型白酒的風味品質具有重要的意義。同時,相關研究證實芽孢桿菌在白酒的發酵過程中發揮著重要的作用,其不僅具有多種酶,可以在發酵過程中分解一些復雜的底物,釋放營養物質促進酵母和其他微生物的生長和代謝,部分芽孢桿菌還會產生一些特殊風味的化合物,影響白酒的最終風味。茅臺鎮作為我國“醬酒圣地”,域內白酒業興盛,這也使得本研究能較好地解析該地區高溫曲中微生物的結構差異,并探究在發酵過程中的積極作用。
酒曲的研究是一個逐步發展的過程,科研人員最初采用純培養技術對酒曲中的細菌和真菌進行分離鑒定,初步探討了酒曲中微生物的構成和多樣性[12]。純培養技術具有自身的局限性,不能準確分析酒曲樣本中微生物的構成[13],但仍可以通過使用選擇性培養基實現對酒曲中目標微生物的分離和鑒定,為后續優良菌株的篩選奠定基礎。隨著高通量測序和生物信息學技術的迅速發展,高通量測序被廣泛應用于傳統發酵食品中[14-16]。值得注意的是,受限于擴增偏好和測序片段長度,該方法在解析物種構成和豐度上存在一定誤差,無法充分挖掘樣本中的微生物信息。近年來,隨著測序成本的快速下降,越來越多的科研人員采用宏基因組測序技術挖掘樣本中的微生物信息[17],而通過加大測序深度(超高深宏基因組測序)不僅可以獲得更加準確的物種鑒定和分類結果,還可以利用Binning技術獲得更加完整的基因組,并發現更多的低豐度物種,為解析生態中微生物的構成、多樣性和功能提供有利的手段。高溫大曲中的微生物結構較為復雜,因此,通過超高深宏基因組測序技術解析其微生物多樣性十分必要。
本研究以茅臺鎮兩個酒廠的6 份高溫大曲為研究對象,分別采用純培養技術和超高深宏基因組測序技術對高溫大曲中可培養微生物和微生物群落結構及功能進行分析,通過對比茅臺鎮不同酒廠高溫大曲中微生物群落結構和功能的差異,以期為提升高溫大曲的品質提供必要的數據支持。
1 材料與方法
1.1 材料與試劑
高溫大曲樣本均采集自貴州省遵義市仁懷市茅臺鎮,從A酒廠和B酒廠各采集3 份成品高溫大曲樣本,均生產自2022年6月21號,編號分別為A1~A3和B1~B3。兩酒廠高溫酒曲的制作工藝除曲母添加量存在差異外(A酒廠的曲母添加量為4.5%~8.0%,B酒廠的曲母添加量為6.0%~8.0%),其他工藝基本一致。
DNeasy mericon Food Kit DNA基因組提取試劑盒 德國QIAGEN公司;蛋白胨 北京陸橋技術股份有限公司;牛肉膏 南京全隆生物技術有限公司;氯化鈉、氫氧化鈉 國藥集團化學試劑有限公司;5×TransStartTMFastPfu緩沖液、FastPfu Fly DNA聚合酶、dNTPs Mix 北京全式金生物技術有限公司;引物27F/1495R 武漢天一輝遠生物科技有限公司。
1.2 儀器與設備
YH-2004電子天平 惠州市英衡電子科技有限公司;DG250型厭氧工作站 英國Don Whitley公司;CR21N型高速離心機 日本日立金屬株式會社;R930機架式服務器 美國Dell公司;Illumina Xten高通量測序平臺 美國Illumina公司;ND-2000C微量紫外分光光度計 美國Thermo Scientific公司;MGC-300H培養箱上海一恒科學儀器有限公司。
1.3 方法
1.3.1 樣品前處理
將收集的高溫大曲樣本破碎為粉,混合均勻后暫存于-80 ℃冰箱直至開展后續實驗。
1.3.2 芽孢桿菌的分離鑒定
使用無菌勺挖取10 g高溫大曲粉加入90 mL生理鹽水中振蕩均勻,使用倍比稀釋法將培養物稀釋至10-4~10-8梯度后涂布于營養瓊脂(nutrient agar,NA)培養基上,37 ℃倒置培養24 h[18]。選取菌落數在30~300的培養皿,并觀察平板中各菌落的形態,根據形態、顏色和大小等特征挑選不同的單菌落進行劃線純化。純化3 次后挑選恰當的菌體進行涂片和革蘭氏染色,使用十六烷基三甲基溴化銨法對疑似芽孢桿菌菌株的DNA進行提取,并使用引物27F(5’-AGAGTTTGATCCTGGCTCAG-3’)和1495R(5’-CTACGGCTACCTTCTTACGA-3’)對其16S rRNA全序列進行擴增。使用瓊脂糖凝膠電泳對擴增產物進行檢驗,并將檢驗合格的擴增產物連接到T載體后轉化至Escherichia colitop10中。最后,將陽性克隆子寄往測序公司進行測序,并將反饋回的序列拼接后在國家生物技術信息中心(National Center for Biotechnology Information,NCBI)數據庫中進行比對并構建系統發育樹[19]。
1.3.3 宏基因組DNA提取和測序
參照DNeasy mericon Food Kit DNA基因組提取試劑盒說明書的步驟對高溫大曲樣本進行宏基因組DNA提取,通過超聲將提取的宏基因組DNA打斷為350 bp左右,并將DNA末端進行補齊后添加A-tailed構建DNA文庫。最后,將構建好的DNA文庫使用Illumina Xten測序儀進行宏基因組測序。
1.3.4 生物信息學分析
使用KneadData軟件對下機數據進行質量控制,并去除人體基因組序列,將剩下的高質量宏基因組序列用于后續分析[20]。
本研究使用MegaHit軟件將高質量序列組裝成contigs,選擇長度大于1000 bp的contigs構建序列集,并基于Sembin軟件對構建的序列集進行宏基因組Binning以獲得宏基因組組裝基因組(metagenome assembly genomes,MAGs)[21];使用CheckM軟件對每一個MAG的質量進行評估,并依照完整度和污染率將MAGs劃分為高質量(完整度>80%,且污染率<10%)、中等質量(60%<完整度≤80%,且10%≤污染率<20%)和低質量(完整度≤60%,或污染率≥20%)[22];使用FastANI軟件將MAGs與NCBI非冗余核苷酸序列數據庫(Nucleotide Sequence Database,NT)和人體腸道微生物的基因組集(Unified Human Gastrointestinal Genome,UHGG)比對以獲得MAGs的注釋信息[23]。使用HUMAnN2(HMP Unified Metabolic Analysis Network 2)分析流程對質控后的宏基因組序列進行分析[24],并通過MetaphlAn 3軟件對高溫大曲中微生物的構成和相對豐度進行解析[25]。
1.3.5 核酸登錄號
本研究所用測序數據正在上傳國家生物技術信息中心SRA數據庫保存,該數據在NCBI數據庫的編號為PRJNA1008639。
1.4 數據分析及可視化
本研究基于R軟件對茅臺鎮不同酒廠高溫大曲中微生物的相對含量、α多樣性指數和微生物代謝通路的差異進行分析;使用PAST軟件對微生物的群落結構進行主坐標分析(principal co-ordinates analysis,PCoA)。本研究所有數據可視化均使用Origin 2021或R軟件完成。
2 結果與分析
2.1 高溫大曲中芽孢桿菌的分離鑒定
本研究從6 份茅臺鎮高溫大曲中共分離鑒定到212 株芽孢桿菌,其中從A酒廠高溫大曲中共分離鑒定到114 株芽孢桿菌(A1、A2和A3分別鑒定到41、38 株和35 株),而從B酒廠高溫大曲中共分離鑒定到98 株芽孢桿菌(B1、B2和B3分別鑒定到31、35 株和32 株),部分純化后的芽孢桿菌菌株在NA培養基中的菌落形態和細胞形態如圖1所示。
由圖1A可知,分離株的菌落形態多為圓形,表面粗糙、部分菌落中間凸起,顏色為透明或不透明的乳白色。由圖1B可知,菌株在電子顯微鏡下呈現短桿狀或桿狀,分離的菌株總體符合芽孢桿菌的典型特征。進一步分析發現,A酒廠分離的菌落形態多為圓形、中間凸起,呈不透明的乳白色,而B酒廠分離的菌落形態大多表面粗糙,呈透明的乳白色。由此可見,不同酒廠高溫大曲中分離的芽孢桿菌的形態存在較大差異。為進一步確定分離菌株的分類學地位,本研究對菌株的16S rRNA基因全長進行測定,并在NCBI網站進行BLAST比對,同時將所獲得的序列與模式菌株序列通過MEGA軟件構建系統發育樹,系統發育樹結果如圖2所示。
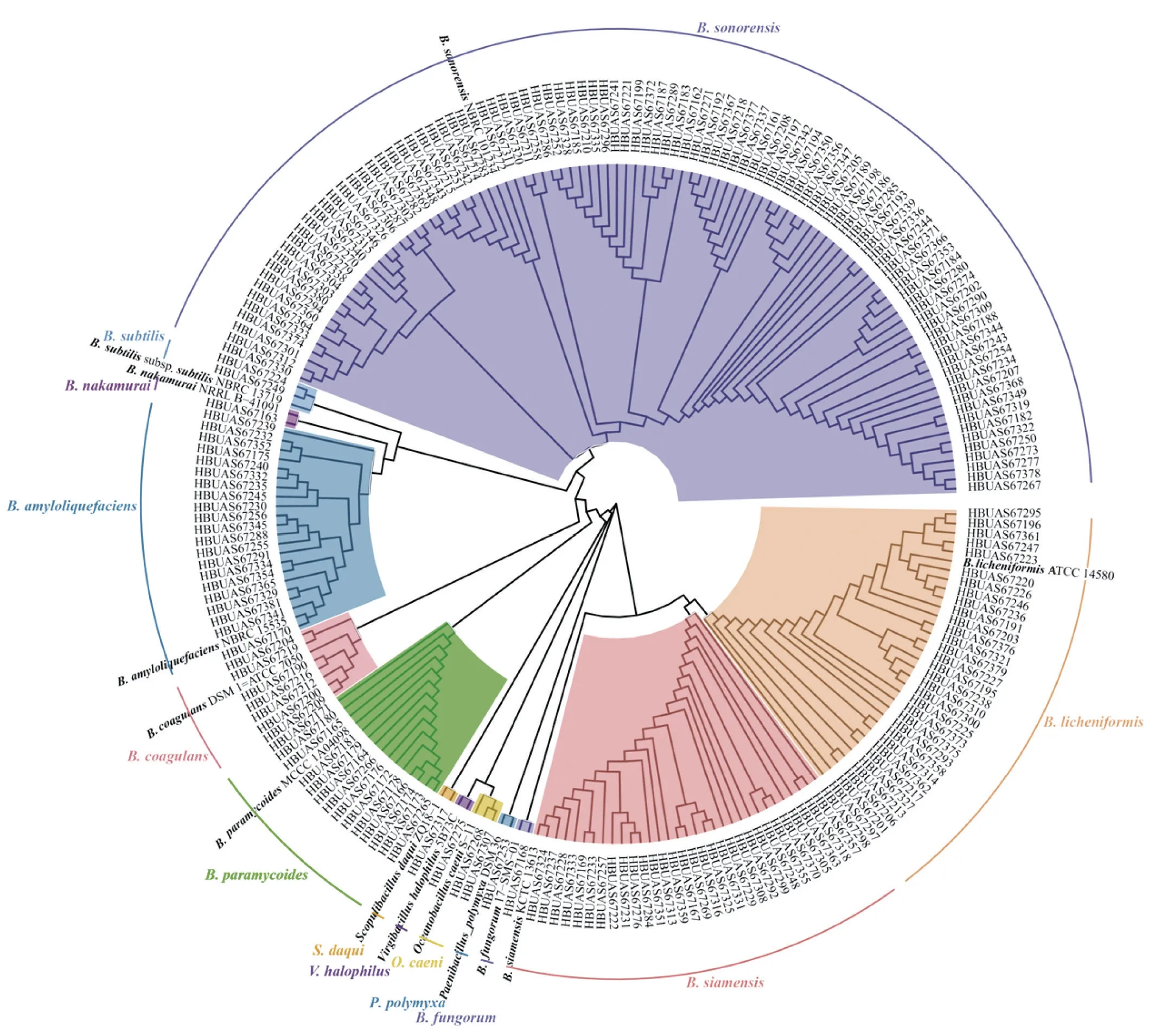
圖2 基于16S rRNA的212 株芽孢桿菌與標準菌株的系統發育樹Fig.2 Phylogenetic tree of 212 Bacillus stains and standard strains based on 16S rRNA sequencing
本研究分離出的212 株芽孢桿菌被鑒定為20 個種,分別為77 株索諾拉沙漠芽孢桿菌(Bacillus sonorensis)、39 株地衣芽孢桿菌(B.licheniformis)、19 株解淀粉芽孢桿菌(B.amyloliquefaciens)、16 株暹羅芽孢桿菌(B.siamensis)、15 株地衣芽孢桿菌(B.licheniformis)、13 株擬蕈狀芽孢桿菌(B.paramycoides)和11 株貝萊斯芽孢桿菌(B.velezensis)等。進一步分析可知,B.sonorensis主要由B酒廠中分離得到,B.licheniformis和B.amyloliquefaciens主要由A酒廠中分離得到,而其他菌株在兩酒廠中分布差異不明顯。綜上所述,茅臺鎮高溫大曲中的芽孢桿菌以B.sonorensis、B.licheniformis和B.amyloliquefaciens等為主,占比63.68%。相關研究顯示,B.sonorensis、B.licheniformis和B.amyloliquefaciens等為高溫大曲中的主要芽孢桿菌,不僅參與醬香型白酒的發酵過程,還可以控制有害微生物的生長,產生一些具有特殊風味的化合物,并可能影響白酒風味。由此可見,通過對高溫大曲的芽孢桿菌進行分離鑒定和保藏,可為后續優良菌種的開發提供必要的原材料。
2.2 高溫大曲中微生物構成和多樣性分析
本研究在純培養的基礎上,通過超高深宏基因組測序對高溫大曲微生物類群進行解析,6 個樣品共獲得867530532 條高質量序列,測序總量為124.08 Gb,每個樣本平均獲得144588422 條高質量序列,平均測序量為20.68 Gb。在獲得宏基因組數據的基礎上,本研究采用Binning技術對茅臺鎮高溫大曲中微生物的基因組進行組裝,結果如圖3所示。
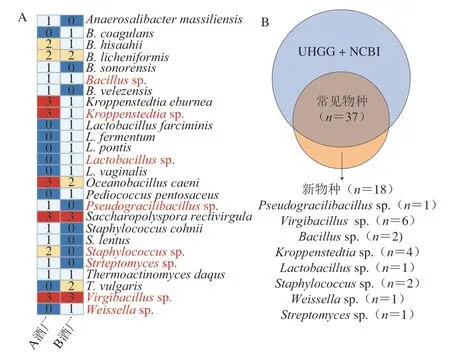
圖3 基于宏基因組Binning技術的物種構成分析Fig.3 Species composition analysis based on metagenome binning technology
本研究共獲得303 個MAGs,其中高質量的MAGs有55 個。由圖3A可知,本研究獲得的55 個高質量MAGs分別隸屬于13 個屬的26 個種。進一步分析可知,Virgibacillussp.和Saccharopolyspora rectivirgula在所有酒曲樣本中均能被鑒定到,而Oceanobacillus caeni、Kroppenstedtiasp.和K.eburnea僅在A酒廠的所有大曲中被鑒定到。值得注意的是,Staphylococcus和Streptomyces僅在A酒廠的大曲中被鑒定到,而Lactobacillus僅在B酒廠的大曲中被鑒定到。
將高質量MAGs與數據庫中基因組的平均核苷酸相似度(average nucleotide identity,ANI)進行計算發現,55 個高質量的MAGs中有37 個與數據庫中的基因組具有較高水平的ANI(≥97%),而剩余18 個與數據庫中基因組的ANI較低(<97%)。由此可見,37 個高質量MAGs可以在種水平被注釋,而其他18 個為疑似新種(Pseudogracilibacillussp.、Virgibacillussp.、Bacillussp.、Kroppenstedtiasp.、Lactobacillussp.、Staphylococcussp.、Weissellasp.和Streptomycessp.)。在此基礎上,本研究進一步對茅臺鎮高溫大曲中微生物的物種構成和多樣性進行解析,結果如圖4所示。
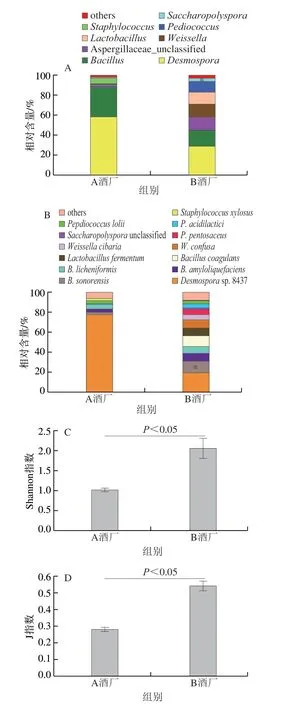
圖4 茅臺鎮高溫大曲中微生物的組成和多樣性Fig.4 Composition and diversity of microorganisms in high temperature Daqu from Maotai town
本研究一共注釋到6 個菌門和109 個菌種。由圖4A可知,高溫大曲中的優勢(平均相對含量大于1.00%)菌屬有8 個,分別為43.21%芽胞鏈菌屬(Desmospora)、22.99%芽孢桿菌屬(Bacillus)、7.07%曲霉科_未識別(Aspergillaceae_unclassified)等。由圖4B可知,優勢菌種有13 個,主要為48.46%Desmosporasp.8437、6.93%B.sonorensis和5.65%B.amyloliquefaciens等。值得注意的是,A酒廠高溫大曲中子囊菌門(Ascomycota)的相對含量高達12.74%,其真菌相對含量較高。本研究進一步對不同酒廠高溫酒曲中優勢菌屬和菌種的相對豐度進行分析,發現Desmospora和Bacillus是高溫酒曲中的主要菌屬,而Desmospora_sp_8437是高溫酒曲中的主要菌種。差異分析發現,在優勢菌屬方面,僅Staphylococcus在A酒廠高溫大曲中顯著較高(P<0.05),而其他優勢菌屬均差異不顯著(P>0.05);在優勢菌種方面,A酒廠高溫大曲在B.sonorensis、B.amyloliquefaciens和B.coagulans等芽孢桿菌的相對豐度上顯著較低(P<0.05)。由圖4C、D可知,A酒廠高溫大曲中微生物的Shannon指數和J指數分別為1.01和0.28,而B酒廠高溫大曲分別為2.06和0.54。經Wilcoxon檢驗發現,A酒廠高溫大曲中微生物的Shannon指數和J指數均顯著低于B酒廠高溫大曲(P<0.05)。由此可見,其微生物的多樣性和豐富度均顯著低于B酒廠高溫大曲(P<0.05)。基于Bray-Curtis和Jaccard距離對茅臺鎮不同高溫大曲的菌群結構進行分析,結果如圖5所示。
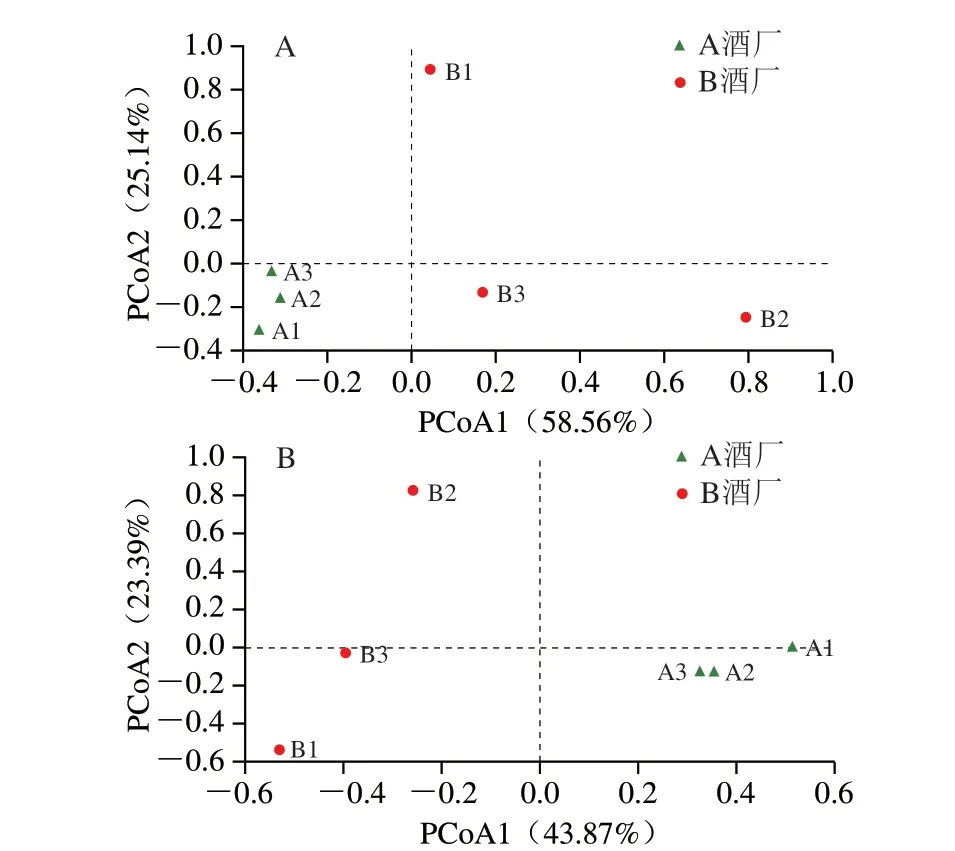
圖5 基于Bray-Curtis(A)和Jaccard(B)距離的PCoA圖Fig.5 PCoA plots based on Bray-Curtis (A) or Jaccard (B) distance
由圖5A可知,在基于Bray-Curtis距離的PCoA圖中,A酒廠高溫大曲主要集中在第3象限,而B酒廠高溫大曲則較為分散,不同高溫大曲在空間上呈現明顯的分離趨勢(F=1.256,P=0.204);由圖5B可知,在基于Jaccard距離的PCoA圖中,A酒廠高溫大曲主要集中在X軸正方向,而B酒廠高溫大曲則主要集中在X軸負方向,兩種高溫大曲亦在空間上呈現明顯的分離趨勢(F=3.453,P=0.024)。由此可見,高溫大曲中的主要微生物存在較大差異,可能各自含有一些較為獨特的細菌類群。值得注意的是,A酒廠高溫大曲在空間上較為集中,而B酒廠高溫大曲在空間上較為分散,表明A酒廠高溫大曲中的微生物結構更為相似穩定。
2.3 高溫大曲中微生物的代謝途徑
本研究基于Humann2分析流程對高溫大曲中微生物的功能進行預測,并通過MetaCyc數據庫對微生物的代謝途徑進行注釋。茅臺鎮不同高溫大曲中微生物差異代謝途徑如圖6所示。
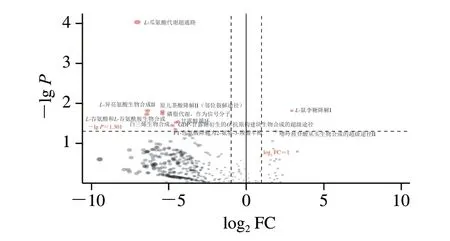
圖6 茅臺鎮不同高溫大曲中微生物差異代謝途徑分析Fig.6 Analysis of microbial differential metabolic pathways between different high-temperature Daqu from Maotai town
由圖6可知,從茅臺鎮高溫大曲中共注釋到367 條代謝途徑,而有11 條代謝途徑在不同高溫大曲中存在顯著性差異(P<0.05),其中9 條代謝途徑的相對豐度在A酒廠高溫大曲中顯著較高(P<0.05),2 條代謝途徑的豐度在B酒廠高溫大曲中顯著較高(P<0.05)。進一步分析發現,在A酒廠高溫大曲中豐度較高的差異代謝通路主要與氨基酸的生物合成相關,而在B酒廠高溫大曲中豐度較高的差異代謝通路主要與L-鼠李糖的降解相關。為此,本研究進一步對在差異代謝途徑中各菌的貢獻度進行分析,具體結果如圖7所示。
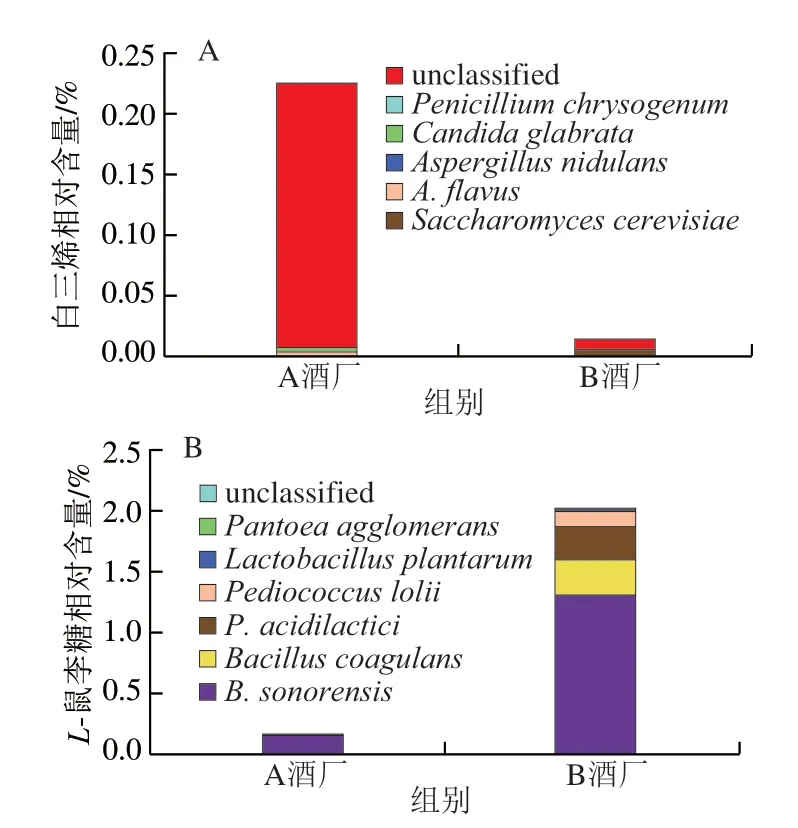
圖7 部分差異代謝途徑中微生物的貢獻度分析Fig.7 Analysis of the contribution of microorganisms to selected differential metabolic pathways
本研究對11 條差異代謝途徑的物種貢獻進行分析,發現有9 條差異代謝途徑的物種貢獻為未知,僅有2 條差異代謝途徑的物種貢獻較為明確。由圖7可知,白三烯的生物合成主要由釀酒酵母(Saccharomyces cerevisiae)、黃曲霉(Aspergillus flavus)和構巢曲霉(A.nidulans)等貢獻,而L-鼠李糖降解則主要由B.sonorensis、B.coagulans和乳酸片球菌(Pediococcus acidilactici)等貢獻。值得注意的是白三烯生物合成的差異主要是由A酒廠高溫大曲中的未知物種所貢獻。
3 討論與結論
酒曲作為白酒發酵的靈魂,在白酒的生產發酵中扮演著重要的角色。作為白酒生產過程中重要的糖化劑和發酵劑,其不僅提供發酵過程中所需要的各種微生物,還提供各種酶類及風味物質的前體,直接或間接影響著酒體最終品質的形成[26]。在高溫大曲制作時,大量環境或原材料中的微生物進入發酵體系進行富集,并經過一段時間高溫發酵,最終形成其所特有的微生物菌群和多樣性[27]。有研究發現高溫發酵會導致大曲中微生物的構成發生明顯變化,使得如Bacillus、嗜熱真菌屬(Thermomyces)和熱子囊菌屬(Thermoascus)等對高溫耐受性較強的微生物占據優勢地位,而酵母等對高溫耐受性相對較差的微生物豐度顯著降低[28]。通常酵母的存在能有效提高白酒中酮類、酯類和酚類等揮發性化合物的濃度,從而提升白酒的感官品質。本研究通過純培養技術從茅臺鎮高溫大曲中分離得到212 株芽孢桿菌,其中大部分為B.sonorensis、B.licheniformis和B.amyloliquefaciens等。侯強川等[29]對茅臺和堯治河高溫大曲中細菌群落結構進行解析時亦發現Bacillus是高溫大曲主要菌屬,且不同地區和類型的高溫大曲中Bacillus的相對含量存在明顯差異。Wu Xue等[30]也證實了Bacillus對醬香型白酒中的風味物質具有較大的影響,在醬香型白酒發酵過程中具有廣泛的應用。由此可見,通過對高溫大曲中的芽孢桿菌進行分離、鑒定和保藏,可為后期菌株的開發利用做準備。
通過Binning技術共獲得55 個高質量MAGs,其中有18 個疑似新菌株,主要為Bacillussp.、Lactobacillussp.和Weissellasp.等。本團隊在前期研究中通過超高深宏基因測序技術發現房縣小曲中存在一株疑似的新菌株Weissellasp.[31],并在后期實驗中進行了驗證,同時證明其在房縣黃酒的發酵過程中扮演著重要的角色[32]。由此可見,通過超高深宏基因組測序結合Binning技術可以為新菌株的鑒定提供可靠技術支持。相關研究證實,真菌在醬香型白酒的發酵過程中起著至關重要的作用,對于其發酵進程和風味形成均有著積極影響[33]。值得注意的是,A酒廠高溫大曲中芽孢桿菌的相對含量相較于B酒廠高溫大曲明顯較低,而這可能與酒曲的制作和發酵環境,以及微生物的相互作用有著緊密的聯系[33]。對高溫大曲中微生物的多樣性進行分析發現,A酒廠高溫大曲中微生物的多樣性和豐富度均顯著低于B酒廠高溫大曲,兩者微生物的菌群結構存在明顯的差異。微生物的多樣性與發酵過程中酵母的代謝活性有著密切聯系,多樣的微生物群落可以在發酵中利用不同底物,從而加速發酵的進行。侯強川等[29]研究發現不同地區采集的大曲中細菌的群落結果存在明顯差異。由此可見,除地區因素外,酒曲的制作工藝也會造成高溫大曲中微生物群落結構的差異。而值得注意的是,A酒廠高溫大曲中微生物的群落結構較為相似,B酒廠高溫大曲中微生物群落結構存在較大的差異,而微生物群落的不同組成可以影響酒的感官特性,使其呈現出個性化風味。
在功能方面,兩種高溫大曲存在多條差異代謝通路,其中在A酒廠高溫大曲中顯著較高的差異代謝通路大多與氨基酸的代謝相關,而B酒廠高溫大曲中的差異代謝通路主要與L-鼠李糖的降解相關。相關研究發現L-谷氨酸和L-異亮氨酸降解加快不僅可以加速發酵的進程,促進酒精的產生,還可能在分解過程中產生香氣化合物(如酯類、醛類和酮類等)和一些揮發性酸(如乙酸和丁酸等),從而賦予白酒特殊的芳香特征和口感[34-35]。L-鼠李糖的降解大多是由酵母菌完成,而更高的L-鼠李糖降解速率不僅可以加快發酵速率,產生更多的酒精和香氣化合物,還可能產生一些代謝產物,提升酒體的穩定性[36]。由此可見,不同酒廠因其獨特的曲種,可能塑造不同的酒體香味特征和口感。對部分差異代謝通路的進一步分析發現,同一代謝通路由多種微生物貢獻。不同菌群的最適生長條件存在差異,且菌群在不同環境中生長狀態也不相同,這也使得某些代謝途徑容易受到環境影響,從而影響白酒的發酵進程。因此,當同一代謝通路由多種微生物貢獻,不僅可以提升代謝效率,還可以提升其環境適應性,使得代謝通路表達能維持相對穩定,從而在一定程度上維持酒曲品質的穩定。綜上所述,后續研究應進一步擴大酒曲的采樣范圍和數量,并結合全基因組測序技術進一步解析微生物在醬香型白酒發酵過程中的積極作用,為后期優良菌株的篩選和發酵工藝的改良提供必要數據支持。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
