
多抗霉素對煙草靶斑病的防效及應用后其葉際微生物的變化規律
2024-04-08 09:59:48郭沫言汪漢成張藝蔡劉體王清李文紅史彩華
江蘇農業科學 2024年3期
關鍵詞:群落結構
郭沫言 汪漢成 張藝 蔡劉體 王清 李文紅 史彩華
摘要:為了評價多抗霉素對煙草靶斑病的田間防效及藥劑施用后不同時期葉際微生物的變化規律,通過田間小區施用多抗霉素,并于施藥后不同時期調查田間煙草靶斑病發病情況計算病情指數,于不同時期取樣并通過Illumina HiSeq高通量測序技術分析多抗霉素對健康與感病煙葉葉際微生物群落結構及多樣性。結果表明,多抗霉素處理3、9 d 的防效分別為89.80%、84.64%。施藥后,感病組織亡革菌屬的相對豐度顯著降低,Symmetrospora的相對豐度呈提高趨勢;健康與感病組織細菌中假單胞菌屬、甲基桿菌屬、鞘氨醇單胞菌屬、金剛藻屬的相對豐度均提高;施藥后 18 d,健康與感病組織葉際真菌和細菌多樣性及豐富度均提高。葉際微生物網絡關系互作圖顯示,施藥后亡革菌屬與鏈格孢屬呈負相關,且與其他真菌屬大多也呈負相關;細菌屬之間互作關系大多呈正相關,部分細菌屬之間呈負相關。綜上,多抗霉素能快速且穩定地防治煙草靶斑病的發生,能有效降低煙草靶斑病病原菌的相對豐度,同時對葉際其余菌屬存在影響,隨著時間的延長,防效較為穩定,證明多抗霉素能廣泛用于煙草靶斑病的防治。
關鍵詞:煙草靶斑病;多抗霉素;田間防效;葉際微生物;群落結構
中圖分類號:S435.72? 文獻標志碼:A
文章編號:1002-1302(2024)03-0138-09
煙草靶斑病(tobacco target spot)是由立枯絲核菌(Rhizoctonia solani)引起的真菌性病害,其有性世代為瓜亡革菌(Thanatephorus cucumeris)[1]。該病害可侵染煙草根部、莖基部和葉部,但以危害煙草葉片為主,病害流行時造成煙草大幅減產,經濟受損嚴重。該病害最早于1948年在巴西發現,2006年在我國遼寧發現并報道,隨后陸續在我國各煙草主產區暴發[2-3]。煙草靶斑病病菌在溫度適中、相對濕度較高和葉片長時間濕潤的條件下可大量繁殖,病害發生嚴重時煙葉壞死穿孔,導致煙葉無法正常采收,造成巨大的經濟損失[4]。
化學藥劑防控是防治煙草靶斑病的主要手段。目前,已登記的防控煙草靶斑病的藥劑主要為8%井崗霉素水劑[5],已經開展過相關研究的藥劑有嘧菌酯、嘧肽菌凈、噻氟酰胺和多抗霉素[6-9]。其中,多抗霉素是一種用現代生物工程技術生產的肽嘧啶核苷類農用抗生素,主要作用于病菌細胞壁幾丁質的生物合成,導致芽管和菌絲局部膨大、破裂、內含物溢出,病原菌不能正常發育乃至死亡[10-12],同時多抗霉素還能有效抑制病原菌產孢,具有內吸性、治療作用及高度的靶標生物選擇性,屬于廣譜性抗生素類殺菌劑[13]。多抗霉素常用來防治西瓜炭疽病、白菜霜霉病、番茄灰霉病和黃瓜白粉病等蔬菜水果以及農作物上的病害,同時也能有效防治煙草赤星病[14-19],但對于同樣在煙草上發生嚴重的煙草靶斑病防控效果研究較少。
植物葉際微生物之間關聯緊密,研究其與病害的關系是防治植物病害的重要途徑[20-21]。煙草病害發生在不同時期,其葉際微生物的多樣性和豐富度都會發生顯著的變化。研究表明,煙草赤星病發生在不同部位煙葉的微生物結構存在差異,時空分布格局中下部葉片葉際微生物的多樣性和豐富度較低,上部葉片較高[22-23]。而隨煙草靶斑病發病程度增加,健康煙葉與感病煙葉的葉際真菌物種多樣性顯著降低,葉際細菌的物種多樣性上升[24]。病害的發生與其葉際微生物的變化趨勢緊密相關,藥劑防控通過對病原菌的作用達到防治效果,但廣譜性殺菌劑作用靶標為葉際多種致病菌[25]。田間病害的發生通常為多種病原菌共同作用的結果,在煙草靶斑病發生期,其葉際優勢真菌有亡革菌屬、Stagonosporopsis、鏈格孢屬、亞隔孢殼屬真菌,葉際優勢細菌有假單胞菌屬、泛菌屬、腸桿菌屬細菌;Didymella segeticola引起的煙草病害葉際優勢真菌屬含有鏈格孢屬和Boeremia等致病菌;感染煙草野火病的煙葉葉際優勢細菌屬含有假單胞菌屬、鞘氨醇單胞菌屬和泛菌屬等有益菌屬;感染煙草赤星病的煙葉葉際同時存在鏈格孢屬、莖點霉屬和鐮刀菌屬等致病菌以及泛菌屬、假單胞菌屬和鞘氨醇單胞菌屬等有益細菌[26-30]。
研究藥劑對葉際微生物的影響有利于揭示植物病害的發生機制以及藥劑的防控效果。因此,本研究通過田間施用多抗霉素,調查其對煙草靶斑病的田間防效,并對田間采集樣品的葉際微生物進行分析,探索總結在施藥后不同時期葉際優勢菌屬的物種組成以及豐富度的變化趨勢,為煙草靶斑病的藥劑防控提供參考依據。
1 材料與方法
1.1 供試藥劑
10%多抗霉素可濕性粉劑(WP),青島瀚生生物科技股份有限公司生產。
1.2 試驗方法
本試驗于2020年8月中旬在貴州省畢節市黔西縣高坡開展,試驗品種為云煙87。隨機選取煙株長勢一致的煙田并劃分小區,每個小區20株煙株,3次重復。于煙葉底部葉片零星出現病斑時開始用藥,參照藥劑說明,多抗霉素用量為1.5 kg/hm2,兌水量為900 L/hm2,采用多功能噴霧施肥器噴灑藥劑,使其均勻噴施于葉片表面。不同小區之間設立保護行,避免相互干擾。在施藥前(0 d)隨機選擇相同部位、相同成熟度的葉片,隨機選取并標記8個病斑,于施藥后0、1、3、9 d,采用“十”字交叉法對標記病斑進行測量,計算多抗霉素處理對煙草靶斑病病斑擴展的抑制率[31]。各小區隨機選擇8株煙株,分別于施藥后0、1、3、9 d調查統計其病情指數,計算多抗霉素對煙草靶斑病的田間防效;參照煙草靶斑病病害分級標準[32]調查病害等級。
病情指數=∑(各級病葉數×相對級數值)/(調查總葉數×最高病級值)×100;
防效=[1-(處理區病情指數增長值/對照區病情指數增長值)]×100%;
病斑增長率=[第n次測量病斑直徑/第(n-1)次測量病斑直徑-1]×100%。
1.3 樣品采集與處理
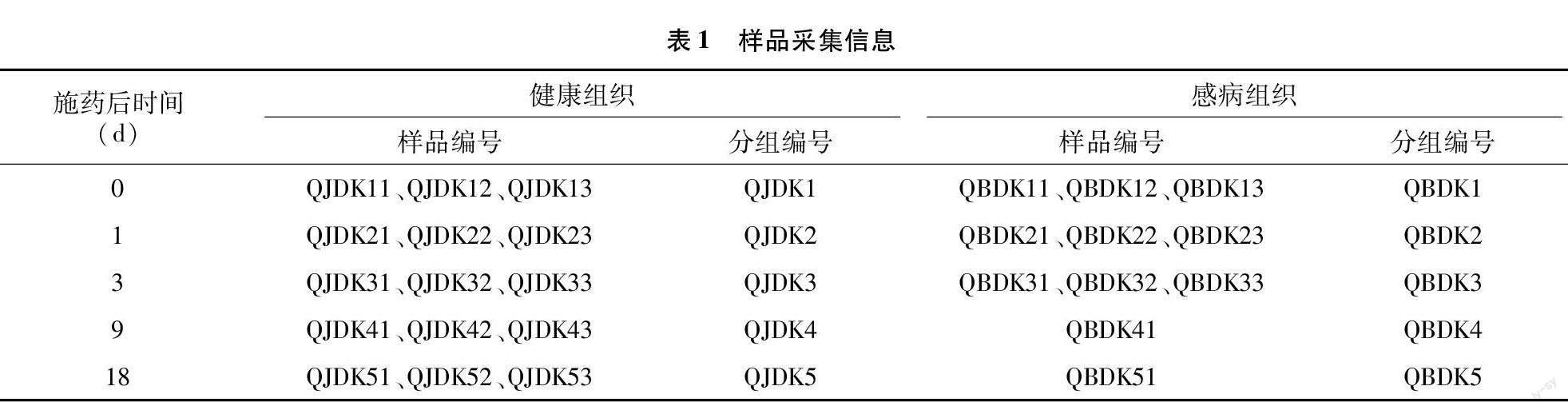
于施藥后0、1、3、9、18 d隨機選取施藥煙苗健康與感病部位,用無菌剪刀對其進行剪取采集,分別裝入50 mL無菌離心管中,每組3次重復,采集完成后迅速進行試驗分析,剩余樣品存放于-80 ℃冰箱中。樣品采集信息詳見表1,施藥后9、18 d 由于病害晚期可供采集的樣品有限,因此選取的樣品為混合樣品。
采用CTAB法分別提取各樣本基因組DNA,使用無菌水稀釋樣本至1 ng/μL,使用特異性引物進行擴增,使用2%瓊脂糖凝膠電泳檢測PCR產物,使用德國QIAGEN公司提供的膠回收試劑盒對目的條帶進行回收,使用 TruSeq DNA PCR-Free Sample Preparation Kit建庫試劑盒進行文庫構建,文庫合格后使用NovaSeq 6000進行上機測序。
根據Barcode序列和PCR擴增引物序列拆分出各樣本數據,過濾低質量和短長度后的序列,截去特異性引物序列后對每個樣本進行拼接,得到原始數據。原始序列通過USEARCH擴增子分析軟件與物種注釋數據庫進行比對檢測嵌合體序列,并最終去除其中的嵌合體序列[33],得到最終的有效數據(effective tags)。
將所有序列聚類成為操作單元(OTU)進行物種注釋,使用Qiime軟件version 1.9.1分析其樣本復雜度,主要包括α多樣性(α diversity)、菌群豐度(community richness)、菌群多樣性(community diversity)、測序深度(sequencing depth)和系統發育多樣性(phylogenetic diversity)、環境關聯因子、網絡互作以及功能注釋以上分析均在北京諾和致源科技股份有限公司完成。
1.4 數據分析
使用Excel 2022對調查數據進行統計分析處理,計算多抗霉素對煙草靶斑病的田間防效。運用DPS v9.01數據處理系統對測序樣品的α多樣性指數和菌群豐度進行差異顯著性分析。采用Adobe Photoshop CS5對相關圖片進行處理。
2 結果與分析
2.1 多抗霉素對煙草靶斑病的田間防效
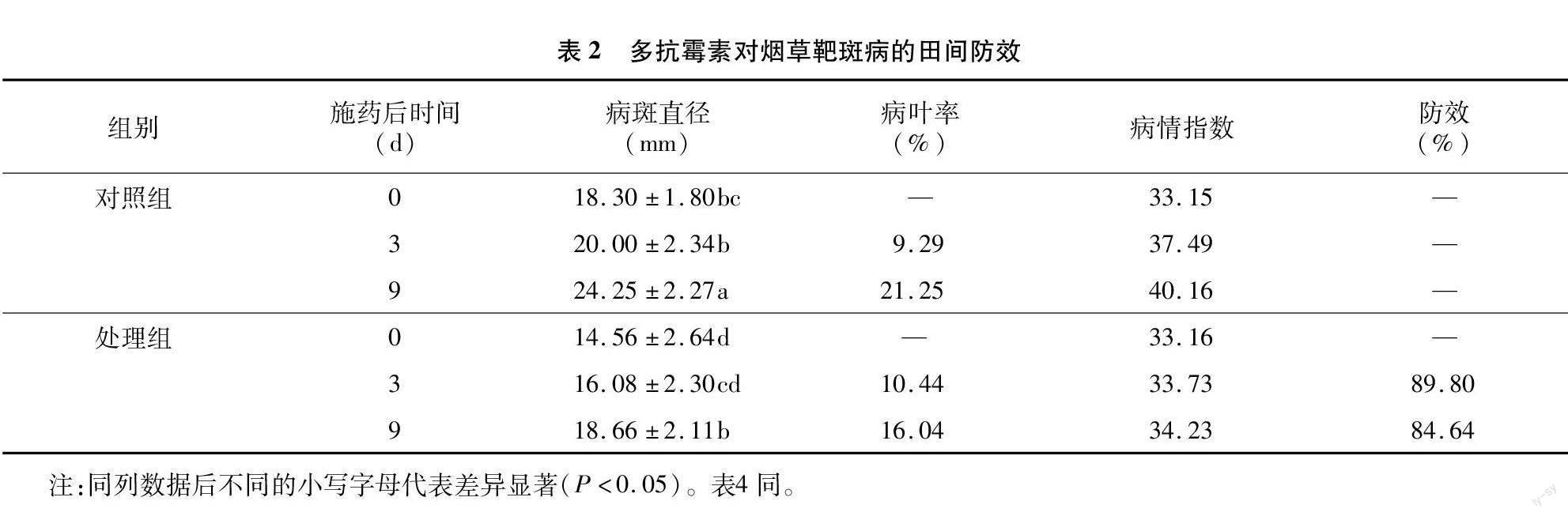
由表2可知,多抗霉素對煙草靶斑病具有良好的防效。施藥后,煙草病斑直徑增長率降低,病情指數增長變緩,煙草靶斑病的發生得到顯著控制。施藥后 3 d,對照組的病情指數達到37.49,處理組病情指數達到33.73,多抗霉素對煙草靶斑病田間防效達到89.80%;施藥后9 d,對照組的病情指數為40.16,處理組病情指數為34.23,多抗霉素對煙草靶斑病田間防效達到84.64%。
2.2 多抗霉素防治煙草靶斑病不同時期煙葉葉際微生物組成
2.2.1 健康與感病組織葉際微生物測序深度
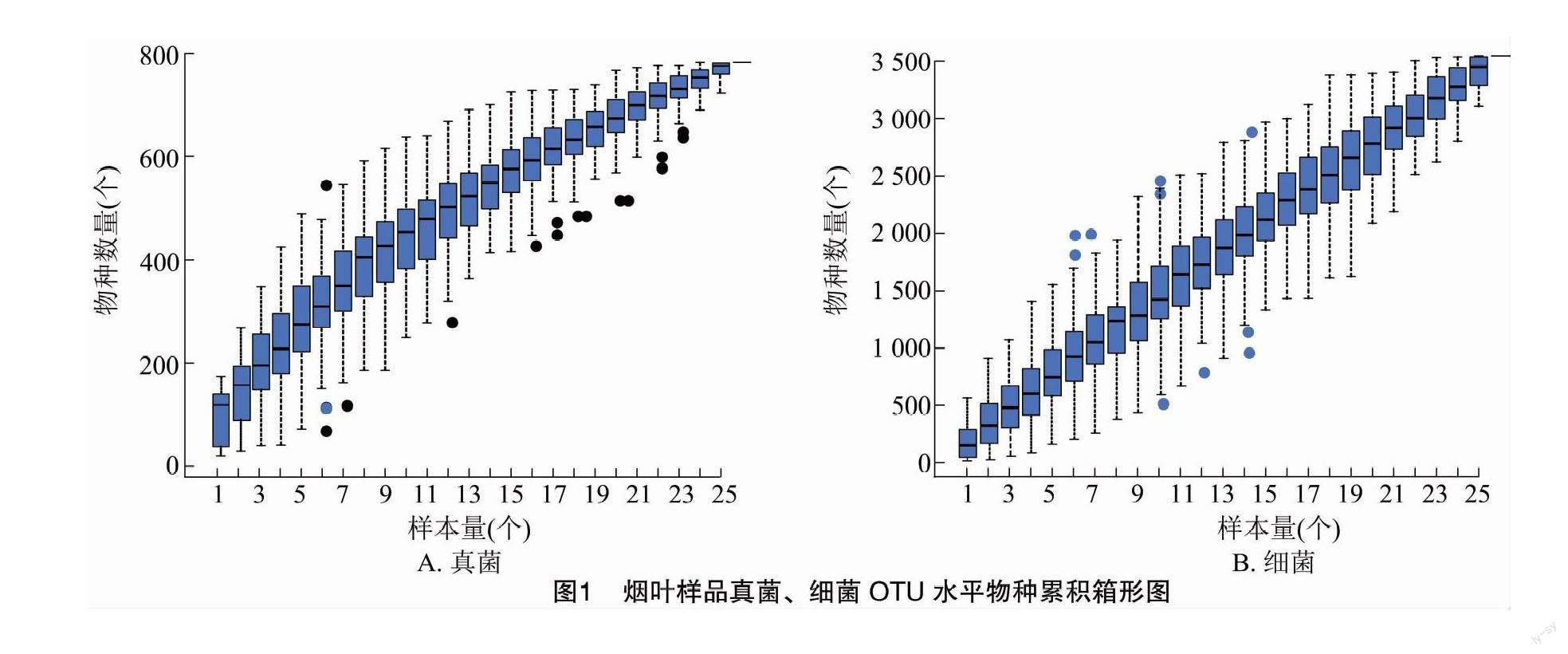
從樣本中隨機抽取一定測序量的數據,統計每組OTU數量,以樣本量與OTU總數構建曲線,可直接反映測序數據量的合理性,間接反映樣本中物種的豐富程度。當曲線趨向平坦時,說明測序樣本量漸進合理,可以進行下一步分析。本次測序共25個樣本量,越接近25個時,真菌的物種數量越接近800個,細菌的物種數量越接近3 500個,曲線逐漸趨于平緩,表明測序深度已經足夠,可以進行數據分析(圖1)。
2.2.2 健康與感病組織葉際微生物OTU結構
如圖2所示,在葉際真菌群落中,健康組織中真菌共有OTU為28個,感病組織真菌共有OTU為10個,感病組織OTU數量低于健康組織OTU數量。多抗霉素處理后1、3、9、18 d,健康組織與感病組織OTU數量呈現“先增加后減少又增加”的趨勢。藥劑處理后18 d,健康組織OTU數量達到峰值,為159個;藥劑處理3 d后,感病組織OTU數量達到峰值,為74個。細菌群落中,健康組織細菌共有OTU為26個,感病組織細菌共有OTU為24個,健康組織OTU數量均高于感病組織OTU數量。多抗霉素處理1、3、9、18 d后,健康與感病組織OTU數量呈現“先增加后減少又增加”的趨勢。健康組織OTU數量在施藥后18 d達到峰值,為1 106個,在施藥前OTU數量最少,為27個;感病組織OTU數量在施藥后1 d達到峰值,為348個,在施藥前OTU數量最少,為46個。
2.2.3 健康與感病組織葉際微生物多樣性
多抗霉素處理后不同時間,煙葉葉際真菌與細菌的相對豐度均存在明顯變化(表3、圖3)。在真菌門水平,其葉際優勢菌門為擔子菌門(Basidiomycota)、子囊菌門(Ascomycota)和毛霉門(Mucoromycota)。藥劑處理后,健康組織與感病組織擔子菌門的相對豐度均提高,子囊菌門的相對豐度降低。在真菌屬水平,優勢菌屬包括亡革菌屬(Thanatephorus)、Symmetrospora、毛霉屬(Mucor)、小不整球殼屬(Plectosphaerella)、尾孢屬(Cercospora)、漢納酵母屬(Hannaella)、鏈格孢屬(Alternaria)、紅酵母屬(Rhodotorula)、枝孢屬(Cladosporium)和頂孢霉屬(Acremonium)。隨著施藥時間的延長,健康組織各優勢菌屬的數量增加。施藥后1~3 d,健康組織與感病組織亡革菌屬的相對豐度持續下降;施藥后 18 d,健康組織中其相對豐度降低為0.24,感病組織中其相對豐度低于健康組織。施藥后,健康組織與感病組織中Symmetrospora的相對豐度呈提高趨勢;施藥后18 d,健康組織與感病組織中其相對豐度達到最大值(0.09和0.82),且健康組織中其相對豐度顯著低于感病組織;此時,健康組織中毛霉屬的相對豐度也達到最大值(0.11)。
細菌優勢菌門包括變形菌門(Proteobacteria)、放線菌門(Actinobacteria)和厚壁菌門(Firmicutes),其中感病組織中變形菌門的相對豐度顯著高于健康組織。多抗霉素處理后1、3 d,變形菌門的相對豐度隨時間增加而升高,并達到峰值;施藥9 d后,其相對豐度有所下降;施藥后18 d,其相對豐度增加,感病組織變形菌門和厚壁菌門的相對豐度顯著高于健康組織。細菌屬水平優勢菌屬包括假單胞菌屬(Pseudomonas)、甲基桿菌屬(Methylobacterium)、鞘氨醇單胞菌屬(Sphingomonas)、金剛藻屬(Aureimonas)、泛菌屬(Pantoea)、根瘤菌科未知屬(unidentified_Rhizobiaceae)、沙雷氏菌屬(Serratia)、腸桿菌科未知屬(unidentified_Enterobacteriaceae)、馬賽菌屬(Massilia)、微桿菌屬(Microbacterium)。施藥后18 d,假單胞菌屬、甲基桿菌屬、鞘氨醇單胞菌屬、金剛藻屬和泛菌屬的相對豐度均有所提高,并達到最大值,同期感病組織中其相對豐度高于健康組織。
2.2.4 煙葉葉際微生物群落結構組成
由表4可知,健康組織與感病組織測序覆蓋度均達到90%以上,表明本測序結果涵蓋了絕大部分葉際微生物,可以真實、合理地反映葉際真菌和細菌的多樣性和豐富度。真菌α多樣性指數表明,施藥前,健康組織(QJDK1)葉際真菌多樣性指數(香農指數和辛普森指數)和豐富度指數(Chao1指數和ACE指數)均高于感病組織(QBDK1),但組間無顯著性差異。施藥后1 d,健康組織(QJDK2)多樣性指數達到最高值;施藥后3 d,感病組織(QBDK3)多樣性指數和豐富度指數在組內達到最高值,健康組織(QJDK3)的多樣性指數有所下降;施藥后9 d,健康組織(QJDK4)多樣性指數有所上升;施藥后18 d,健康組織(QJDK5)多樣性指數趨于平緩。
細菌α多樣性指數表明,施藥前健康組織(QJDK1)與感病組織(QBDK1)葉際細菌多樣性指數和豐富度指數均最低,且感病組織多樣性指數與豐富度指數高于健康組織,但組間無顯著性差異。采用多抗霉素進行處理后,葉際細菌多樣性指數與豐富度指數開始提高,健康組織與感病組織的多樣性指數與豐富度指數均在施藥后18 d明顯增加,且與其余不同施藥時間存在顯著差異。
2.2.5 健康與感病組織葉際微生物的互作關系
選取葉際優勢真菌與細菌屬水平top50并對其進行相關性分析,結果(圖4)表明,健康組織葉際真菌屬間關聯性大于感病組織,其絕大部分真菌屬之間呈正相關。亡革菌屬與其他菌屬之間大多呈現負相關,其中健康組織中亡革菌屬與莖點霉屬(Phoma)、鏈格孢屬和枝孢屬之間均呈負相關;感病組織中亡革菌屬與Sampaiozyma、漢納酵母屬、Papiliotrema、Vishniacozyma、Boeremia、枝孢屬和鏈格孢屬之間均呈負相關。
健康組織細菌屬之間相關性大于感病組織,多數細菌屬之間同樣呈正相關,少數細菌屬呈負相關。健康組織中甲基桿菌屬與寡養單胞菌屬(Stenotrophomonas)和阿克曼菌屬(Akkermansia)、短波單胞菌屬(Brevundimonas)與勞爾氏菌屬、假單胞菌屬與布勞特氏菌屬(Blautia)、馬賽菌屬與鞘氨醇單胞菌屬之間呈負相關,其余菌屬均呈正相關;感病組織中假單胞菌屬與金剛藻屬、勞爾氏菌屬與馬賽菌屬、甲基桿菌屬與沙雷氏菌屬之間呈負相關,其余菌屬均呈正相關。
3 結論與討論
本研究探索了多抗霉素對煙草靶斑病的田間防效,在施用多抗霉素后3、9 d,田間相對防效均達到80%以上,表明藥劑作用迅速、病斑增長速率顯著降低,病情指數得到有效控制,防治效果穩定。表明多抗霉素能快速且穩定地抑制煙草靶斑病的發生,可以廣泛應用至田間防控技術上。
葉部病害的發生常伴隨著多種病原菌的作用,葉際微生物復雜多樣,相互之間關聯緊密[34]。本研究發現,施用多抗霉素對煙草靶斑病進行防治后3 d,其煙草靶斑病病原菌立枯絲核菌的相對豐度達到最低值,表明多抗霉素能在短時間內達到較好防效。感煙草靶斑病煙葉的葉際優勢真菌屬為亡革菌屬、Symmetrospora和毛霉屬等,優勢細菌屬為假單胞菌屬、甲基桿菌屬和鞘氨醇單孢菌屬等,這一結果與前人對煙草靶斑病葉際微生物多樣性的研究結果[35]基本相同,其葉際微生物之間存在動態平衡。煙草靶斑病葉際除存在亡革菌屬外,還同時存在鏈格孢屬、假單胞菌屬、泛菌屬等微生物。施用
多抗霉素對煙草靶斑病進行防控后1、3、9、18 d,其葉際優勢真菌屬亡革菌屬的相對豐度基本呈先下降再上升的趨勢,而Symmetrospora、毛霉屬、小不整球殼屬和鏈格孢屬的相對豐度則略有增加;在優勢細菌屬中,假單胞菌屬、泛菌屬、鞘氨醇單孢菌屬、金剛藻屬和沙雷氏菌屬的相對豐度基本呈現先上升再下降的趨勢,甲基桿菌屬的相對豐度則是持續變化,但總體呈上升趨勢。韓永琴對辣椒根際微生物進行研究,結果同樣顯示,化學藥劑的施用會導致根際微生物發生變化[36]。劉亭亭等在波爾多液對煙草赤星病葉際微生物物群落結構與代謝功能的影響研究中發現,施藥后其葉際真菌群落的多樣性顯著增加[37];且波爾多液不僅作用于煙草赤星病病原菌鏈格孢屬,同樣作用于鞘脂單胞菌屬、Kosakonia等有益菌群[36-37]。本研究結果同樣顯示,用多抗霉素防治煙草靶斑病時,不僅有效作用于立枯絲核菌,同樣作用于尾孢屬、枝孢屬、甲基桿菌屬等葉際真菌和細菌,影響微生物菌群變化。
本研究結果顯示,在煙葉健康組織與感病組織中,亡革菌屬與鏈格孢屬、枝孢屬之間均呈負相關。Sun等的研究結果顯示,施用嘧菌酯防治煙草靶斑病后,其葉際微生物發生變化,亡革菌屬與鐮刀菌屬、小不整球殼屬之間呈負相關,其關聯性表現無共性,可能是由于不同菌屬之間存在競爭或共生關系,藥施后導致某些菌群的相對豐度下降,增長速率得到顯著控制,非靶標菌群相對豐度則上升[38]。不同菌屬之間同樣存在對于環境中營養物質與生長空間的競爭關系,導致施用化學藥劑進行病害防治時,可能刺激某些非靶標菌群的相對豐度提高,不同菌屬之間增長趨勢存在差異,而藥劑的施用加大了菌屬之間動態平衡的變化,而此種變化或許有利于植物代謝活動的增強,對病原菌的侵染抵抗力增強,進而影響藥劑對于植物病害的防控效率[39]。目前,在施用化學藥劑防控植物病害時,其葉際微生物的變化規律還需要進一步探索總結,了解其間微生態的變化規律有益于為病害防控以及化學藥劑的施用提供參考依據。
參考文獻:
[1]Bacharis C,Gouziotis A,Kalogeropoulou P,et al. Characterization of Rhizoctonia spp. isolates associated with damping-off disease in cotton and tobacco seedlings in greece[J]. Plant Disease,2010,94(11):1314-1322.
[2]Costa A S. Mancha aureolada erequeima do fumo causades por Corticium solani[J]. Biologicol,1948,14:113-114.
[3]吳元華,王左斌,劉志恒,等. 我國煙草新病害——靶斑病[J]. 中國煙草學報,2006(6):22,51.
[4]吳元華,趙艷琴,趙秀香,等. 煙草靶斑病原鑒定及生物學特性研究[J]. 沈陽農業大學學報,2012,43(5):521-527.
[5]李再明,楊學紅,王廟昌,等. 9種防治煙草靶斑病的藥劑篩選試驗[J]. 云南農業科技,2022(2):13-15.
[6]孫美麗,汪漢成,郭沫言,等. 4種殺菌劑對煙草靶斑病菌的抑菌活性[J]. 貴州農業科學,2022,50(9):62-68.
[7]王左斌,吳元華,趙秀香,等. “嘧肽菌凈”對煙草靶斑病的抑菌作用及田間藥效試驗[J]. 煙草科技,2007(9):61-64.
[8]周建全,張忠光,董 雪,等. 不同藥劑對煙草靶斑病的抑菌作用及田間藥效研究[J]. 安徽農業科學,2015,43(25):96-97,99.
[9]馬 欣,寇寶石,李繼博,等. 新農藥8%井岡霉素可溶液劑對煙草靶斑病的防治效果[J]. 安徽農業科學,2022,50(11):133-134.
[10]吳家全,李軍民. 多抗霉素研究現狀與市場前景[J]. 農藥科學與管理,2010,31(11):21-23.
[11]Hori M,Eguchi J,Kakiki K,et al. Studies on the mode of action of polyoxins. Ⅵ. Effect of polyoxin B on chitin synthesis in polyoxin-sensitive and resistant strains of Alternaria kikuchiana[J]. The Journal of antibiotics,1974,27(4):260-266.
[12]Nobuya O,Kazuo K,Tomomasa M. Studies on the mode of action of polyoxin D[J]. Agricultural and Biological Chemistry,2014,34(8):1224-1234.
[13]吳 祥,吉沐祥,郭玉人,等. 多抗霉素與丁子香酚混配對草莓灰霉病的防治效果[J]. 江蘇農業科學,2017,45(22):94-97.
[14]康萍芝,杜玉寧,白小軍,等. 不同化學殺菌劑與多抗霉素混配對壓砂西瓜炭疽病的聯合生物活性及田間防效[J]. 農藥,2021,60(1):70-73.
[15]韓 磊,楊超然,樊 榮,等. 幾種殺菌劑對白菜霜霉病的防治效果[J]. 農藥,2019,58(12):915-917.
[16]張海寬,周 斌.? 春雷霉素、多抗霉素防治番茄灰霉病田間藥效試驗[J]. 農藥科學與管理,2010,31(5):48-49.
[17]陶樹春,賈秀芬,李錦龍,等. 多抗霉素等6種生物農藥對溫室黃瓜白粉病的防治效果[J]. 農業科技與信息,2007(10):19.
[18]趙 綱,劉保安. 多抗霉素防治煙草赤星病的效果[J]. 煙草科技,1983(4):30-33.
[19]連 海,申昌優,鐘善良,等. 贛州煙區煙草赤星病防治藥劑篩選[J]. 安徽農業科學,2020,48(22):148-149,165.
[20]Wang R,Zhang H C,Sun L G,et al. Microbial community composition is related to soil biological and chemical properties and bacterial wilt outbreak[J]. Scientific Reports,2017,7:343.
[21]Yang H W,Li J,Xiao Y H,et al. An integrated insight into the relationship between soil microbial community and tobacco bacterial wilt disease[J]. Frontiers in Microbiology,2017,8:2179.
[22]Dai Y F,Wu X M,Wang H C,et al. Spatio-gemporal variation in the phyllospheric microbial biodiversity of Alternaria alternata-infected tobacco foliage[J]. Front Microbiol 2022,13:920109.
[23]劉亭亭,曾隕濤,汪漢成,等. 赤星病發生期不同成熟度煙葉葉際微生物代謝與群落結構[J]. 中國煙草科學,202 2(6):22-29.
[24]Sun M L,Shi C H,Huang Y,et al. Effect of disease severity on the structure and diversity of the phyllosphere microbial community in tobacco[J]. Frontiers in Microbiology,2023,4(13):1081576.
[25]刁春玲,劉 芳,宋寶安. 農用殺菌劑作用機理的研究進展[J]. 農藥,2006(6):374-377.
[26]Huang Y,Wang H C,Cai L T,et al. Phyllospheric microbial composition and diversity of the tobacco leaves infected by Didymella segeticola[J]. Frontiers in Microbiology,2021,12:699699.
[27]向立剛,汪漢成,鄭 蘋,等. 一種未知真菌性葉斑病發病煙葉真菌群落分析[J]. 中國煙草科學,202 2(1):40-46.
[28]劉天波,滕 凱,周向平,等. 拮抗菌群對煙草野火病的防治效果及葉際微生物群落多樣性的影響[J]. 微生物學通報,202 8(8):2643-2652.
[29]劉 暢,汪漢成,謝紅煉,等. 感赤星病煙葉的真菌群落結構分析[J]. 貴州農業科學,2019,47(7):54-59.
[30]劉 暢,汪漢成,謝紅煉,等. 感染赤星病煙草葉際細菌的多樣性分析[J]. 煙草科技,2020,53(2):8-14.
[31]譚舒心. 蜘蛛蘭褐斑病及其病原菌鑒定與病害的藥劑控制試驗[D]. 重慶:西南大學,2009.
[32]劉斯泓,紀明山. 防治煙草靶斑病的復配藥劑配方篩選及田間藥效試驗[J]. 江蘇農業科學,2014,42(6):140-143.
[33]Haas B J,Gevers D,Earl A M,et al. Chimeric 16S rRNA sequence formation and detection in Sanger and 454-pyrosequenced PCR amplicons[J]. Genome Res 2011,21(3):494-504.
[34]刁春玲,劉 芳,宋寶安. 農用殺菌劑作用機理的研究進展[J]. 農藥,2006(6):374-377.
[35]孫美麗,史彩華,肖本青,等. 煙草靶斑病葉際微生物群落結構與多樣性分析[J]. 煙草科技,2023,56(4):1-9.
[36]韓永琴. 多黏類菌劑和化學藥劑對辣椒疫病的防效及對根際微生物的影響[D]. 長沙:湖南農業大學,2019.
[37]劉亭亭,汪漢成,孫美麗,等. 波爾多液對煙草葉際微生物群落結構與代謝功能的影響[J]. 農藥學學報,2022,24(6):1446-1455.
[38]Sun M L,Wang H C,Shi C H,et al. Effect of azoxystrobin on tobacco leaf microbial composition and diversity[J]. Frontiers in Plant Science,2023,13:1101039.
[39]Zeng Y T,Xiong T,Bai Z Q,et al. Establishment of the sensitivity baseline of Diaporthe citri population to azoxystrobin (in Chinese)[J]. Plant Protection,2022,48(4):138-142.
猜你喜歡
熱帶農業科學(2016年11期)2017-01-21 14:59:21
熱帶農業科學(2016年11期)2017-01-21 14:57:08
河北漁業(2015年12期)2015-12-21 14:37:31
河北漁業(2015年11期)2015-11-18 16:39:54
河北漁業(2015年11期)2015-11-18 16:37:49
河北漁業(2015年10期)2015-10-15 11:14:57
湖北農業科學(2015年18期)2015-10-13 21:34:01
河北漁業(2015年7期)2015-07-21 11:06:52
河北漁業(2015年6期)2015-06-24 11:04:37
安徽農學通報(2015年5期)2015-03-23 23:51:20
