雞腺苷琥珀酸裂解酶互作蛋白的篩選及其功能分析
2024-04-08 13:51:06陳友波石鈺仕趙德鵬譚啟松
浙江農(nóng)業(yè)學報 2024年3期
余 歡,李 輝,陳友波,石鈺仕,趙德鵬,龍 霞,譚啟松
(貴州大學 動物科學學院,高原山地動物遺傳育種與繁殖教育部重點實驗室,貴州 貴陽 550025)
肌苷酸(inosine monophosphate, IMP)作為眾多畜禽肉質(zhì)風味評價指標之一,其含量是目前衡量肉類新鮮程度的重要指標,是公認的影響肌肉風味的重要物質(zhì)。IMP來源是2個體內(nèi)合成途徑和三磷酸腺苷的分解過程,合成途徑包括以氨基酸、磷酸核糖、一碳單位和CO2等原料為主的從頭合成途徑[1]和體內(nèi)嘌呤核苷或游離嘌呤合成IMP的補救途徑[2]。腺苷琥珀酸裂解酶(adenylosuccinate lyase,ADSL)是IMP合成酶系中催化合成嘌呤核苷酸起始合成與循環(huán)中的唯一雙功能酶[3],是畜禽體內(nèi)IMP合成的關鍵酶。因此,編碼腺苷琥珀酸裂解酶的ADSL基因被認為是影響肌肉IMP含量的主效基因[4]。相關研究表明,ADSL基因與畜禽IMP含量具有顯著相關性[5-7]。張梅[8]對玫瑰冠雞的研究結(jié)果表明,ADSL基因的表達與肌肉的IMP含量呈顯著正相關。劉長青等[9]用熒光定量PCR檢測了ADSL基因在北京油雞的心、肝、脾、肺、腎、腿肌、胸肌和腦等組織的表達情況,結(jié)果顯示,ADSL基因在胸肌中表達水平最高。Yuan等[10]研究顯示,草魚ADSL基因在肌肉中的表達水平最高,而在魚鰓中表達量最低。以上研究表明,ADSL基因與畜禽肉質(zhì)相關,其表達水平在不同組織中均有差異,但是其在肌肉中明顯高表達。目前,關于ADSL蛋白對肌肉風味的影響機制報道較少。
赤水烏骨雞,又名“竹香雞”,原產(chǎn)于貴州省赤水市,因其肉質(zhì)鮮美、風味獨特、營養(yǎng)豐富,受到廣大消費者喜愛。本研究以赤水烏骨雞為對象,采用免疫共沉淀(co-immunoprecipitation, Co-IP)聯(lián)合高效液相色譜串聯(lián)質(zhì)譜聯(lián)用技術(LC-MS/MS)進行蛋白質(zhì)譜分析,篩選出與雞ADSL蛋白相互作用的細胞蛋白,并進行互作蛋白的生物信息學分析,旨在為進一步深入探究雞ADSL基因在肌肉的表達調(diào)控機制提供參考。
1 材料與方法
1.1 材料
pEGFP-C1質(zhì)粒來自貴州大學動物遺傳育種與繁殖教育部重點實驗室,細胞來源為本課題組保存的原代細胞,大腸埃希菌感受態(tài)TOP10細胞購自貴州宏達爾生物科技有限公司,ADSL基因的干擾載體為本課題組前期構(gòu)建、保存[11]。
1.2 主要試劑
DNA凝膠回收試劑盒和無內(nèi)毒素小提中量試劑盒-離心柱型購于天根生化科技(北京)有限公司;4%多聚甲醛、1×RIPA裂解液(弱)、TriX-100通透液、蛋白酶抑制劑(PMSF)、1×PBS緩沖液、脫脂奶粉、DMEM-F12基礎培養(yǎng)基購于北京索萊寶科技有限公司;兔抗綠色熒光蛋白(GFP)單克隆抗體購自Proteintech公司(美國);辣根過氧化物酶(HRP)結(jié)合的親和山羊抗兔抗體(H+L)購自ABclonal公司;RevertAid First Strand cDNA Synthesis Kit、T4 DNA連接酶、DNA Marker、PfuTaqDNA高保真聚合酶、限制性內(nèi)切酶(XhoⅠ和EcoRⅠ)、Pierce經(jīng)典磁珠式免疫沉淀試劑盒、LipofectamineTM 3000 轉(zhuǎn)染試劑盒購自Thermo Fisher公司;細胞轉(zhuǎn)染試劑使用Neofect DNA轉(zhuǎn)染試劑盒;細胞核染料DAPI、SDS-PAGE蛋白上樣緩沖液(5×)和ECL發(fā)光顯色液購于上海碧云天生物技術公司;聚偏二氟乙烯膜(polyvinylidene fluoride, PVDF)和胎牛血清購自GIBCO公司;TRIzol購自南京諾唯贊生物科技股份有限公司;Opti-MEMTM購自貴州格瑞恩公司;anti-desmin、山羊抗兔IgG-cy3購自北京博奧森生物技術有限公司;其他試劑均為國產(chǎn)分析純。
1.3 雞ASDL基因真核表達載體的構(gòu)建和鑒定
根據(jù)NCBI網(wǎng)站的雞ADSL基因序列(基因登錄號:NM_205529.2),利用primer 5.0軟件篩選出酶切位點(XhoⅠ和EcoRⅠ),根據(jù)ADSL基因的蛋白質(zhì)編碼序列(coding sequence, CDS)和酶切位點設計引物(上游引物序列5′-TGGACGAGCTGTACAAGTCCGGACTCAGATCTCGAG-GCATGGCGACCCCCTGCGCCGAGGAGGACCCG-C-3′,帶下劃線的序列是XhoⅠ酶切位點;下游引物序列5′-GATCCCGGGCCCGCGGTACCGTCGA-CTGCAGAATTCTTAAAGTGTCAGCTCAATTTTC-CCACCCATCAT-3′,帶下劃線的序列是EcoRⅠ酶切位點),由北京擎科生物科技股份有限公司合成相關引物。
提取赤水烏骨雞成肌細胞總RNA,逆轉(zhuǎn)錄成cDNA后,擴增ADSL基因。利用無縫克隆試劑盒將純化后的PCR產(chǎn)物和酶切后的pEGFP-C1表達載體連接并轉(zhuǎn)化進大腸埃希菌感受態(tài)TOP10細胞中。利用質(zhì)粒提取試劑盒提取陽性單克隆菌種的質(zhì)粒DNA,進行雙酶切驗證。將酶切驗證正確的質(zhì)粒送北京擎科生物科技股份有限公司進行測序驗證。
1.4 細胞培養(yǎng)與鑒定
復蘇赤水烏骨雞成肌細胞,使用Desmin抗體對成肌細胞的肌間線蛋白(desmin)進行鑒定。在含5% CO2的37 ℃細胞培養(yǎng)箱中,用含15%胎牛血清的DMEM-F12培養(yǎng)基(添加1%青霉素和1%鏈霉素)在六孔板中培養(yǎng)赤水烏骨雞成肌細胞,當六孔板中的成肌細胞密度生長到80%左右,棄去原有的培養(yǎng)基,加入無菌1×PBS沖洗3次;每孔加入800 μL預冷的4%多聚甲醛后固定30 min,清洗3次后加入800 μL 0.2%(體積分數(shù))的Triton X-100溶液,室溫孵育15 min;清洗3次后加入200 μL稀釋的山羊血清(稀釋比1∶500,每孔),37 ℃封閉30 min;清洗3次后加入100 μL一抗Desmin(1∶500稀釋),4 ℃冰箱內(nèi)孵育過夜。棄掉一抗清洗3次,在避光的暗室,加入100 μL Cy3標記的山羊抗兔IgG-Cy3(1∶500稀釋),37 ℃孵育1 h。清洗3次后加入200 μL DAPI細胞核染料,室溫孵育5~7 min,再次清洗3次,最后加入600 μL抗熒光猝滅劑,在熒光倒置顯微鏡下觀測。
1.5 轉(zhuǎn)染和融合蛋白的Wester blot檢測
將成肌細胞置于細胞六孔板中,用含15%胎牛血清、1%青霉素和1%鏈霉素的DMEM-F12培養(yǎng)基,置于37 ℃,在含5% CO2的細胞培養(yǎng)箱培養(yǎng)。待細胞密度達到80%左右,提前2 h將待轉(zhuǎn)染細胞的培養(yǎng)基更換成同等濃度不含雙抗的DMEM-F12培養(yǎng)基。將2 μg提取的質(zhì)粒DNA用100 mL OPTI-MEM溶液稀釋,充分混勻;向混合液中加入2 μL Neofect DNA轉(zhuǎn)染試劑,混勻后,室溫靜置15~30 min。將轉(zhuǎn)染復合物加入細胞培養(yǎng)基中后放入培養(yǎng)箱中培養(yǎng)24 h。
將pEGFP-C1和pEGFP-C1-ADSL轉(zhuǎn)染后的成肌細胞用PBS清洗3次,加入1×RIPA細胞裂解液進行裂解,裂解30 min后,使用細胞刮收集細胞,4 ℃、12 000×g離心10 min,收集細胞總蛋白。加入適量的蛋白上樣緩沖液,煮沸后進行SDS-PAGE凝膠電泳分離。通過濕轉(zhuǎn)轉(zhuǎn)移系統(tǒng)(BIO-RAD,美國)將蛋白轉(zhuǎn)印至PVDF膜上,用含TBST(TBS+Tween)的5%脫脂奶粉封閉2 h;棄掉液體,加入anti-GFP兔單克隆抗體,4 ℃搖床過夜孵育;用TBST溶液清洗3次后,加入HRP標記山羊抗兔IgG (H+L),室溫孵育1.5 h;用TBST溶液清洗3次后,使用ECL發(fā)光液進行顯色并拍照。
1.6 重組蛋白的亞細胞定位分析
將轉(zhuǎn)染pEGFP-C1和pEGFP-C1-ADSL的成肌細胞用PBS洗3次,依次加入預冷的4%多聚甲醛,室溫固定30 min,然后用3% Triton X-100通透處理10 min,再用PBS清洗3次,加入稀釋后的DAPI細胞核染料,靜置8 min后用PBS清洗3次,最后置于倒置熒光顯微鏡下觀察結(jié)果。將獲得的熒光圖片用Photoshop CS6進行合并(merge)處理,以熒光共定位的方法來確定重組蛋白的亞細胞定位情況。
1.7 免疫共沉淀
把成肌細胞接種于六孔板,待細胞密度達80%左右時,分別將pEGFP-C1-ADSL和pEGFP-C1轉(zhuǎn)染成肌細胞。在細胞轉(zhuǎn)染后24 h,用PBS清洗細胞3次,然后用免疫沉淀裂解緩沖液裂解細胞,4 ℃、13 000×g離心10 min后,收集上清液,向每個樣品中加入10 μg anti-GFP兔單克隆抗體,混合均勻后在4 ℃過夜。向樣品中加入25 μL Pierce蛋白A/G,混勻后室溫孵育1 h。用磁力架收集磁珠,經(jīng)過免疫沉淀裂解緩沖液清洗磁珠3次后,加入500 μL超純水輕柔混勻,棄上清液。重復2次后,向磁珠中加入100 μL蛋白上樣緩沖液,室溫靜置10 min,分離磁珠,保留上清液。
1.8 LC-MS/MS分析
蛋白質(zhì)復合物送至金開瑞生物工程有限公司(武漢)用于LC-MS/MS質(zhì)譜鑒定。將蛋白樣品進行還原烷基化和酶解處理,酶解產(chǎn)生的多肽用C18柱子除鹽,抽干后用15 μL上樣緩沖液(含0.1%甲酸和3%乙腈)溶解多肽,上LC-MS/MS(ekspertTM nanoLC,AB Sciex TripleTOF5600-plus)儀器進行分析,LC-MS/MS下機后,將原始下機數(shù)據(jù)提交到Proteinpilot 5.0 (Applied Biosystems,Sciex)軟件中進行數(shù)據(jù)庫檢索。在設置可信度conf≥95%、unique peptides≥1時,檢測樣品中所含蛋白質(zhì)的種類與數(shù)量。
1.9 細胞蛋白的生物信息學分析
利用維恩圖(Venn圖)在線軟件(http://bioinformatics.psb.ugent.be/webtools/Venn/)繪制C1標簽組和C1-ADSL重組蛋白組交集的蛋白數(shù)量;利用在線網(wǎng)站UniProt (https://www.uniprot.org)進行蛋白的基因本體(gene ontology, GO)注釋(包含生物學進程、細胞組分和分子功能);利用KEGG(Kyoto Encyclopedia of Genes and Genomes)數(shù)據(jù)庫(http://www.genom e.jp/kegg/mappe r.html)分析蛋白參與的信號通路;利用STRING Version 11.0在線網(wǎng)站(https://string-db.org/)進行蛋白互作網(wǎng)絡圖的繪制。
1.10 ADSL基因與相關基因之間的調(diào)控表達研究
根據(jù)生物信息學分析,篩選出RPL4、PDLIM5、ACTG1、SRSF10這4個基因,設計這些基因的特異性引物,通過在成肌細胞中轉(zhuǎn)染ADSL過表達載體和用LipofectamineTM 3000 轉(zhuǎn)染ADSL干擾載體后(方法參考說明書),檢測ADSL過表達和沉默狀態(tài)下成肌細胞中RPL4、PDLIM5、ACTG1和SRSF10的表達情況,探究ADSL基因與它們之間的調(diào)控關系。
2 結(jié)果與分析
2.1 雞ADSL基因重組真核表達載體的鑒定
以逆轉(zhuǎn)錄合成的cDNA為模板,使用特異性引物PCR擴增雞ADSL基因的CDS,利用無縫克隆技術導入pEGFP-C1載體質(zhì)粒中。電泳檢測結(jié)果顯示,目的片段長度為1 495 bp(圖1-A),與預期結(jié)果一致。對pEGFP-C1-ADSL載體進行XhoⅠ和EcoRⅠ雙酶切,結(jié)果顯示,pEGFP-C1-ADSL酶切后分別獲1 495 bp和4 686 bp的目的基因條帶,與預測大小一致(圖1-B)。測序結(jié)果表明,雞ADSL基因重組真核表達載體pEGFP-C1-ADSL構(gòu)建成功。
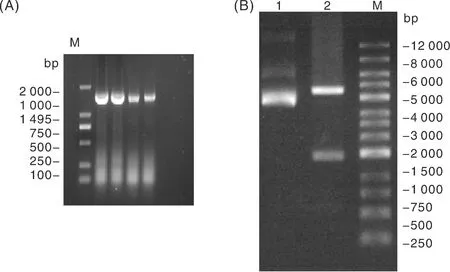
A,雞ADSL基因的PCR擴增圖;B,重組真核表達載體雙酶切鑒定圖,其中1號泳道為pEGFP-C1質(zhì)粒,2號泳道為重組質(zhì)粒pEGFP-C1-ADSL的雙酶切。M為DNA marker。A, PCR amplification plot of chicken ADSL gene; B, double-enzyme identification plot of recombinant eukaryotic expression vector. In which, lane 1 was tpEGFP-C1 plasmid, lane 2 was the biallelic graph of recombinant plasmid pEGFP-C1-ADSL. M was DNA marker.圖1 雞ADSL基因PCR擴增和重組質(zhì)粒的雙酶切鑒定結(jié)果Fig.1 PCR amplification of chicken ADSL gene and double digestion identification of recombinant plasmids
2.2 雞成肌細胞的培養(yǎng)與鑒定
對赤水烏骨雞成肌細胞進行免疫熒光鑒定,結(jié)果如圖2所示:圖2-A是明場下的赤水烏骨雞成肌原代細胞;圖2-B是使用DAPI進行核染后的赤水烏骨雞成肌細胞,可見細胞核顏色為明亮的藍色;圖2-C為使用一抗與成肌細胞進行孵育,一抗與成肌細胞中的desmin(肌間線蛋白)相結(jié)合,再與二抗(Cy3標記的山羊抗兔IgG-Cy3)發(fā)生作用后,進行熒光檢測可見成肌細胞被染為明亮的紅色,圖2-D是圖2-B與圖2-C的疊加圖,可見紅色部分為成肌細胞中的desmin,表明成功復蘇赤水烏骨雞成肌細胞。
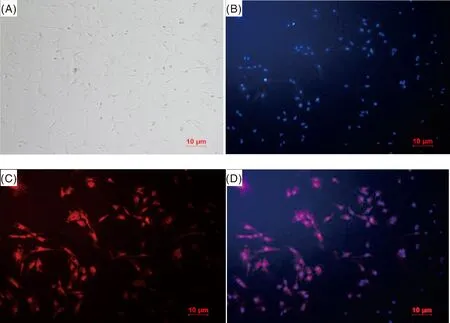
A,明場圖;B,DAPI核染圖;C,一抗反應熒光圖;D,B與C的疊加。A, Bright-field picture; B, DAPI nuclear staining picture; C, Fluorescence plot of primary antibody reaction; D, Superimposition of B and C.圖2 赤水烏骨雞成肌細胞鑒定結(jié)果Fig.2 Identification results of myoblasts of Chishui black-bone chicken
2.3 雞ADSL基因在細胞中的表達
將pEGFP-C1和pEGFP-C1-ADSL轉(zhuǎn)染赤水烏骨雞成肌細胞后,對重組蛋白進行檢測,結(jié)果顯示,獲得了大小為82 ku左右的pEGFP-C1-ADSL重組蛋白條帶(圖3),與預測大小(81.9 ku)相一致。

M為蛋白質(zhì)分子量標準;1、2、3號泳道為pEGFP-C1蛋白印跡圖;4、5、6泳道為pEGFP-C1-ADSL蛋白印跡圖。M was protein marker; Lanes 1, 2, and 3 were pEGFP-C1 protein blots, and lanes 4, 5 and 6 were pEGFP-C1-ADSL protein blots.圖3 pEGFP-C1和pEGFP-C1-ADSL在成肌細胞中的蛋白質(zhì)印跡Fig.3 Western blotting of pEGFP-C1 and pEGFP-C1-ADSL in myoblasts
2.4 雞ADSL的亞細胞定位
對pEGFP-C1標簽蛋白和pEGFP-C1-ADSL重組蛋白的亞細胞定位進行分析,結(jié)果發(fā)現(xiàn),EGFP標簽蛋白主要定位在細胞質(zhì),而ADSL蛋白在細胞核和細胞質(zhì)均存在(圖4)。
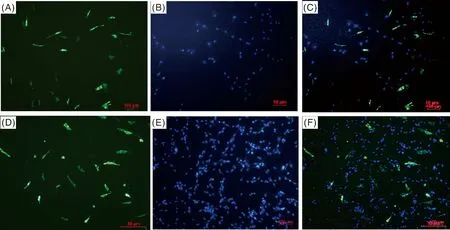
圖A、B、C為pEGFP-C1標簽蛋白,圖D、E、F為pEGFP-C1-ADSL重組蛋白。綠色是分別用pEGFP-C1和pEGFP-C1-ADSL轉(zhuǎn)染成肌細胞后的發(fā)光效果圖,藍色是DAPI核染結(jié)果。Figures A, B and C were pEGFP-C1 tag protein, and Figures D, E and F were pEGFP-C1-ADSL recombinant protein. The green color was the luminescence effect of myoblasts transfected with pEGFP-C1 and pEGFP-C1-ADSL respectively, and the blue color was the DAPI nuclear staining result.圖4 pEGFP-C1和pEGFP-C1-ADSL在成肌細胞中的亞細胞定位Fig.4 Subcellular localization of pEGFP-C1 and pEGFP-C1-ADSL in myoblasts
2.5 互作蛋白鑒定
利用LC-MS/MS技術把與GFP-ADSL重組蛋白和GFP標簽蛋白結(jié)合的蛋白進行蛋白質(zhì)譜鑒定,在可信度conf≥95、unique peptides≥1時,樣品ADSL(GFP-ADSL)、C1(GFP-C1)質(zhì)譜產(chǎn)生的二級譜圖數(shù)分別為16 057、13 090,解析的二級譜圖數(shù)分別為2 616、1 593,分別鑒定出916、598個肽段數(shù)。另外,3次重復實驗中至少出現(xiàn)2次的蛋白被視為存在相互作用,結(jié)果發(fā)現(xiàn)與GFP-C1標簽相互作用的蛋白有161個,與GFP-ADSL重組蛋白相互作用的細胞蛋白有197個。使用Venn在線軟件分析發(fā)現(xiàn),有94個蛋白可能與ADSL蛋白存在相互作用關系(圖5),部分蛋白信息見表1。
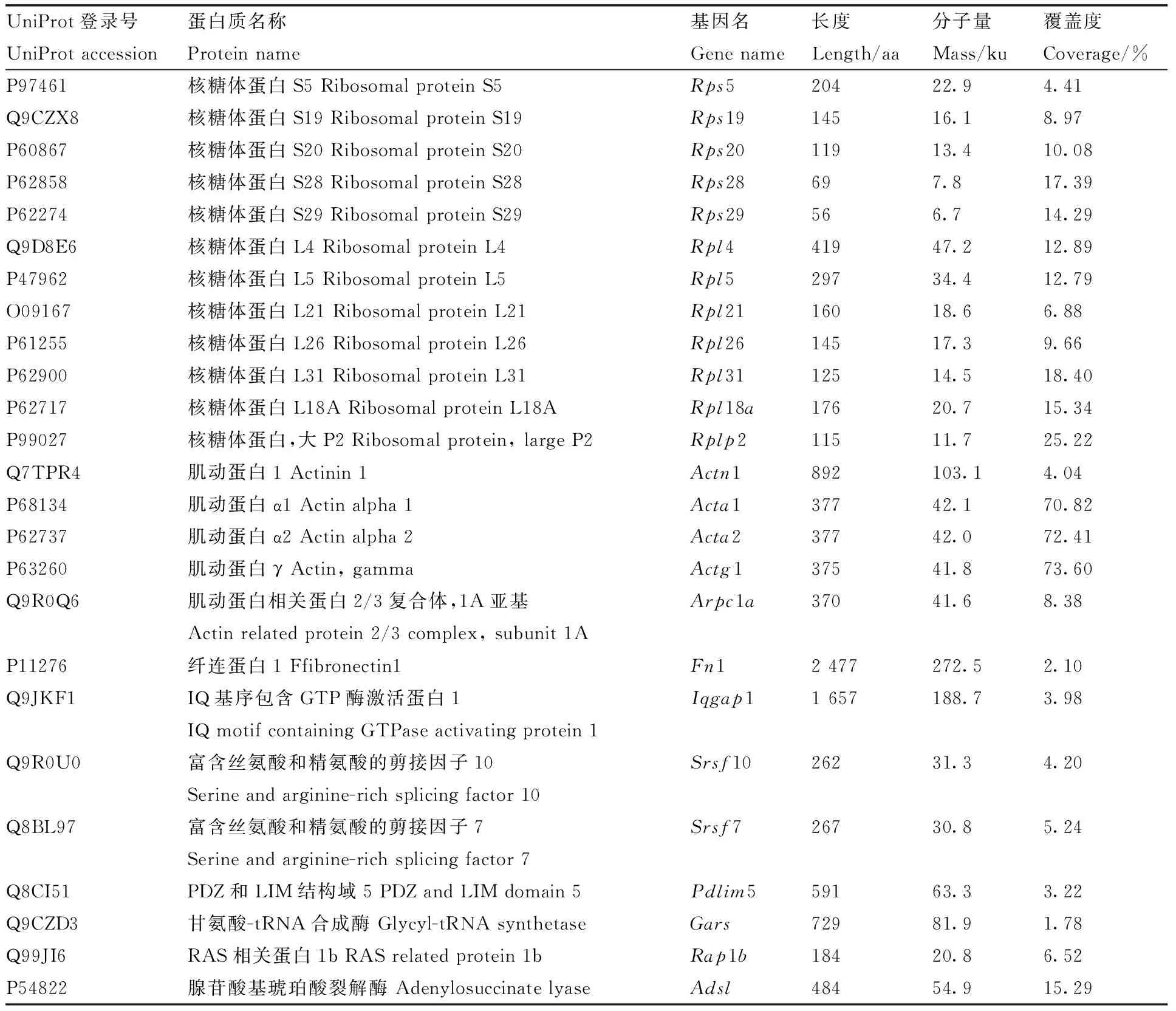
表1 與雞ADSL蛋白相互作用的部分蛋白列表Table 1 List of selected proteins interacting with chicken ADSL protein
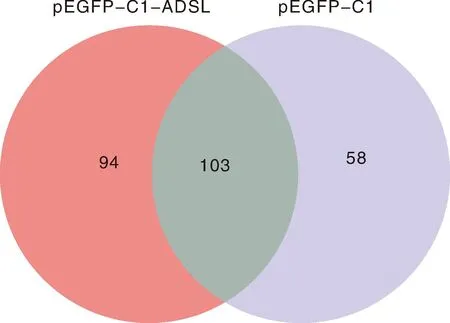
圖5 pEGFP-C1-ADSL和pEGFP-C1互作蛋白的Venn圖Fig.5 Venn diagram of pEGFP-C1-ADSL and pEGFP-C1 interacting proteins
2.6 GO功能注釋
GO功能注釋表明,大部分互作細胞蛋白具有多種功能,參與多種細胞內(nèi)生物反應。生物過程富集結(jié)果表明,它們主要參與細胞進程、生物調(diào)節(jié)和刺激反應等生物過程;細胞組分分析顯示,這些蛋白主要定位于細胞結(jié)構(gòu)體、胞內(nèi)和含蛋白質(zhì)的復合物上;雞ADSL蛋白富集的分子主要發(fā)揮結(jié)合、結(jié)構(gòu)分子活性和催化活性的分子功能(圖6)。
2.7 與雞ADSL蛋白互作的蛋白的KEGG通路分析
利用KEGG信號通路對質(zhì)譜鑒定到的互作蛋白進行通路富集,獲得的信號通路見圖7。與雞ADSL蛋白互作的蛋白同時參與多條信號通路,主要參與代謝、遺傳信息處理、環(huán)境信息處理、細胞過程、有機系統(tǒng)和疾病等信號通路。
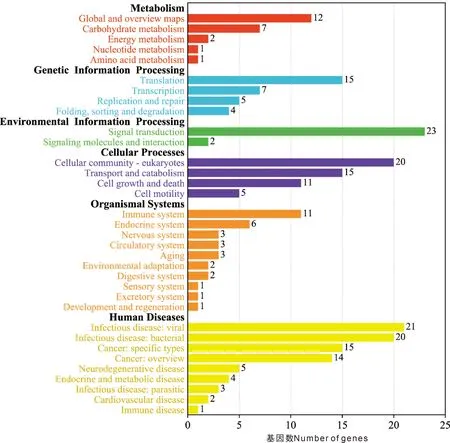
Metabolism,代謝;Genetic information processing,遺傳信息處理;Environmental information processing,環(huán)境信息處理;Cellular processes,細胞過程;Organismal systems,有機系統(tǒng);Human diseases,人類疾病。圖7 與雞ADSL蛋白互作的蛋白的KEGG通路分析Fig.7 KEGG pathway analysis of proteins interacting with chicken ADSL protein
2.8 雞ADSL蛋白的蛋白質(zhì)-蛋白質(zhì)相互作用(PPI)網(wǎng)絡圖
為研究雞ADSL蛋白相互作用網(wǎng)絡信息,利用在線軟件STRING繪制PPI網(wǎng)絡圖。將上文得到的94個蛋白導入STRING軟件,發(fā)現(xiàn)有9個蛋白沒有相關信息(圖8-A)。通過知網(wǎng)查詢與ADSL相關的基因,引入61個不在質(zhì)譜數(shù)據(jù)的蛋白,得到由146個蛋白構(gòu)成的共1 261個網(wǎng)絡節(jié)點蛋白互作圖(圖8-B)。為得到更清晰的蛋白信息圖,利用Cytoscape對該結(jié)果進行cytoHubba數(shù)據(jù)優(yōu)化,結(jié)果見圖8-C,圓圈越往內(nèi)表示這些蛋白互作關系越強烈,顏色越深表示相關性越強。
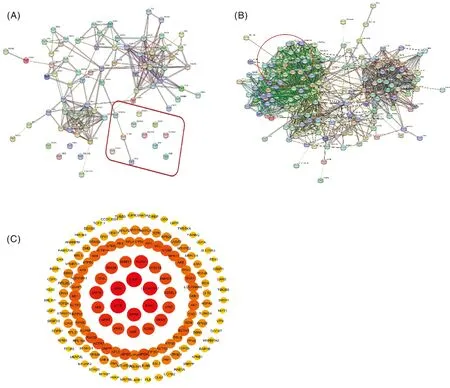
A,引入61個蛋白前;B,引入61蛋白后;C,Cytoscape優(yōu)化后。A, Before the introduction of 61 proteins; B, After the introduction of 61 proteins; C, Cytoscape optimized.圖8 雞ADSL蛋白的PPI網(wǎng)絡圖Fig.8 PPI network diagram of chicken ADSL protein
2.9 ADSL基因表達對相關基因表達的影響
ADSL基因過表達時,成肌細胞內(nèi)RPL4、PDLIM5基因表達水平顯著(P<0.05)降低,ACTG1、SRSF10基因表達水平極顯著(P<0.01)升高(圖9-A);當ADSL基因沉默后,成肌細胞內(nèi)RPL4和ACTG1基因的表達水平極顯著(P<0.01)上調(diào),PDLIM5、SRSF10基因表達水平無顯著變化(圖9-B)。
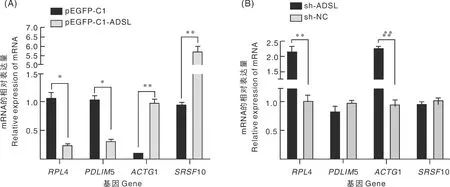
*表示差異顯著(P<0.05);**表示差異極顯著(P<0.01)。* and ** meant significant differences at the levels of P<0.05 and P<0.01, respectively.圖9 ADSL基因過表達(A)和沉默(B)對相關基因表達的影響Fig.9 Effect of ADSL gene overexpression (A) and silencing (B) on the expression of related genes
3 討論
ADSL作為調(diào)控家禽IMP的重要候選基因,對家禽肉質(zhì)品質(zhì)的改良有著重要參考價值[12]。目前關于ADSL基因的研究多集中在單核苷酸多態(tài)性和IMP相關性的研究上,欒德琴[13]的研究表明,雞ADSL基因表達水平較高,屬于影響IMP沉積的重要基因。ADSL基因主要在肌肉中,尤其是家禽胸肌中高表達。本研究以赤水烏骨雞成肌細胞為研究對象,利用Co-IP聯(lián)合LC-MS/MS的方法,檢測與雞ADSL相互作用的蛋白,進而研究其在胸肌中高表達的原因與機制。
真核表達載體pEGFP-C1是一種自帶綠色熒光的蛋白,可以與目的基因形成融合蛋白,并在真核細胞內(nèi)高表達[14],利用pEGFP-C1來研究目的基因編碼的蛋白的生物學功能是現(xiàn)在常用的一種實驗手段。間接免疫熒光實驗結(jié)果顯示,赤水烏骨雞pEGFP-C1-ADSL融合蛋白定位于細胞質(zhì)和細胞核。Zhou等[15]通過在DF-1細胞內(nèi)過表達GFP和GFP-CHBRD2,利用Co-IP聯(lián)合LC-MS/MS蛋白質(zhì)譜鑒定技術,鑒定到233個與GFP互作的蛋白,414個與GFP-CHBRD2互作的蛋白;Venn分析發(fā)現(xiàn)只有225個蛋白與GFP-CHBRD2有互作關系。本實驗通過Co-IP聯(lián)合LC-MS/MS蛋白質(zhì)譜鑒定技術,在pEGFP-C1蛋白樣品中鑒定到161個蛋白,pEGFP-C1-ADSL樣品中鑒定到197個蛋白,二者之間有103個共有蛋白,Venn結(jié)果顯示,只有94個蛋白與pEGFP-C1-ADSL有互作關系。蛋白質(zhì)之間通過形成蛋白復合物在特定的時空內(nèi)完成特定的功能,研究蛋白質(zhì)相互作用對于全面揭示蛋白質(zhì)的功能是必要的[16]。這些互作蛋白主要參與代謝、遺傳信息處理、環(huán)境信息處理、細胞過程、有機系統(tǒng)和疾病等幾種信號通路,這些蛋白大部分具有復合功能,參與多條通路,在多種生物功能中都在發(fā)揮作用。
本次鑒定到12個蛋白屬于核糖體蛋白家族的成員,它們分別是Rps5、Rps19、Rps20、Rps28、Rps29,以及Rpl4、Rpl5、Rpl21、Rpl26、Rpl31、Rpl18a和Rplp2,它們不但參與蛋白質(zhì)的生物合成,同時參與細胞的分裂與分化,對細胞增殖和細胞凋亡也起到重要的調(diào)節(jié)作用[17],對基因的表達發(fā)揮著關鍵調(diào)控功能。其次,Actg1、Actn1、Arpc1a、Fn1和Iqgap1參與肌動蛋白細胞骨架調(diào)節(jié)。SRSF10能夠促進成肌細胞的分化,在橫紋肌發(fā)育、成肌細胞分化中發(fā)揮重要調(diào)控作用[18],對肌肉的發(fā)育有著重要意義。在這些蛋白中,Gars[19]、Rap1b[20]、Srsf10[21-22]、Acta1[23-24]、Rpl26[25]、Pdlim5[26]、Actg1[27]、Copb1[28]等蛋白都與畜禽肉質(zhì)風味等相關,以上8個蛋白可能與雞ADSL蛋白存在互作關系。ADSL基因過表達時,RPL4的表達受到了抑制,將ADSL進行沉默以后,RPL4的表達水平顯著上升,這可能與RPL4的高度保守性有關,表明ADSL與RPL4在赤水烏骨雞成肌細胞中可能存在反向調(diào)控的關系。ACTG1不僅位于肌細胞,也存在在非肌細胞內(nèi),它不僅參與細胞分裂,還在細胞運動和遷移等活動中發(fā)揮功能。ADSL處于過表達和沉默時,ACTG1的表達均表現(xiàn)為上調(diào),ADSL作為調(diào)控風味的主效基因在肌肉中出現(xiàn)了高表達的現(xiàn)象,當ADSL過表達時,ACTG1表達上調(diào),當ADSL沉默時,ACTG1表達依舊上調(diào),可能是因為ACTG1是一類作為細胞骨架成分而存在的蛋白質(zhì),具有高度保守性,它作為內(nèi)部細胞運動的介質(zhì)而存在,對維持細胞骨架和肌細胞的正常發(fā)育均起到重要作用。
4 結(jié)論
本研究構(gòu)建了雞ADSL基因的重組真核表達載體pEGFP-C1-ADSL,并將該載體轉(zhuǎn)入赤水烏骨雞的成肌細胞,隨后利用Co-IP聯(lián)合LC-MS/MS技術鑒定到94個與雞ADSL存在互作的蛋白。pEGFP-C1-ADSL重組蛋白于細胞核和細胞質(zhì)。GO功能注釋表明,這些蛋白在細胞進程、生物調(diào)節(jié)和刺激反應等生物過程中發(fā)揮作用;KEGG信號通路分析顯示,這些蛋白主要參與代謝、遺傳信息處理、環(huán)境信息處理、細胞過程、有機系統(tǒng)和疾病等通路。
