棗樹novel-miR16靶基因ZjTCP4鑒定及生物信息學分析
2024-04-08 13:51:10張露荷王多鋒張廣忠呂斌燕張洋軍
浙江農業學報 2024年3期
張露荷,王多鋒,張 德,張廣忠,趙 通,呂斌燕,張洋軍,李 毅
(1.甘肅省林業科學研究院,甘肅 蘭州 730020; 2.甘肅農業大學 林學院,甘肅 蘭州 730070)
干旱是世界上最嚴重的自然災害之一。我國地處典型季風氣候區,干旱災害的影響尤為突出[1]。干旱缺水是限制沙漠、戈壁區農業發展的關鍵因素,棗樹作為一種抗旱性較強的經濟林樹種,其林地也出現了嚴重的土壤干燥化問題,給棗樹正常生長造成了障礙。棗樹花小、人工去雄困難、坐果率低且胚敗育現象嚴重,使得棗樹雜交育種非常困難,所以生物技術育種是棗樹未來育種的主要途徑。前人通過轉錄組分析,鑒定了許多參與干旱響應的基因,如棗樹的抗壞血酸過氧化物酶基因(ZjAPX基因)[2],耐旱相關的轉錄因子有DREB、NAC、MYB、TCP等,但對這些基因的轉錄后調控機理還不清楚。miRNA是一類非編碼小分子RNA,通常在轉錄后水平調控靶基因的表達,參與植物的生長發育,響應著各種非生物脅迫和生物脅迫。雖然已經報道了棗樹棗瘋病[3]、酸棗在鹽脅迫下miRNA的一些研究[4],證實miRNA在調控植物干旱脅迫反應中發揮著重要的調控作用[5],但關于miRNA調控棗樹響應干旱脅迫的研究未見報道。
TCPs是一類植物特有的轉錄因子,通過調控下游基因的表達,在調控植物的形態構型、配子體發育、晝夜節律以及脅迫響應等多種生命活動過程發揮著重要作用;是響應干旱脅迫、鹽脅迫、高溫脅迫等非生物脅迫的十分重要的轉錄因子。已有研究表明,TCPs轉錄因子主要通過調節細胞增殖和植物激素途徑來調控植物器官發生和形態構型[6]。最近研究解析了TCP基因編碼蛋白與下游靶基因啟動子DNA互作的調控模式[7]。TCP轉錄因子在調控植物生長發育和脅迫響應中的功能被廣泛鑒定,其結構和調控機理被相繼報道[8]。已經鑒定了許多植物中的TCP 基因;如在擬南芥中鑒定了24個TCP成員,在水稻中發現了26個TCP成員,在毛竹基因組中有19個TCP成員,紫花苜蓿中發現了40個TCP成員,這些基因大多數都定位于細胞核[9],作為轉錄因子主要在細胞核里面發揮調控功能。轉錄因子 TCP4 參與下胚軸延伸、次生細胞壁的形成、子葉的舒展、葉片形態和表皮毛分化等植物生長發育過程[10]。大豆的轉錄因子TCP4可以通過調節乙烯和脫落酸信號來響應干旱脅迫[11]。
關于棗樹 TCP轉錄因子的功能研究還鮮有報道[12]。選擇干旱敏感型棗樹品種冬棗和耐旱型棗樹品種贊皇大棗為實驗材料,通過small RNA-seq和降解組測序技術在棗樹全基因組水平挖掘干旱相關的miRNAs及其調控的靶基因,對棗樹干旱相關的miRNAs的鑒定得知:干旱脅迫和正常澆水相比較,novel-miR16在冬棗和贊皇大棗中都差異表達,說明棗樹novel-miR16響應干旱脅迫;RT-qPCR分析表明,novel-miR16在干旱脅迫下明顯差異表達[13]。對棗樹干旱相關的miRNA靶基因的鑒定中,驗證了TCP4為棗樹novel-miR16的靶基因,靶基因功能注釋和相關研究報道表明TCP類轉錄因子響應干旱脅迫。因此,選擇novel-miR16研究其在棗樹響應干旱脅迫過程中的功能。本研究從棗樹中研究發現novel-miR16的靶基因為TCP轉錄因子,并在棗樹中克隆到ZjTCP4基因的序列,對該基因進行生物信息學分析,亞細胞定位和干旱脅迫下的表達分析,以期為TCP轉錄因子響應干旱脅迫的機理研究提供參考。
1 材料與方法
1.1 試驗材料
本氏煙草無菌苗由陜西博瑞德生物科技有限公司提供。培養室溫度為26 ℃,光照1 500 lx,12 h光照、12 h黑暗。贊皇大棗棗樹苗由河北農業大學提供。載體及試劑:DH5α感受態細胞購自北京全式金生物技術有限公司,pMD?19-T Vector購自TaKaRa公司,pART-CAM-EGFP由陜西博瑞德生物科技有限公司提供,植物總RNA提取試劑盒購自天根生化科技(北京)有限公司,DNA 膠回收試劑盒、DL2000 Marker、DNA聚合酶和各類限制性內切酶均購自上海生物工程有限公司。
1.2 數據庫與分析軟件
已知植物miR319成熟體序列從miRNA數據庫 (http://www.mirbase.org)中獲得;棗樹novel-miR16靶基因利用在線預測軟件psRNATarget (http://plantgrn.noble.org/psRNATarget/)預測獲得,novel-miR16的靶基因序列在棗樹基因組數據庫(Ziziphus_jujuba.version2_13chr )中下載獲得;Expasy(http://www.expasy.org/tools)用于棗樹novel-miR16靶基因ZjTCP4編碼蛋白的基本性質分析;軟件UNAFold (http://www.bioinfo.rpi.edu/applications/mfold/old/rna/)預測novel-miR16前體序列的二級結構;利用DNAMAN(Lynnon Biosoft, San Ramon, CA, USA)構建系統進化樹。
1.3 棗樹novel-miR16序列和結構分析
將高通量測序獲得棗樹novel-miR16成熟體序列與miRNA數據庫miRBase中已知的植物miRNA成熟序列比對,分析其保守性。利用在線分析軟件UNAFold (http://www.bioinfo.rpi.edu/applications/mfold/old/rna/)預測novel-miR16前體序列的二級結構。
1.4 棗樹novel-miR16靶基因的預測
根據高通量測序獲得棗樹novel-miR16的成熟體序列;利用植物miRNA靶基因在線預測軟件psRNATarget預測棗樹novel-miR16的靶基因,通過對靶基因的功能注釋鑒定其功能。
1.5 棗樹novel-miR16靶基因ZjTCP4的驗證
為了驗證棗樹novel-miR16預測的靶基因,我們用RNA連接酶介導的RLM-5′RACE(Invitrogen公司的GeneRaeer 試劑盒)對棗樹novel-miR16預測靶基因的降解位點進行了驗證。采集棗樹葉片并提取總RNA,連接RNA接頭并反向轉錄獲得cDNA,根據 GeneRacer試劑盒5′接頭引物和靶基因ZjTCP4的特異性引物(Outer primer:5′-CAAGCCTCTTGCTTCTGATCC-3′和Inner primer:5′-CTGTGAATTTGTACAATTGCTTTC-TTAGCAACCTGTGAATTTGTACAATTGCTTTCTT-AGCAAC-3′)進行重疊PCR擴增獲得靶基因ZjTCP4的5′端序列,PCR擴增片段經凝膠電泳純化回收后克隆到pMD19-T進行測序,通過分析靶基因的降解位點來驗證預測棗樹novel-miR16靶基因。
1.6 棗樹TCP4基因的克隆
在棗樹基因組數據庫(Ziziphus_jujuba.version2_13chr)中下載獲得ZjTCP4(CCG006181)基因序列,利用軟件DNAMAN設計棗樹ZjTCP4 基因的PCR 克隆引物;正向引物:5′-CGGGATCCTAACATGGAC GCCATCAATC TC-3′,退火溫度67.5 ℃;反向引物:5′-CGAGCTCCAATCTCACAAACCACTCTCTTCTTC-3′,退火溫度67 ℃;并送生物工程有限公司。以提取到的棗樹葉片總 RNA 為模板,按AMVcDNA合成試劑盒合成cDNA,用ZjTCP4 基因特異的引物進行PCR 擴增,反應體系:TaqPCR Master Mix(上海生工,貨號:B639295)10 μL,ZjTCP4-F(10 μmol·μL-1) 1 μL, ZjTCP4-R(10 μmol·μL-1) 1 μL, 模板cDNA 1 μL,ddH2O 7 μL。將PCR產物與pMD?19-T Vector 載體進行連接,用熱激法將連接產物轉入大腸埃希菌 DH5α感受態細胞進行測序。
1.7 ZjTCP4的生物信息學分析
利用在線分析工具Expasy(http://web.expasy.org/protparam/)分析ZjTCP4基因編碼多肽的等電點、分子量及相關理化性質;Swiss-Model(http://swissmodel.expasy.org/)預測棗樹ZjTCP4基因編碼蛋白質三級結構模型;用TMHM-M-2.0(http://www.cbs.dtu.dk/services/TMHMM/)軟件進行ZjTCP4基因編碼蛋白跨膜結構域分析;利用SignalP 3.0 Server (http://www.cbs. dtu.dk/services/SignalP)信號肽預測軟件分析ZjTCP4基因編碼蛋白信號肽;利用NetPhos 2.0 Server軟件進行ZjTCP4基因編碼蛋白質的翻譯后加工修飾預測;利用軟件ProtScale(http://web.expasy.org/protscale/)分析ZjTCP4基因的親疏水性;利用Plant-mPLoc蛋白亞細胞定位預測軟件預測ZjTCP4基因的作用位置[14];通過軟件DNAMAN(Lynnon Biosoft, San Ramon, CA, USA)對靶基因ZjTCP4及擬南芥TCP轉錄因子構建系統進化樹。
1.8 ZjTCP4亞細胞定位
將靶基因ZjTCP4與亞細胞定位載體pART-CAM-EGFP連接構建表達載體,將本氏煙草植株置于白色熒光燈下一段時間使葉片氣孔打開,再用注射器將導入亞細胞定位載體的菌液注入煙草植株葉片的表皮,直到看到液體擴散,再侵染其他部位,侵染后,用記號筆圈定侵染區域。然后將葉片噴水,套上保鮮袋后置于黑暗環境培養過夜。培養48 h后打開保鮮袋,培養72 h后切取標記區域,撕下煙草葉片表皮在熒光顯微鏡下觀察拍照。
1.9 棗樹novel-miR16及靶基因ZjTCP4在干旱脅迫下的表達分析
通過qRT-PCR技術檢測棗樹novel-miR16及其靶基因ZjTCP4在贊皇大棗棗樹品種中的表達情況,贊皇大棗為定干80 cm 的3年生截干苗。2020年秋季將棗樹苗栽植在花盆中后,將花盆埋在地里面越冬。2021 年7月在甘肅農業大學的標準設施溫室進行盆栽的干旱試驗,干旱處理的土壤含水量為田間持水量的25%±5%,定期用土壤濕度計進行監測,用稱重法進行測定和澆水。分別提取干旱脅迫0、5、10、15、20、25 d時贊皇大棗棗樹的葉片總RNA,并用One Step PrimeScript1 miRNA cDNA synthesis kit(TaKaRa)試劑盒反轉錄獲得各自的cDNA,檢測棗樹novel-miR16及其靶基因ZjTCP4在干旱脅迫不同時期的表達情況,以ef1a基因為內參基因,PCR條件為:94 ℃預變性10 min;94 ℃變性 20 s,58 ℃ 退火30 s,設置35個循環。最后用2-ΔΔCT公式計算novel-miR16及其ZjTCP4基因在不同時間的相對表達量。
2 結果與分析
2.1 棗樹novel-miR16的二級結構分析
UNAFold 預測novel-miR16前體序列的二級結構表明,其二級結構具有miRNA典型的發夾結構(圖1),20 bp的novel-miR16成熟體序列(UUGGACU GAAGGGAGCUCCC)位于二級結構的5′端。

紅色部分表示novel-miR16成熟序列。Mature novel-miR16 sequences are in red.圖1 棗樹novel-miR16二級發夾結構Fig.1 The secondary hairpin structures of novel-miR16 in Chinese jujube
2.2 棗樹novel-miR16的保守性分析
為了研究棗樹novel-miR16的功能,我們分析了棗樹novel-miR16成熟體序列的保守性,結果表明,棗樹novel-miR16的成熟體序列與miRBase中植物miR319成熟體序列高度保守,與67個已知植物miR319成熟體序列完全相同,與其他33個已知植物miR319成熟體序列僅有1個堿基差異。因此,認為novel-miR16為棗樹miR319家族成員。
2.3 棗樹novel-miR16靶基因的預測
棗樹novel-miR16靶基因利用psRNATarget軟件預測獲得(表1)。通過對預測靶基因的序列比對和結構域分析表明,棗樹novel-miR16靶基因與TCP轉錄因子高度同源,且具有TCP轉錄因子特征結構域,屬于TCP轉錄因子家族,并將靶基因命名為ZjTCP4(CCG006181)。

表1 棗樹novel-miR16的靶基因預測Table 1 Predicted target genes of novel-miR16 in Chinese jujube
2.4 棗樹novel-miR16靶基因ZjTCP4的驗證
為了進一步驗證novel-miR16預測的靶基因ZjTCP4,我們利用RLM-5′RACE技術對novel-miR16在靶基因ZjTCP4上的切割位點進行驗證。靶基因ZjTCP4的降解位點多數都在novel-miR16結合位點的第10和11個核苷酸的位置 (圖2),表明ZjTCP4為棗樹novel-miR16的靶基因。
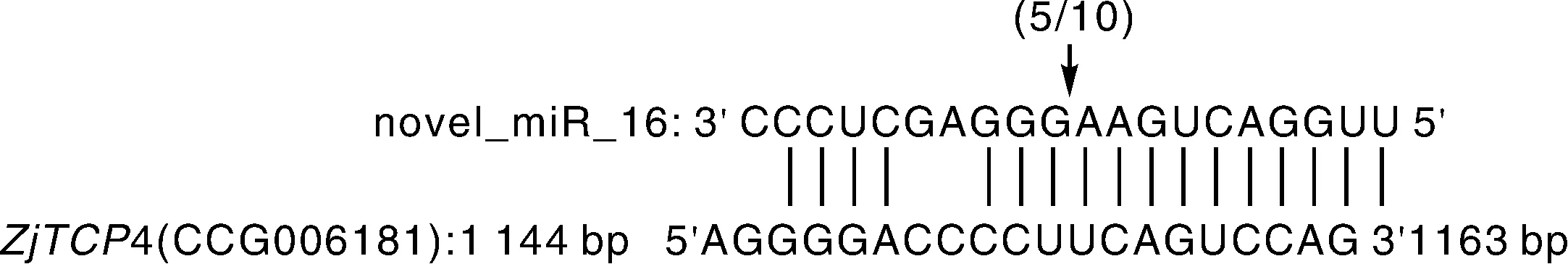
圖2 棗樹novel-miR16靶基因的驗證Fig.2 Verification of novel-miR16 target mRNA in Chinese jujube
2.5 棗樹ZjTCP4基因的克隆
以ZjTCP4-F和ZjTCP4-R 為引物,在棗樹中擴增ZjTCP4基因,擴增獲得1 386 bp大小的預期條帶(圖3),初步判定靶基因ZjTCP4(CCG006181)擴增成功,將PCR產物與克隆載體連接后送上海生物工程有限公司進行測序,測序結果表明,擴增序列與ZjTCP4(CCG006181)序列只有3個堿基的差異,說明該基因擴增成功。
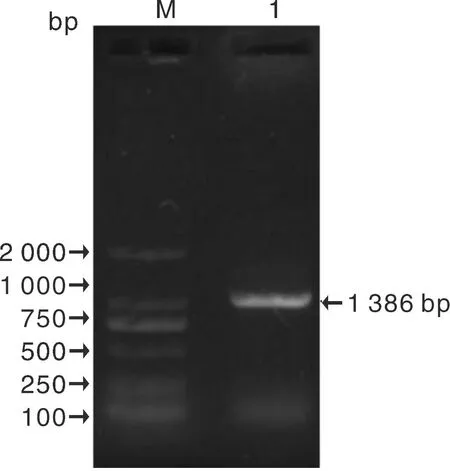
M,2 000 bp ladder Marker;1,PCR 產物。M, 2 000 bp ladder Marker; 1, PCR products.圖3 PCR產物電泳檢測Fig.3 Result of electrophoresis of PCR products
2.6 棗樹ZjTCP4基因的生物信息學分析
2.6.1 棗樹ZjTCP4基因編碼蛋白質理化性質分析
生物信息學分析表明,ZjTCP4基因的全長cDNA序列為1 386 bp,預測編碼452個氨基酸的多肽,預測編碼蛋白質的等電點(pI)為6.51,分子量為49 091.55 u。其分子量、等電點及其預測蛋白質的長度與已報道擬南芥、水稻和楊樹等模式植物TCP轉錄因子家族成員極為相似。
2.6.2 棗樹ZjTCP4基因編碼蛋白質三級結構
Swiss-Model預測棗樹ZjTCP4基因編碼蛋白質三級結構模型,結果表明,該基因主要有2個α螺旋和2個折疊,詳細結果見圖4。
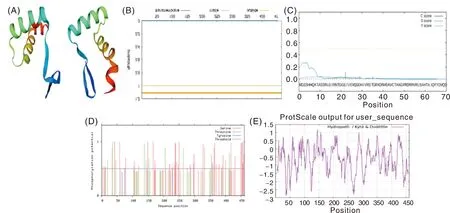
A,ZjTCP4基因編碼的蛋白質三級結構預測;B,ZjTCP4基因編碼的蛋白質跨膜結構域分析圖;C,ZjTCP4基因編碼的蛋白質信號肽預測;D,ZjTCP4基因編碼蛋白質翻譯后加工修飾預測;E,ZjTCP4基因編碼的蛋白質疏水性分析。A, Tertiary structure prediction of protein encoded by ZjTCP4 gene;B, Transmembrane domain analysis of protein encoded by ZjTCP4 gene; C, Signal peptide prediction of protein encoded by ZjTCP4 gene;D, Post-translational modification prediction of protein encoded by ZjTCP4 gene; E, Hydrophobicity analysis of protein encoded by ZjTCP4 gene.圖4 ZjTCP4基因編碼的蛋白質生物信息分析圖Fig.4 Bioinformatics analysis of protein encoded by ZjTCP4 gene
2.6.3 棗樹ZjTCP4基因的跨膜結構域分析
用TMHM-M-2.0軟件進行ZjTCP4基因編碼蛋白跨膜結構域分析,結果表明,該基因沒有跨膜結構域,應該是一個膜內或膜外蛋白,詳細見圖4。
2.6.4ZjTCP4基因的信號肽分析
利用SignalP 3.0 Server軟件預測ZjTCP4編碼蛋白的信號肽,結果表明,該基因編碼蛋白未發現信號肽(圖4),可能為非分泌性蛋白。
2.6.5 棗樹ZjTCP4基因編碼蛋白翻譯后加工修飾預測
對ZjTCP4翻譯后加工修飾的預測發現,ZjTCP4基因編碼蛋白有較高的磷酸化修飾,磷酸化修飾位點有76個,其中Ser磷酸化修飾位點 39個,Thr 磷酸化修飾位點17個(圖4)。
2.6.6 棗樹ZjTCP4基因編碼蛋白疏水性分析
ZjTCP4基因編碼蛋白親水性分析表明,該基因整體的親水性明顯強于疏水性,為親水性蛋白質(圖4)。
2.6.7 棗樹novel-miR16靶基因ZjTCP4的亞細胞定位預測
利用Plant-mPLoc蛋白亞細胞定位預測軟件分析ZjTCP4基因編碼蛋白在細胞內的作用位置,結果表明,該基因定位于細胞核,與預期結果一致,也符合轉錄因子的位置特征。
2.6.8 棗樹ZjTCP4基因的保守性分析
將棗樹ZjTCP4基因編碼蛋白與擬南芥33個TCP轉錄因子進行序列比對分析,并用DNAMAN(Lynnon Biosoft, San Ramon, CA, USA)構建系統進化樹。結果表明,棗樹ZjTCP4基因編碼的氨基酸序列與擬南芥AtTCP4最為相似,與擬南芥AtTCP17,AtTCP18和AtTCP12的相似性也較高,詳細結果見圖5。
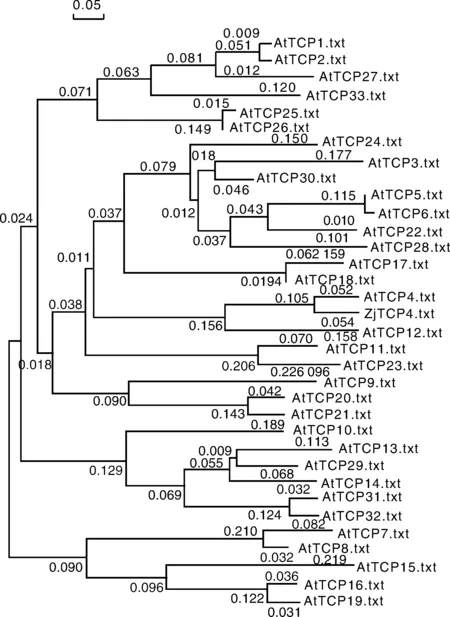
圖5 ZjTCP4基因編碼的蛋白質系統進化樹Fig.5 Phylogenetic tree of protein encoded by ZjTCP4
2.7 棗樹novel-miR16靶基因ZjTCP4的亞細胞定位驗證
將構建的pCAM-GFP-ZjTCP4載體和pCAM-GFP空載體(陰性對照)注射至煙草葉片,黑暗環境培養48 h后用激光共聚焦顯微鏡在488 nm波長處觀察拍照。結果顯示:空載體pCAM-GFP定位于細胞質、細胞核和細胞膜中,而pCAM-GFP-ZjTCP4融合蛋白主要定位于細胞核(圖6)。
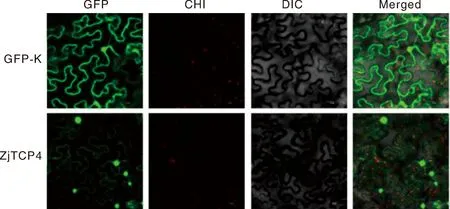
從左到右分別為 CK/基因的 GFP 綠色熒光蛋白、CHI、明場和3個通道的疊加照片。From left to right, superimposed photos of GFP green fluorescent protein, CHI, bright field and three channels of CK/gene, respectively.圖6 ZjTCP4基因的亞細胞定位圖Fig.6 Subcellular localization of the ZjTCP4 protein
2.8 棗樹novel-miR16及靶基因ZjTCP4響應干旱脅迫
為了檢測棗樹novel-miR16及靶基因ZjTCP4(CCG006181)是否響應干旱脅迫,本研究利用qRT-PCR分析了棗樹novel-miR16及靶基因ZjTCP4 (CCG006181)在干旱脅迫不同時期的表達模式。結果表明,棗樹novel-miR16及靶基因ZjTCP4(CCG006181)在響應干旱脅迫的過程中呈現相反的表達模式,棗樹novel-miR16在響應干旱脅迫的過程中上調表達,而其靶基因ZjTCP4(CCG006181)在干旱脅迫下下調表達(圖7)。
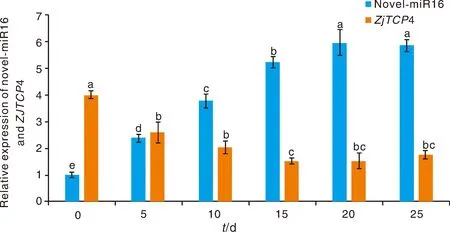
同一基因不同時間點間沒有相同小寫字母表示差異顯著(P<0.05)。No identical lowercase letters between different time points of the same gene indicate significant differences (P<0.05).圖7 棗樹novel-miR16及基因靶ZjTCP4在干旱脅迫下的表達分析Fig.7 The expression level of novel-miR16 and ZjTCP4 under drought stress in Chinese jujube
3 討論
3.1 novel-miR16是棗樹中保守的miR319家族成員
MiR319是植物中最為保守miRNA家族之一[15-16]。另外,miR319靶向TCP轉錄因子在植物中同樣保守[17-19]。已知miR319靶向TCP轉錄因子在多種植物發育和對各種脅迫的響應中發揮重要作用[17,20]。例如,在鹽脅迫下,miR319在擬南芥[16]、小麥[21]、蔓生彎草[19]和柳枝稷[22]等多種植物中都上調表達,而其靶基因TCP類轉錄因子下調表達。這說明miR319靶向TCP轉錄因子的調控模式在植物中也同樣保守。本研究在棗樹中鑒定出1個 miR319 家族新成員(novel-miR16),保守性分析表明,棗樹novel-miR16的成熟體序列與miRBase中植物miR319成熟體序列高度保守,與67個已知miR319成熟體序列完全相同,與其他33個已知miR319成熟體序列僅有1個堿基差異。表明novel-miR16是棗樹中保守的miR319家族成員之一。另外,靶基因預測表明,棗樹novel-miR16的靶基因為ZjTCP4(CCG006181),5′-RACE對靶基因ZjTCP4 (CCG006181)驗證表明,其切割位點多數呈現在novel-miR16結合位點的第10和11個核苷酸之間,這與擬南芥miR319對靶基因TCP4的切割位點一致,說明植物miR319與靶基因TCP類轉錄因子的剪切位點在不同物種間保守性較強。
3.2 棗樹novel-miR16靶向ZjTCP4基因響應干旱脅迫
眾所周知,miRNA通過調控其靶基因的表達來調節植物對干旱脅迫的響應,大量響應干旱脅迫的miRNA及其靶基因已在多種植物中被鑒定出來[23-26]。由于miRNA在轉錄后調控靶基因mRNA的降解,miRNA與靶基因呈現相反的表達模式,因此,通常通過分析miRNA與靶基因在脅迫下的表達模式來鑒定miRNA在脅迫響應中的功能。例如,擬南芥在磷脅迫下miR399的表達量上升,而靶基因泛素結合酶基因UBC24下調表達[24];水稻在冷脅迫下miR159的表達量會明顯降低,導致其靶基因MYB3R-2類轉錄因子表達量的升高而增強水稻的耐凍性[25];植物miR395的靶基因為APS基因,植物中miR395表達量的降低會導致其靶基因APS表達量的積累,最終會使植物體內谷胱甘肽的含量增加,提高植物對多種非生物脅迫的抗性[26]。
本研究通過高通量測序在棗樹中鑒定出1個miR319新成員novel-miR16,并通過生物信息學分析和實驗驗證鑒定出其靶基因ZjTCP4。研究發現,TCP 轉錄因子通過激活或抑制下游干旱相關基因的表達,在植物響應逆境脅迫中發揮著重要的調控作用[27-28]。也有報道表明,miR319與TCP存在靶向關系,響應著植物逆境脅迫過程。我們利用qRT-PCR檢查novel-miR16及靶基因ZjTCP4 (CCG006181)在干旱脅迫不同時期的表達模式發現,棗樹novel-miR16及靶基因ZjTCP4 (CCG006181)在響應干旱脅迫的過程中呈負相關,棗樹novel-miR16在響應干旱脅迫的過程中上調表達,而其靶基因ZjTCP4 (CCG006181)在干旱脅迫下下調表達。這與在模式植物中的研究結果基本一致[29-30]。研究發現,在擬南芥、水稻、白菜、棉花、蘭花的TCP基因中發現了miR319的靶基因位點,并且在脅迫條件下miR319與TCP基因的表達模式就是呈負相關關系。miR319主要是通過靶向轉錄因子,參與影響調控植物的激素合成、信號轉導、抗逆作用和生長發育。有研究表明:過表達水稻miR319可以提高水稻的耐鹽性[17],與本研究結果相似。因此,本研究表明,棗樹novel-miR16通過靶向調控ZjTCP4轉錄因子響應干旱脅迫,干旱脅迫下棗樹novel-miR16的上調表達導致靶基因ZjTCP4 (CCG006181)下調表達來提高棗樹的耐旱性。TCP4轉錄因子可能在ABA和JA激素介導的干旱脅迫響應中起正調控作用,這可以為深入解析棗樹novel-miR16靶向TCP4轉錄因子調控植物響應干旱脅迫的分子機理提供了理論基礎。
4 結論
本研究在棗樹中鑒定出miR319家族成員novel-miR16,并通過靶基因預測和實驗驗證鑒定出棗樹novel-miR16的靶基因為ZjTCP4(CCG006181)轉錄因子。通過分析棗樹novel-miR16及靶基因ZjTCP4轉錄因子在干旱脅迫下的表達模式,結果表明,在干旱脅迫下棗樹novel-miR16上調表達,而其靶基因ZjTCP4 (CCG006181)下調表達,表明novel-miR16及其靶基因ZjTCP4在棗樹對干旱脅迫的響應過程中發揮著重要的調控作用,但棗樹novel-miR16靶向ZjTCP4調控棗樹響應干旱脅迫的分子機制還需進一步研究。
猜你喜歡
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
山東工業技術(2016年15期)2016-12-01 05:31:22
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
