RasGRP1啟動子區(qū)甲基化對脂多糖誘導(dǎo)人T淋巴細(xì)胞炎癥的作用及機制*
2024-04-22 13:21:50劉磊劉美言高源1閔潔煜1殷永強鐘毅
貴州醫(yī)科大學(xué)學(xué)報 2024年3期
劉磊, 劉美言, 高源1,, 閔潔煜1,, 殷永強, 鐘毅
(1.貴州醫(yī)科大學(xué)附屬醫(yī)院 麻醉科, 貴州 貴陽 550001; 2.貴州醫(yī)科大學(xué) 麻醉學(xué)院, 貴州 貴陽 550004)
膿毒癥是宿主對感染的反應(yīng)失調(diào)引起的危及生命的器官功能障礙,嚴(yán)重時可發(fā)展為膿毒癥休克,甚至死亡[1]。DNA甲基化是一種重要的表觀遺傳學(xué)修飾,已有研究表明在膿毒癥診療中某些基因的甲基化修飾對患者疾病轉(zhuǎn)歸具有重要作用[2]。在膿毒癥早期炎癥階段,體內(nèi)數(shù)千個基因的表達(dá)就已經(jīng)發(fā)生了改變,涉及數(shù)百個基因的DNA甲基化水平也發(fā)生了改變[3-4],這些基因大多與細(xì)胞炎癥反應(yīng)和干擾素(interferon,IFN)信號轉(zhuǎn)導(dǎo)有關(guān),同時還能夠決定T細(xì)胞的分化方向[5-6]。Hopp等[7]在對社區(qū)獲得性肺炎患者全血樣本的分析中發(fā)現(xiàn),DNA甲基化在膿毒癥的急性炎癥中起重要作用;新生兒膿毒癥患者中炎癥相關(guān)基因的甲基化水平有顯著改變,某些基因的DNA甲基化可作為生物標(biāo)志物區(qū)分早期和晚期膿毒癥[8]。采用生物信息學(xué)分析篩選膿毒癥診斷和預(yù)后生物標(biāo)志物[9],并結(jié)合本課題組前期研究,RAS鳥苷酸釋放蛋白1(ras guanyl nucleotide releasing protein 1,RasGRP1)在膿毒癥患者中存在表達(dá)差異且甲基化程度高。因此,本研究旨在通過由細(xì)菌脂多糖(lipopolysaccharide,LPS)誘導(dǎo)淋巴細(xì)胞炎癥模型,使用甲基化轉(zhuǎn)移酶抑制劑5-Aza-2′-脫氧胞苷(5-Aza-2′-deoxycytidine)與LPS共處理,驗證RasGRP1及其甲基化修飾在LPS誘導(dǎo)淋巴細(xì)胞炎癥模型中的作用。
1 材料與方法
1.1 實驗材料
1.1.1細(xì)胞株來源 人T淋巴細(xì)胞白血病細(xì)胞JurkatE6-1(CL-0129),由武漢普諾賽生命科技有限公司提供,由學(xué)校轉(zhuǎn)化醫(yī)學(xué)研究中心保存。
1.1.2主要試劑與儀器 JurkatE6-1細(xì)胞專用培養(yǎng)基洛斯維·帕克紀(jì)念研究所(Roswell Park Memorial Institute,RPMI)1640培養(yǎng)基,由武漢普諾賽生命科技有限公司提供;甲基化轉(zhuǎn)移酶抑制劑5-Aza-2′-脫氧胞苷(美國MedChemExpress公司),脂多糖(北京索萊寶科技有限公司),細(xì)胞總RNA、DNA提取試劑盒(廣州美基生物科技有限公司),二喹啉甲酸(bicinchoninicacid,BCA)蛋白定量試劑盒(上海碧云天生物技術(shù)有限公司),DNA重亞硫酸鹽轉(zhuǎn)化試劑盒(北京天根生化科技有限公司),RasGRP1一抗、細(xì)胞外調(diào)節(jié)蛋白激酶(extracellular regulated protein kinases,ERK1/2)一抗、磷酸化細(xì)胞外調(diào)節(jié)蛋白激酶(phosphorylated extracellular regulated protein kinases,p-ERK1/2)一抗(武漢ABclonal公司),3-磷酸甘油醛脫氫酶(glyceraldehyde-3-phosphate dehydrogenase,GAPDH)一抗(武漢三鷹生物技術(shù)有限公司),山羊抗兔二抗(英國Abcam公司),實時熒光定量PCR(real-time fluorescent quantitative PCR,RT-qPCR)第一鏈互補DNA(complementary DNA,cDNA)合成試劑盒、熒光定量試劑盒(上海翌圣生物科技股份有限公司),酶聯(lián)免疫吸附實驗(enzyme linked immunosorbent assay,ELISA)試劑盒(武漢伊萊瑞特生物科技股份有限公司),乳酸脫氫酶(lactate dehydrogenase,LDH)測定試劑盒(南京建成生物工程有限公司);酶標(biāo)儀(美國Thermo Fisher 公司),CFX96實時熒光PCR儀(美國Bio-Rad公司),奧林巴斯激光共聚焦顯微鏡(日本Olympus公司)。
1.2 研究方法
1.2.1細(xì)胞培養(yǎng)和分組 JurkatE6-1細(xì)胞復(fù)蘇后使用RPMI-1640培養(yǎng)基于5% CO2、37 ℃的培養(yǎng)箱培養(yǎng);取生長狀態(tài)良好的細(xì)胞計數(shù)后接種于培養(yǎng)皿中,細(xì)胞培養(yǎng)箱中培養(yǎng)24 h,使細(xì)胞同步化;當(dāng)細(xì)胞生長至融合度達(dá)80%左右時傳代并進(jìn)行后續(xù)實驗;確認(rèn)細(xì)胞生長狀態(tài)良好后,分為對照(完全培養(yǎng)基,C)組、1 mg/L(低濃度)LPS(L1)組、10 mg/L(高濃度)LPS(L2)組及10 μmol/L甲基化轉(zhuǎn)移酶抑制劑5-Aza-2′-脫氧胞苷+10 mg/L LPS(LA)組。LA組細(xì)胞給予10 μmol/L 5-Aza-2′-脫氧胞苷處理4 d,期間每12 h更換含相同濃度5-Aza-2′-脫氧胞苷的完全培養(yǎng)基;C組、L1組及L2組等時等量完全培養(yǎng)基換液4 d。持續(xù)4 d,L1組細(xì)胞予1 mg/L LPS處理24 h,L2組和LA組細(xì)胞給予10 mg/L LPS處理24 h,C組細(xì)胞繼續(xù)用完全培養(yǎng)基培養(yǎng)24 h。各組細(xì)胞繼續(xù)培養(yǎng)24 h,1 200 r/min離心3 min,收集細(xì)胞、取上清夜,保存于-80 ℃冰箱。
1.2.2熒光顯微鏡觀察細(xì)胞形態(tài)學(xué)變化 取“1.2.1”項下處于對數(shù)生長期的JurkatE6-1細(xì)胞(5×106個/孔)接種于六孔板,分組及處理同“1.2.1”,置于熒光顯微鏡下觀察細(xì)胞形態(tài)學(xué)變化。
1.2.3ELISA法檢測細(xì)胞上清液炎癥因子濃度和比色法檢測LDH活性 取“1.2.1”項下處于對數(shù)生長期的JurkatE6-1細(xì)胞(5×106個/孔)接種于六孔板,分組及處理同“1.2.1”,參照ELISA試劑盒,使用酶標(biāo)儀檢測各孔450 nm處的吸光度值,繪制標(biāo)準(zhǔn)曲線,計算腫瘤壞死因子-α(tumor necrosis factor-α,TNF-α)、白細(xì)胞介素-6(interleukin-6,IL-6)濃度;參照LDH測定試劑盒,使用酶標(biāo)儀測定450 nm處吸光度值,繪制標(biāo)準(zhǔn)曲線,計算LDH活性。
1.2.4RT-qPCR法檢測細(xì)胞中總RNA的表達(dá) 取“1.2.1”項下處于對數(shù)生長期的JurkatE6-1細(xì)胞(5×106個/孔)接種于6孔板,分組及處理同“1.2.1”,參照細(xì)胞總RNA提取試劑盒,按照說明書提取細(xì)胞總RNA,測定總RNA濃度和純度。按照說明書逆轉(zhuǎn)錄合成第一鏈cDNA;使用RT-qPCR試劑盒,按說明書進(jìn)實驗,擴增條件為95 ℃預(yù)變性2 min,95 ℃變性10 s、60 ℃退火延伸30 s、共40個循環(huán);以GAPDH為內(nèi)參,以2-ΔΔCt法計算RasGRP1、ERK1/2、TNF-α、IL-6、DNA甲基轉(zhuǎn)移酶(DNA methyl trans-ferases,DNMT)1、DNMT3a基因mRNA的相對表達(dá)量。引物序列見表1。

表1 RT-qPCR引物序列Tab.1 Primer sequences of RT-qPCR
1.2.5Western blotting法檢測細(xì)胞RasGRP1、ERK1/2及p-ERK1/2蛋白的表達(dá) 取“1.2.1”項下處于對數(shù)生長期的JurkatE6-1細(xì)胞(5×106個/孔)接種于六孔板,分組及處理同“1.2.1”,加適量細(xì)胞裂解液,充分裂解后4 ℃、12 000 r/min離心10 min,收集上清液,使用BCA蛋白定量試劑盒檢測蛋白質(zhì)濃度,定量后蛋白質(zhì)經(jīng)95 ℃煮沸10 min變性,-80 ℃冰箱中備用;取待測樣本50 μg于10%聚丙烯酰胺凝膠進(jìn)行電泳,結(jié)束后將分離的蛋白轉(zhuǎn)印至聚偏二氟乙烯膜(polyvinylidene fluoride,PVDF);高效封閉液封閉30 min,加一抗RasGRP1(1∶1 000)、ERK1/2(1∶1 500)、p-ERK1/2(1∶1 000)及GAPDH(1∶2 000),4 ℃孵育過夜;第2天洗膜后加入辣根過氧化物酶標(biāo)記的山羊抗兔二抗(1∶ 6 000)孵育,室溫下輕搖1.5 h,洗膜后于化學(xué)發(fā)光法顯色,用紅外成像儀對膜進(jìn)行掃描;以GAPDH為內(nèi)參,ImageJ軟件進(jìn)行蛋白灰度值相對定量分析。
1.2.6甲基化特異性PCR(methylating-specific PCR,MSP)法和瓊脂糖凝膠電泳檢測RasGRP1啟動子區(qū)甲基化狀態(tài) 取“1.2.1”項下處于對數(shù)生長期的JurkatE6-1細(xì)胞(5×106個/孔)接種于6孔板,分組及處理同“1.2.1”,使用DNA提取試劑盒提取細(xì)胞DNA,使用DNA重亞硫酸鹽轉(zhuǎn)化試劑盒將DNA樣品中非甲基化胞嘧啶轉(zhuǎn)化為尿嘧啶,而甲基化胞嘧啶保持不變,因此使用Methyl Primer Express v1.0軟件設(shè)計出甲基化引物和非甲基化引物。引物序列如下:RasGRP1非甲基化引物(unmethylation primer,UM)的上游引物序列為5-ATTGTAGTGTTTTGAGTAGTGGTT-3,下游引物序列為5-AAAACAAACTCCCAACTACCA-3;RasGRP1甲基化引物(methylation primer ,M)的上游引物序列為5-GTAGTGTTTCGAGTAGCGGTC-3,下游引物序列為5-AACGAACTCCCGACTACC-3。將經(jīng)過DNA重亞硫酸鹽轉(zhuǎn)化試劑盒處理后的DNA樣本按照設(shè)計好的引物序列進(jìn)行甲基化特異性PCR,擴增條件:95 ℃ 5 min預(yù)變性,94 ℃ 20 s、60 ℃ 30 s、72 ℃ 20 s,共35個循環(huán),再以72 ℃擴增5 min。將擴增產(chǎn)物經(jīng)160 V瓊脂糖凝膠電泳25 min,用凝膠成像分析系統(tǒng)對凝膠進(jìn)行拍照。
1.3 統(tǒng)計學(xué)分析
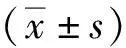
2 結(jié)果
2.1 細(xì)胞形態(tài)學(xué)
各組細(xì)胞相應(yīng)處理5 d后,光學(xué)顯微鏡結(jié)果顯示(圖1),C組JurkatE6-1細(xì)胞生長狀態(tài)良好;L1組、L2組使用LPS處理24 h后細(xì)胞數(shù)量明顯減少,細(xì)胞形態(tài)趨于碎片化,死亡細(xì)胞逐漸增加,隨LPS濃度增加而增加;LA組使用甲基化轉(zhuǎn)移酶抑制劑5-Aza-2′-脫氧胞苷預(yù)處理4 d、LPS處理細(xì)胞24 h后,較L2組細(xì)胞數(shù)量及形態(tài)明顯好轉(zhuǎn)。

圖1 各組JurkatE6-1細(xì)胞相應(yīng)處理5 d后明視夜野下細(xì)胞數(shù)量及形態(tài)(20×)Fig.1 The number and morphology of JurkatE6-1 cells in each group after 5 days of treatment under bright light field (20×)
2.2 細(xì)胞炎癥損傷水平
LPS處理細(xì)胞24 h后,ELISA和RT-qPCR結(jié)果顯示(表2),與C組比較,L1組細(xì)胞上清液LDH活性和TNF-α、IL-6濃度及其mRNA表達(dá)升高,差異均有統(tǒng)計學(xué)意義(P<0.05);與L1組比較,L2組細(xì)胞上清液LDH活性和TNF-α、IL-6濃度及其mRNA表達(dá)升高,差異均有統(tǒng)計學(xué)意義(P<0.05);與L2組比較,LA組細(xì)胞上清液LDH活性和TNF-α、IL-6濃度及mRNA表達(dá)降低,差異均有統(tǒng)計學(xué)意義(P<0.05)。
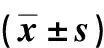
表2 各組細(xì)胞LDH活性和TNF-α、IL-6濃度及mRNA表達(dá)Tab.2 LDH activity, the concentrations and mRNA expression levels of TNF-α and IL-6 in each
2.3 細(xì)胞RasGRP1和ERK1/2 mRNA的表達(dá)
LPS處理細(xì)胞24 h,RT-qPCR結(jié)果顯示(表3),與C組比較,L1組細(xì)胞RasGRP1 mRNA表達(dá)升高,差異有統(tǒng)計學(xué)意義(P<0.05);與L1組比較,L2組細(xì)胞RasGRP1 mRNA表達(dá)降低,差異有統(tǒng)計學(xué)意義(P<0.05);與L2組比較,LA組細(xì)胞RasGRP1 mRNA表達(dá)升高,差異有統(tǒng)計學(xué)意義(P<0.05);各組間ERK1/2 mRNA表達(dá)比較,差異無統(tǒng)計學(xué)意義(P>0.05)。
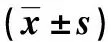
表3 各組細(xì)胞RasGRP1和ERK1/2 mRNA的表達(dá)Tab.3 mRNA expression levels of RasGRP1 and ERK1/2 in each
2.4 細(xì)胞RasGRP1、ERK1/2及p-ERK1/2蛋白的表達(dá)
LPS處理細(xì)胞24 h,Western blotting結(jié)果顯示(圖2),與C組比較,L1組細(xì)胞RasGRP1、p-ERK1/2蛋白表達(dá)升高(P<0.05),差異均有統(tǒng)計學(xué)意義(P<0.05);與L1組比較,L2組細(xì)胞RasGRP1、p-ERK1/2蛋白表達(dá)降低(P<0.05),差異均有統(tǒng)計學(xué)意義(P<0.05);與L2組比較,LA組細(xì)胞RasGRP1、p-ERK1/2蛋白表達(dá)升高(P<0.05),差異均有統(tǒng)計學(xué)意義(P<0.05);各組間ERK1/2 蛋白總量比較,差異無統(tǒng)計學(xué)意義(P>0.05)。
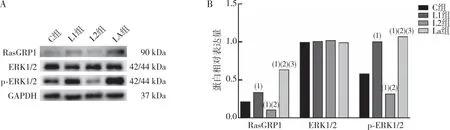
注:A為各組細(xì)胞蛋白表達(dá)的Western blotting檢測結(jié)果;B:各組細(xì)胞蛋白表達(dá)的定量結(jié)果;(1)與C組比較,P<0.05;(2)與L1組比較,P<0.05;(3)與L2組比較,P<0.05。
2.5 細(xì)胞DNMT1和DNMT3a mRNA的表達(dá)
甲基化轉(zhuǎn)移酶抑制劑5-Aza-2′-脫氧胞苷預(yù)處理4 d,LPS處理細(xì)胞24 h后,RT-qPCR結(jié)果顯示(表4),與C組比較,L1組DNMT1、DNMT3a mRNA表達(dá)比較,差異無統(tǒng)計學(xué)意義(P>0.05);與L1組比較,L2組DNMT1、DNMT3a mRNA表達(dá)升高,差異均有統(tǒng)計學(xué)意義(P<0.05);與L2組比較,LA組DNMT1、DNMT3a mRNA表達(dá)降低,差異均有統(tǒng)計學(xué)意義(P<0.05)。
表4 各組細(xì)胞DNMT1和DNMT3a mRNA的表達(dá)Tab.4 mRNA expressions of DNMT1 and DNMT3a
2.6 RasGRP1啟動子區(qū)甲基化狀態(tài)
甲基化轉(zhuǎn)移酶抑制劑5-Aza-2′-脫氧胞苷預(yù)處理4 d,LPS處理細(xì)胞24 h后,MSP及瓊脂糖凝膠電泳結(jié)果顯示(圖3),與C組比較,L1組RasGRP1啟動子區(qū)未檢測到甲基化改變;與L1組比較,L2組RasGRP1啟動子區(qū)呈現(xiàn)高甲基化表達(dá);與L2組比較,LA組RasGRP1啟動子區(qū)未檢測到甲基化。

注:U為非甲基化引物擴增產(chǎn)物,M為甲基化引物擴增產(chǎn)物。
3 討論
膿毒癥是一種病理生理機制難以闡明的臨床綜合征,現(xiàn)免疫抑制已被公認(rèn)是膿毒癥患者重要死亡原因之一[10]。近年來,膿毒癥導(dǎo)致的免疫抑制可能比最初的高炎癥反應(yīng)更能導(dǎo)致死亡率上升,免疫抑制程度較高的基因與死亡率增加相關(guān)[11]。在膿毒癥的發(fā)展過程中,淋巴細(xì)胞等免疫細(xì)胞凋亡可導(dǎo)致免疫功能紊亂,導(dǎo)致對繼發(fā)感染的敏感性增加,促炎反應(yīng)和抗炎反應(yīng)失衡,嚴(yán)重時可發(fā)展為多器官功能障礙,甚至引起死亡[12-13]。與本研究結(jié)果一致,使用LPS處理引起淋巴細(xì)胞炎癥損傷,細(xì)胞數(shù)量減少,碎片化細(xì)胞增多,且隨濃度升高而加重;因此,本研究使用不同濃度LPS誘導(dǎo)JurkatE6-1細(xì)胞炎癥模型,探究體外膿毒癥可能的發(fā)病機制。
RasGRP1又稱RAS鳥苷酸釋放蛋白1,是鳥嘌呤核苷酸交換蛋白家族成員之一,主要在T淋巴細(xì)胞中表達(dá),通過介導(dǎo)抗原啟動的信號通路激活淋巴細(xì)胞中的Ras-Raf-MEK-ERK通路,調(diào)控T細(xì)胞激活和發(fā)育[14]。RasGRP1表達(dá)異常的患者會出現(xiàn)淋巴細(xì)胞功能障礙,而適應(yīng)性免疫功能受損增加了對各種病原體反復(fù)、早發(fā)和嚴(yán)重感染的易感性[15]。研究發(fā)現(xiàn),核受體相關(guān)轉(zhuǎn)錄因子1(nuclear receptor related 1,Nurr1)通過與RasGRP1基因第二內(nèi)含子中特定位置結(jié)合來調(diào)節(jié)LPS誘導(dǎo)的炎癥信號級聯(lián)反應(yīng),在轉(zhuǎn)錄水平負(fù)向調(diào)節(jié)RasGRP1的表達(dá),在神經(jīng)炎癥中起到抗炎介質(zhì)的作用[16];在自身免疫的患者中,活動性炎癥的嚴(yán)重程度與CD4+T細(xì)胞中RasGRP1水平降低相關(guān)[17]。另外,Guo等[18]在RasGRP1基因敲除小鼠行盲腸結(jié)扎穿刺的膿毒癥模型研究中發(fā)現(xiàn),RasGRP1敲除小鼠的IgM抗體分泌和IL-10產(chǎn)生的障礙,且10 d存活率明顯降低,表明RasGRP1基因缺陷增加了小鼠患膿毒癥的敏感性;以上研究表明,RasGRP1與膿毒癥炎癥反應(yīng)密切相關(guān)。與本研究結(jié)果一致,JurkatE6-1細(xì)胞在低濃度LPS處理后RasGRP1的表達(dá)升高;高濃度LPS處理后,RasGRP1表達(dá)下降,DNMT1、DNMT3a mRNA的表達(dá)升高。不同濃度LPS誘導(dǎo)后RasGRP1表達(dá)趨勢相反,可能是低濃度LPS刺激淋巴細(xì)胞保護(hù)性應(yīng)激反應(yīng)使其表達(dá)升高,而高濃度LPS刺激細(xì)胞導(dǎo)致嚴(yán)重的炎癥反應(yīng),并可能誘導(dǎo)了RasGRP1啟動子區(qū)高甲基化,基因表達(dá)水平受到抑制;為了進(jìn)一步驗證RasGRP1啟動子區(qū)甲基化改變其及在炎癥反應(yīng)中調(diào)控作用,本研究使用甲基化轉(zhuǎn)移酶抑制劑5-Aza-2′-脫氧胞苷和LPS共處理JurkatE6-1細(xì)胞,使用DNA重亞硫酸鹽轉(zhuǎn)化法,將DNA樣品中非甲基化胞嘧啶轉(zhuǎn)化為尿嘧啶,而甲基化胞嘧啶保持不變,使用Methyl Primer Express v1.0軟件設(shè)計出甲基化引物和非甲基化引物,將硫化后的DNA進(jìn)行甲基化特異性PCR擴增,并將產(chǎn)物進(jìn)行瓊脂糖凝膠電泳檢測RasGRP1啟動子區(qū)甲基化狀態(tài)。
DNA甲基化是指真核生物中DNA甲基轉(zhuǎn)移酶類將甲基供體S-腺苷甲硫氨酸中的甲基轉(zhuǎn)移到DNA序列中CpG島的胞嘧啶上的過程[19]。在人單核細(xì)胞系中,脂多糖刺激導(dǎo)致腫瘤壞死因子啟動子的低甲基化,提高腫瘤壞死因子轉(zhuǎn)錄水平,促進(jìn)了炎癥反應(yīng)[20-21]。Shih等[22]DNMT1抑制劑普魯卡因胺治療內(nèi)毒素休克大鼠的研究顯示,抑制抗炎基因IL27RA的DNA甲基化,對循環(huán)衰竭和多器官功能障礙具有保護(hù)作用;Huang等[23]研究表明,DNMT抑制劑5-Aza-2′-脫氧胞苷可抑制膿毒癥性急性肺損傷(acute lung injury,ALI)小鼠炎癥反應(yīng)和氧化應(yīng)激;Rump等[24]研究顯示,水通道蛋白5(aquaporin 5,AQP5)基因nt-937胞嘧啶位點可與NF-κB結(jié)合有關(guān),膿毒癥死亡患者該位點上的甲基化水平明顯高于膿毒癥幸存者。根據(jù)體內(nèi)外研究和現(xiàn)有人類膿毒癥表觀遺傳學(xué)研究的數(shù)據(jù)證明,表觀遺傳學(xué)修飾可能是膿毒癥發(fā)病機制的核心,DNA甲基化可能是治療膿毒癥和改善器官衰竭的預(yù)后靶點。本研究參照文獻(xiàn)[25]方法,使用5-Aza-2′-脫氧胞苷與高濃度LPS同時處理JurkatE6-1細(xì)胞,使用MSP法和瓊脂糖凝膠電泳檢測RasGRP1啟動子區(qū)甲基化表達(dá)情況,結(jié)果顯示,高濃度LPS刺激誘導(dǎo)了細(xì)胞RasGRP1啟動子區(qū)高甲基化,甲基化轉(zhuǎn)移酶抑制劑預(yù)處理逆轉(zhuǎn)了RasGRP1啟動子區(qū)高甲基化,因此RasGRP1及其mRNA表達(dá)上調(diào);而細(xì)胞培養(yǎng)上清液炎癥因子TNF-α、IL-6濃度以及LDH活性降低,提示細(xì)胞炎癥損傷有所減輕。另外,本研究結(jié)果表明,細(xì)胞p-ERK1/2蛋白表達(dá)也隨之上調(diào),ERK1/2總蛋白不受影響,表明RasGRP1可能通過激活MAPK通路,調(diào)節(jié)下游蛋白ERK1/2磷酸化發(fā)揮作用。以上研究結(jié)果表明,高濃度LPS刺激細(xì)胞的嚴(yán)重炎癥反應(yīng)誘導(dǎo)了RasGRP1啟動子區(qū)高甲基化改變,通過抑制其啟動子區(qū)甲基化,上調(diào)RasGRP1的表達(dá)水平,可以減輕細(xì)胞炎癥反應(yīng),這可能與激活MAPK通路促進(jìn)ERK1/2磷酸化,調(diào)節(jié)淋巴細(xì)胞免疫應(yīng)答有關(guān),其具體調(diào)節(jié)機制還需深入研究。
綜上所述,本研究結(jié)果揭示RasGRP1表達(dá)及其啟動子區(qū)甲基化在LPS誘導(dǎo)的JurkatE6-1細(xì)胞炎癥模型中可能有調(diào)控作用,甲基化修飾療法可減輕細(xì)胞炎癥損傷,并可能通過調(diào)控下游ERK信號通路發(fā)揮作用。
