β-谷甾醇對豬卵母細胞體外成熟和胚胎發育的影響
2024-05-07 13:52:30藍昕蕊趙寶寶張碧菡林曉語馬會明王勇勝
畜牧獸醫學報 2024年4期
藍昕蕊,趙寶寶,張碧菡,林曉語,馬會明,王勇勝*
(1.西北農林科技大學動物醫學院,楊凌 712100;2.寧夏醫科大學 生育力保持教育部重點實驗室,銀川 750004)
卵母細胞體外成熟(invitromaturation,IVM)的關鍵在于用體外成熟培養基模擬卵泡液微環境,使未成熟的卵母細胞同時達到細胞核和質成熟。但是,卵母細胞體外成熟效率普遍低于體內成熟[1-3],氧化應激是一大重要因素。卵母細胞氧化應激和抗氧化能力在體內能夠達到平衡狀態,但在IVM過程中,線粒體電子傳遞鏈中產生的活性氧(ROS)過度積累,氧化和抗氧化功能紊亂,阻礙卵母細胞的成熟[4],另外還會造成線粒體功能障礙、紡錘體異常、細胞凋亡和發育受損,從而損傷卵母細胞,進一步導致卵母細胞成熟受阻[5-7]。
目前,已有多種抗氧化劑及其他類型的物質被添加到體外成熟培養基中,如白藜蘆醇、褪黑素和亞硒酸鈉[5,8-12]等,但因其可能存在細胞毒性作用、成本較高而并不被廣泛用于卵母細胞體外成熟培養。研究表明,藥用植物中的抗氧化劑能夠對多種疾病發揮作用,具有出色的抗氧化能力。Imai等[13]及Zhou等[14]曾從枸杞葉和枸杞根中分離出β-谷甾醇(β-sitosterol, SITO),它是一種常見的植物甾醇,具有廣泛的生物學效應,能夠治療多種慢性疾病,如氧化應激、肥胖、糖尿病和癌癥等[15-20],SITO可以恢復谷胱甘肽/氧化谷胱甘肽比例的損傷,這表明它可能是ROS清除劑[21]。SITO的抗氧化能力提示其可能減少卵母細胞體外培養過程中的氧化應激,可能有助于促進卵母細胞體外成熟。
目前,尚無任何研究表明SITO對卵母細胞體外成熟和早期胚胎發育有影響。因此,本研究評估了添加SITO能否改善豬卵母細胞IVM效率和早期胚胎發育質量,對提高卵母細胞體外成熟率及胚胎發育具有重要的理論意義和實用價值。
在預習的過程中,要善于發現問題,對于發現的問題,首先要進行獨立思考,看看是否能夠解決掉。如果解決掉了,很好;如果解決不了,就需要記錄下來,這樣學生就可以在課堂上有針對性地聽教師講解與這個問題相關的部分,這樣聽課的目的性就很強了,從而有利于學生提高聽課效率。
1 材料與方法
1.1 主要試劑
除特別說明,本試驗所有試劑均購自Sigma公司。活性氧檢測試劑盒、細胞凋亡檢測試劑盒、線粒體膜電位檢測試劑盒購自碧云天生物技術有限公司;反轉錄試劑盒、熒光定量試劑盒購自TaKaRa公司;引物由上海生工生物工程有限公司合成。
1.2 試驗方法
1.2.1 豬卵母細胞的收集和體外培養 從屠宰場獲得豬卵巢,在雙抗生理鹽水37 ℃、2 h內運至實驗室。注射器抽取卵泡直徑3~8 mm的卵泡液,在體視顯微鏡下挑揀出卵泡液中的卵丘-卵母細胞復合體(cumulus-oocyte complexes, COCs),選擇有至少3層致密顆粒細胞且胞質均勻的COCs進行培養。培養條件為5% CO2、38.5 ℃恒溫培養箱。處理組培養基分別添加10、20、40 μmol·L-1SITO(將1 g SITO粉末溶于0.5 mL DMSO+29.64 mL培養基中,制得80 mmol·L-1母液,再分別稀釋至10、20、40 μmol·L-1),含0.16% DMSO的培養基為對照組。培養46 h后,將COCs置于0.1%透明質酸酶中,反復吹打脫去顆粒細胞,以排出第一極體作為成熟的標志,統計成熟率(成熟率=排出第一極體卵母細胞數/培養卵母細胞總數×100)。
與對照組相比,處理組卵母細胞成熟率顯著增加(P<0.05,圖1A),10與40 μmol·L-1組之間差異不顯著(P>0.05)(對照組:(65.28±3.13)%,n=183;10 μmol·L-1SITO:(73.1±1.93)%,n=174;20 μmol·L-1SITO:(78.89±4.61)%,n=169;40 μmol·L-1SITO:(74.55±4.66)%,n=150)。各處理組與對照組卵裂率差異不顯著(P>0.05)(圖1B),但囊胚率均顯著增加(P<0.05,圖1C);(對照組:(25.60±2.99)%,n=25;10 μmol·L-1SITO:(32.95±1.55)%,n=33;20 μmol·L-1SITO:(43.39±2.49)%,n=43;40 μmol·L-1SITO:(37.08±2.17)%,n=32)。結果表明,在IVM培養基中添加SITO可以促進豬卵母細胞體外成熟和囊胚發育能力,20 μmol·L-1的作用最為顯著。
1.2.6 囊胚免疫熒光染色 收集發育至第7天的囊胚,用0.1% PBS-PVA洗滌3次,使用500 μL DAPI染色5 min,再洗滌3次后封片,在熒光顯微鏡下觀察。
1.2.2 孤雌激活和體外胚胎培養 在5 mmol·L-1ion中孵育4 min激活成熟卵母細胞,再在2 mmol·L-16-DMAP中培養3~5 h,然后在PZM-3中進行培養,培養條件為5% CO2+5% O2、38.5 ℃恒溫培養箱。將孤雌激活當天記為第0天,于第2天統計卵裂率,第7天統計囊胚率(卵裂率=卵裂胚胎數/排出第一極體卵母細胞數×100,囊胚率=囊胚數/卵裂胚胎數×100)。
1.2.3 活性氧含量檢測 根據DCFH-DA ROS檢測試劑盒說明書,將成熟卵母細胞用0.1% PBS-PVA洗滌3次,置于10 mmol·L-1DCFH-DA染色液中,37 ℃避光染色20 min。染色完成后再洗滌3次,封片,倒置熒光顯微鏡下拍照觀察。
如圖3所示,碘化丙啶(PI)為紅色熒光,Annexin V-FITC為綠色熒光,20 μmol·L-1SITO處理的卵母細胞紅色熒光強度顯著高于對照組(P<0.05),綠色熒光強度顯著低于對照組(P<0.05),說明SITO處理使豬卵母細胞凋亡顯著減少。如圖4所示,在線粒體膜電位較高時產生紅色熒光,在線粒體膜電位較低時產生綠色熒光,紅/綠熒光比則說明了ΔΨm的水平,20 μmol·L-1SITO處理卵母細胞的紅/綠熒光強度比高于對照組(P<0.05),表明20 μmol·L-1SITO具有增強線粒體膜電位,改善線粒體功能的作用。
1.2.5 實時熒光定量PCR 將成熟卵母細胞用0.1% PBS-PVA洗滌3次,轉移到Cell to Signal Lysis Buffer中,待其完全裂解,根據反轉錄試劑盒說明書將其反轉為cDNA。以cDNA為模板,使用SYBR Green PCR試劑盒進行qRT-PCR檢測。以GAPDH為內參基因,特異性qRT-PCR引物序列見表1。反應程序:95 ℃ 30 s,75 ℃ 10 s,60 ℃ 30 s,循環40次。反應體系:cDNA 1 μL,2×ChamQ SYBR qPCR Master Mix 5 μL,引物0.4 μL,RNase Free ddH2O 3.6 μL,總計10 μL。
1.2.7 數據分析 利用Image J軟件分析圖像熒光強度,使用GraphPad-prism 8繪制圖表并進行數據處理,當P<0.05時認為差異顯著。每組試驗均重復3次。

表1 引物序列Table 1 Primers sequence
4.2.2 教師要根據初中生的生理機能和運動能力的變化規律,結合中考項目的要求,改變單一的教學方法,不要被一種教學方法所禁錮,主動嘗試新型的教學方法,爭取達到更好的教學目標。
船舶造價參照2014—2016年國際新造船市場平均值,ARC6型船舶造價在普通船型的基礎上增加30%,船舶折舊方法參照年限平均折舊法,年限25 a,船舶拆船價格參照2014—2016年國際拆船市場平均值。
2 結 果
2.1 SITO對豬卵母細胞成熟及早期胚胎發育的影響
李揚應用現金比率法對我國1984—1997年的游資相對量進行了分析。〔3〕在此基礎上,劉超對1997年至2005年10月國內的社會游資相對量進行了測算。根據李揚和劉超的測算,從1984年以來,M0/(M1-M0)、M0/M1的比率從1984年以來呈現先上升,后下降的過程。M1/(M2-M1)、M0/(M2-M1)的比率總體呈現下降趨勢。尤其1992年全面建立社會義義市場經濟以來,M0/(M1-M0)、M0/M1比率就一直呈下降的趨勢。

A.對照組與處理組卵母細胞成熟率;B.對照組與處理組卵裂率;C.對照組與處理組囊胚率A. Oocyte mature rate in control group and treatment group; B. Cleavage rate of control group and treatment group; C. Blastocyst rate of control group and treatment group圖1 SITO促進卵母細胞成熟和囊胚形成Fig.1 SITO promotes oocyte maturation and blastocyst formation
2.2 SITO對豬卵母細胞ROS水平的影響
如圖2所示,各處理組ROS熒光強度均顯著低于對照組(P<0.05),說明SITO對于減少卵母細胞ROS水平具有顯著作用。其中20 μmol·L-1SITO處理的熒光強度降低最為明顯,因此選擇添加20 μmol·L-1SITO用于后續試驗。

A.對照組與處理組的ROS熒光圖像(Bar=100 μm);B.對照組與處理組卵母細胞ROS熒光值分析A. ROS fluorescence images of control group and treatment group(Bar=100 μm); B. Analysis of ROS fluorescence values of oocytes in control group and treatment group圖2 SITO降低豬卵母細胞中的ROS水平Fig.2 SITO decreases ROS levels in pig oocytes
2.3 SITO對豬卵母細胞凋亡及ΔΨ m水平的影響
1.2.4 細胞凋亡檢測及線粒體膜電位ΔΨm檢測 根據Annexin V-FITC細胞凋亡檢測試劑盒說明書,將成熟卵母細胞用0.1% PBS-PVA洗滌3次,然后加入188 μL Annexin V-FITC結合液、5 μL Annexin V-FITC、2 μL Mito tracker Red CMXROs染色液、5 μL Hoechst 33342染色液,輕輕混合,室溫避光孵育30 min,隨后置于冰浴,染色完成再洗滌3次后封片。根據線粒體膜電位檢測試劑盒(JC-1)說明書,將成熟卵母細胞用0.1% PBS-PVA洗滌3次后,在10 mmol·L-1JC-1中于38.5 ℃、5% CO2下孵育30 min,染色完成再洗滌3次后封片,在共聚焦顯微鏡下觀察。
如圖5所示,20 μmol·L-1SITO處理后BCL-2、SOD、CAT、TFAM基因的表達上調,Caspase3和BAX的表達下調。表明20 μmol·L-1SITO處理具有抗凋亡、抗氧化以及促進線粒體DNA復制和轉錄的作用。
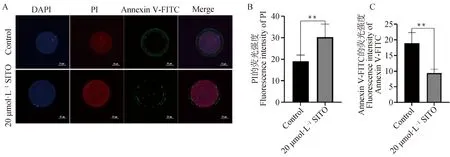
A. 對照組和20 μmol·L-1處理組激光共聚焦圖像(Bar=50 μm);B. 對照組和20 μmol·L-1處理組紅色熒光強度數據分析;C. 對照組和20 μmol·L-1處理組綠色熒光強度數據分析A. Laser confocal images of control group and 20 μmol·L-1 treatment group(Bar=50 μm); B. Analysis of red fluorescence intensity data in control group and 20 μmol·L-1 treatment group;C. Analysis of green fluorescence intensity data in control group and 20 μmol·L-1 treatment group圖3 SITO減少卵母細胞凋亡Fig.3 SITO decreased apoptosis of porcine oocytes
2.4 SITO處理對豬卵母細胞相關基因表達的影響
康軒版的分數除法都通過單位量的轉換來計算.如通過單位量的轉換得到8個除以15個,即.其用圖形來表征分數除法(如圖1),使得學生很容易理解單位量轉換的過程.
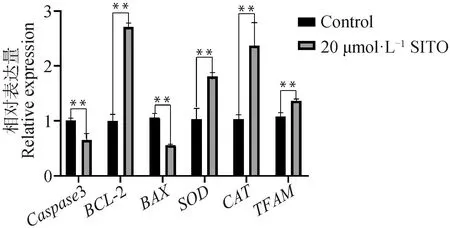
圖5 SITO處理對卵母細胞相關基因表達的影響Fig.5 Effect of SITO treatment on oocyte related genes expression
2.5 SITO對豬孤雌囊胚細胞數的影響
成熟培養液中添加20 μmol·L-1SITO使囊胚率明顯提高(圖1C),經過囊胚免疫熒光染色,如圖6A、B所示,20 μmol·L-1SITO組的囊胚細胞數顯著高于對照組(P<0.05),說明20 μmol·L-1SITO可以提高囊胚細胞總數。
西沙群島不僅擁有豐富的自然旅游資源,也擁有獨具魅力的人文旅游資源。目前西沙群島旅游開放的三個島嶼,主要的旅游活動集中在原生態自然體驗,較少開放人文活動。西沙群島的人文旅游資源主要集中在永興島和石島,主要有:海軍收復西沙紀念碑、石島中國主權碑、西沙將軍林、日本炮樓、西沙海軍博物館、南海島礁保衛戰紀念碑等。在政策和地理環境的允許下,適當穩妥開放西沙群島其他島嶼,增加西沙群島旅游資源的多樣性。
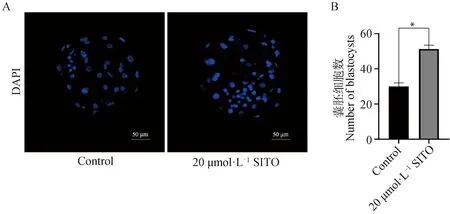
A.對照組和20 μmol·L-1處理組囊胚及免疫熒光染色圖(Bar=50 μm;B.對照組和20 μmol·L-1處理組囊胚細胞數A.Blastocyst and immunofluorescence staining of control group and 20 μmol·L-1 treatment group(Bar=50 μm); B. Total blastocysts in control group and 20 μmol·L-1 treatment group 圖6 SITO增加豬囊胚細胞數Fig.6 SITO increases the total number of pig blastocysts
3 討 論
卵母細胞在正常生理條件下通過線粒體氧化磷酸化產生ROS,但它的過度積累會導致氧化和抗氧化功能的紊亂。ROS會影響細胞活力、破壞細胞結構,甚至導致細胞死亡,最終對胚胎發育產生負面影響,在培養基中添加抗氧化劑或ROS清除劑有助于減少氧化應激,利于胚胎存活和囊胚的形成[22-23],氧化還原相關基因的表達也與體外成熟卵母細胞中的ROS水平有關[24]。過氧化氫酶(CAT)和超氧化物歧化酶(SOD)在抵抗氧化應激中起著重要的作用,具有抵抗自由基的攻擊、保護細胞免受損害的作用[25],在本研究中,SITO可以顯著降低豬卵母細胞的ROS水平,同時CAT和SOD基因的表達被SITO激活,這與之前報道的SIRT1通過調控氧化還原狀態參與卵母細胞成熟的結果一致[26]。還有研究表明,SITO可以通過減少DNA損傷來延長細胞壽命[27],通過增強AMPK磷酸化來激活脂肪體,并具有較強的自由基清除活性[28],自由基是機體生命活動中各種氧化反應的代謝產物,這進一步說明了SITO具有抗氧化的能力。
眾所周知,氧化應激超載可以調節與卵母細胞凋亡相關的BAX/BCL-2比值[29-30]。在線粒體介導的凋亡途徑中,細胞內異常信號導致BAX活化,抑制BCL-2的表達,誘導線粒體釋放細胞色素c,形成凋亡小體或啟動caspase級聯激活,從而導致細胞凋亡的發生[31]。在SITO方面,有報道稱其可誘導細胞外信號調節激酶(ERK)和p38絲裂原活化蛋白激酶(MARK)的磷酸化,下調PI3K/Akt,從而促進細胞凋亡和細胞死亡[32],為了驗證這一點,本研究檢測了Caspase3、BAX和BCL-2的轉錄水平,結果顯示SITO降低了豬卵母細胞Caspase3基因和促凋亡基因BAX的表達,促進了抗凋亡基因BCL-2的表達。結果表明,SITO抑制了卵母細胞的凋亡,這與Moon等[32]的結果不符,推測這是由于SITO的抗氧化能力使卵母細胞的氧化應激顯著減少,從而抑制了細胞凋亡的發生。這也反映出抗氧化作用對于抑制細胞凋亡能夠產生明顯影響。
線粒體是細胞質中必不可少的細胞器,被認為是評價卵母細胞質量的重要標志[33-34],線粒體膜電位ΔΨm是ATP產生的驅動力。在卵母細胞發育成熟的過程中,對能量的需求是遞增的,直到排卵時達到高峰,此時線粒體發生氧化磷酸化產生ATP,同時也產生一定量的ROS[35-37]。本研究結果表明,添加SITO后ΔΨm有所增加,這表明SITO具有提高線粒體膜電位,促進ATP產生的作用,這有利于卵母細胞成熟。線粒體的生物合成和功能主要受線粒體DNA復制和轉錄的直接調節因子TFAM的調控[38-39],在本研究中,SITO處理使TFAM表達上調,因此,認為SITO可能通過激活線粒體功能減少細胞凋亡。
另外,囊胚細胞總數體現了早期胚胎發育的質量[40],本研究結果顯示,SITO處理的囊胚細胞總數顯著高于對照組,這表明成熟液中添加SITO能顯著提高體外成熟卵母細胞的早期胚胎發育潛力。
4 結 論
綜上所述,本研究通過在IVM培養基中添加SITO,結合免疫熒光染色,說明SITO對豬卵母細胞體外發育起重要作用,SITO增強了豬卵母細胞抗氧化和抗凋亡能力,增強了線粒體膜電位和ATP的含量。因此,在豬卵母細胞IVM中添加β-谷甾醇對支持和改善卵母細胞質量和早期胚胎發育具有重要作用。
