定植早期多效唑處理對甘薯塊根發育及其蔗糖代謝的影響
2024-06-09 15:12:40趙添辛曙麗黃哲瑞劉永華朱國鵬
熱帶作物學報 2024年4期
趙添 辛曙麗 黃哲瑞 劉永華 朱國鵬
關鍵詞:甘薯;多效唑;蔗糖代謝;蔗糖分解酶;塊根發育
中圖分類號:S531 文獻標志碼:A
甘薯[Ipomoea batatas(L.)Lam.]屬旋花科一年生或多年生草本植物,其塊根富含淀粉、可溶性糖、膳食纖維、蛋白質、維生素、礦物質以及多酚、多糖、花青素等活性成分,是我國重要的糧食、經濟、工業原料作物[1-2]。據聯合國糧農組織(FAO)最新數據顯示,2020 年我國甘薯種植面積為225 萬hm2,約占全世界種植面積的30.27%,總產量為4920 萬t,約占全世界總產量的54.97%,穩居世界首位[3]。雖然我國甘薯種植面積和總產量極大,但平均單產較前20 年的數據相比,并未進一步提升,一直徘徊在22 t/hm2左右,和發達國家相比仍有一定差距[3]。同時,甘薯品質相較發達國家也有待提升,特別是我國甘薯的淀粉和干物質含量均相對較低[4]。
多效唑(paclobutrazol,PBZ)作為具有生長調節作用的三氮唑類化合物之一,在農業上廣泛應用[5]。對塊根/塊莖類作物施加PBZ,可抑制莖葉的生長,增加其根冠比,促進光合同化產物向作物產品器官的分配,從而增加產量,如火蔥的鱗莖[6]、馬鈴薯的塊莖[7-8]、胡蘿卜的直根等[9]。此外,PBZ 還可促進葉綠體分化,促進葉綠素的生物合成,防止葉綠素降解,從而提高光合速率,達到增產的效果[10-11]。甘薯生產上常發生因莖葉徒長而導致產量下降的現象,而噴施PBZ 也可有效抑制莖葉徒長,從而增加塊根產量[11-12]。除了增加產量外,施用PBZ 還可以提高甘薯塊根的品質。如PBZ 處理可將甘薯塊根的淀粉含量提高42%[13];此外,PBZ 處理還會提高塊根中的蛋白質含量和淀粉的崩解值,有利于改善甘薯的食味[11]。
甘薯產量的形成主要受到單株結薯數和單薯重的影響。研究表明,PBZ 處理可顯著提高單薯重和結薯數2 種指標中的一種或2 種來增加甘薯產量。例如,在正常栽培條件下,封壟期進行PBZ處理可通過增加單薯重來提高甘薯品種北京553塊根產量,但對單株結薯數無顯著影響[13]。同樣在品種北京553 上,封壟期(定植后46 d)的高濃度PBZ 處理雖然顯著提高了甘薯結薯數,但卻導致單薯重出現輕微下降[14]。此外,定植后50 d的PBZ 處理不但提高濟薯25 的單薯重,而且還顯著增加其結薯數[15]。目前有關PBZ 提升塊根/塊莖類作物產量和品質的研究主要集中在馬鈴薯上,對甘薯的研究相對較少,尚不清楚PBZ 影響單薯重、結薯數以及淀粉含量的生理生化機制。
植物光合產生的蔗糖不僅是合成淀粉的原料,其分解產生的己糖還可作為糖信號調控各種新陳代謝過程和植物器官(包括塊根/塊莖)的生長發育[16]。在蔗糖代謝過程中,蔗糖分解酶扮演著重要角色,當蔗糖從葉片運輸到庫器官(如種子、果實和塊根/塊莖)后,首先被蔗糖分解酶分解為己糖或己糖衍生物,然后再用于淀粉的合成和其他各種生物代謝過程[16]。在植物體內蔗糖分解酶共有兩大類,分別為蔗糖合成酶(sucrosesynthase,Sus)和轉化酶(invertase,INV)。根據INV 的亞細胞定位,可將轉化酶分為細胞壁轉化酶(cell wall invertase,CWIN)、液泡轉化酶( vacuolar invertase , VIN) 和細胞質轉化酶(cytoplasmic invertase,CIN)[17]。
蔗糖分解酶在植物塊根/塊莖的發育中發揮著不可或缺的重要作用。例如,利用反義RNA 技術分別下調胡蘿卜中CWIN、VIN、Sus 基因的表達均會導致胡蘿卜肉質根的生長發育受到顯著抑制[18-19]。在馬鈴薯中下調Sus 基因表達則導致塊莖數量減少,同時淀粉含量也下降[20-21]。因此,推測蔗糖代謝在甘薯產量和品質形成中也發揮著重要的調控作用,而PBZ 可能通過調控蔗糖分解酶的活性來影響甘薯塊根的生長發育和產量形成。
本研究以海南鮮食甘薯主栽品種高系14 為實驗材料,在定植早期(定植后20~30 d)對其進行PBZ 處理后測定塊根產量、單薯鮮重、單株結薯數等指標,以明確PBZ 處理對甘薯產量的影響。其次,測定PBZ 處理對不同發育階段(定植后30、60、90、120 d)塊根中可溶性糖和淀粉含量、4種蔗糖分解酶活性的影響,同時測定PBZ 處理對蔗糖分解酶基因家族成員表達水平的影響。上述研究可初步闡明PBZ 調控甘薯塊根發育的蔗糖代謝相關生理生化機制并確定其中的關鍵蔗糖分解酶基因,為甘薯高產、穩產甘薯新品種的選育提供理論依據。
1 材料與方法
1.1 材料
本研究于2022 年1—4 月在海南大學實驗基地大棚內進行,試驗材料選取海南甘薯主要栽培品種高系14。在大棚內采用盆栽種植的方式,根據甘薯每個采樣階段設置處理組與對照組,處理組在甘薯定植后20 d 和30 d,采用100 mg/L 的PBZ 進行葉面噴施處理,每個階段每組設置4 個生物學重復。定植前將土壤與有機肥按3∶1 混勻,再按每盆0.8 g 的量加入史丹利復合肥(N∶P∶K=15∶15∶15)混勻后,分裝至每個直徑約30 cm 的花盆。定植時打孔深度約15 cm,用75%的酒精消毒后的剪刀剪取約20 cm 長,頂端含有莖尖的甘薯莖段進行盆栽扦插,莖段切口為平切口。取樣時期為定植后30、60、90、120 d,對甘薯塊根進行取樣。每個樣品的取樣重量為0.2 g,于液氮中速凍后置于–80 ℃超低溫冰箱中保存備用。
1.2 方法
參照LUNN 等[22]的酶學方法測定甘薯塊根中葡萄糖、果糖及蔗糖含量;參照SMITH 等[23]的酶學方法測定甘薯塊根中的淀粉含量;采用分光光度計法,具體參照TOMLINSON 等[24]的方法測定甘薯塊根中轉化酶和蔗糖合成酶活性。
根據試劑盒的方法,使用總RNA 提取試劑盒[康為世紀OmniPlant RNA Kit(Dnase I)全能型植物RNA 提取試劑盒]從處理組與對照組4 個生長階段的塊根中提取總RNA,接著使用超微量分光光度計測定總RNA 的濃度與質量,并通過凝膠電泳檢查總RNA 的完整性;然后采用高質量的總RNA 作為模板,使用諾唯贊HiScript III 1st StrandcDNA Synthesis Kit 逆轉錄試劑盒合成cDNA;最后使用前人已發表以及近期本課題組篩選和設計的20 個關鍵蔗糖分解酶基因和5 個SWEET 基因的特異引物用于qRT-PCR 測定(表1),使用諾唯贊ChamQ Universal SYBR qPCR Master Mix 試劑盒,在德國耶拿qTOWER3G 定量PCR 儀上進行基因表達水平的測定。qRT-PCR 反應程序如下:95 ℃預變性5 min;95 ℃變性5 s,60 ℃退火30 s,72 ℃延伸30 s,40 個循環。采用2–ΔΔCT法對qRTPCR的測定結果進行計算,以甘薯塊根內ACTIN基因為內參基因。
1.3 數據處理
利用Excel 2020 軟件作圖,使用SPSS 26.0軟件的t-test 功能進行差異顯著性分析。
2 結果與分析
2.1 PBZ 對甘薯地上部生物量和塊根產量的影響
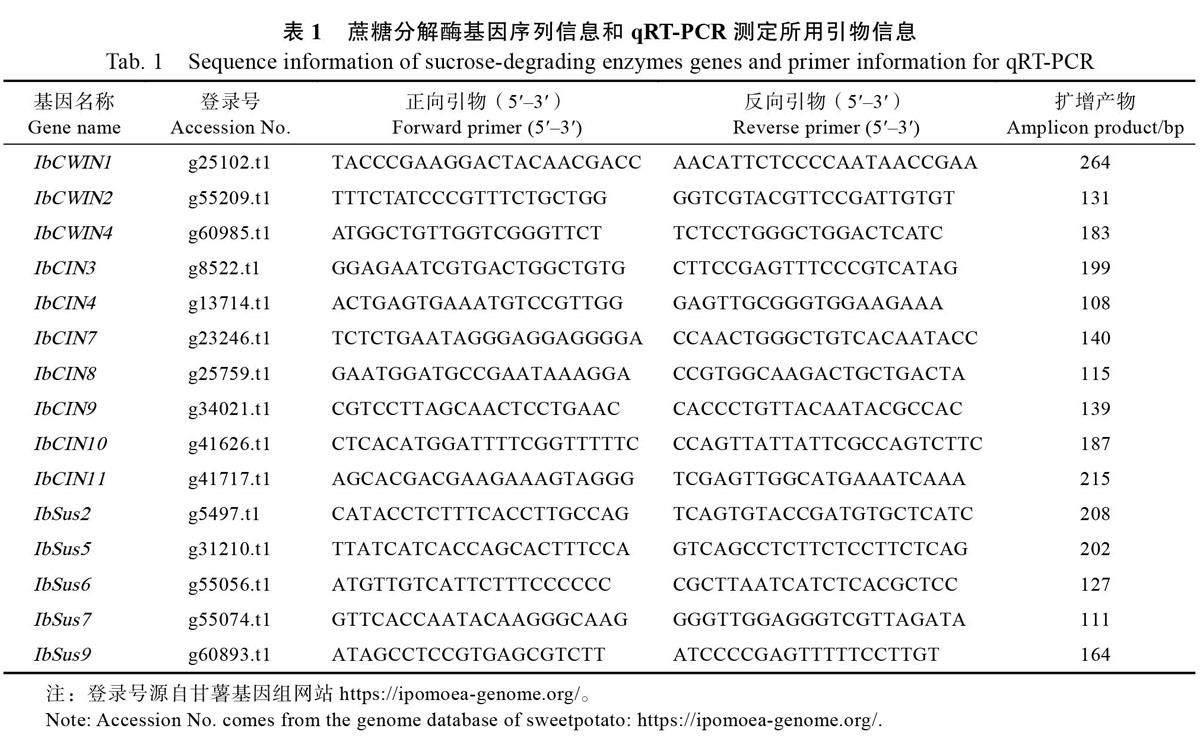
葉面噴施PBZ 導致高系14 的地上部(莖葉)鮮重呈現出低于對照的趨勢,特別是在定植后60 d時,PBZ 處理顯著降低了高系14 的地上部鮮重(圖1A)。在地下部薯塊鮮重方面,PBZ 處理的單株薯塊鮮重在定植后30、60 d 大于對照組,并在定植后60 d 單株薯塊鮮重顯著大于對照,但PBZ處理并沒有提高定植后90、120 d 的薯塊鮮重(圖1B)。上述數據表明,定植早期的PBZ 處理(定植后20、30 d 噴施)會在短時間內(定植后60 d)抑制甘薯地上部生長,提高甘薯地下部薯塊的產量,但隨著時間的延長,PBZ 的效果逐漸消失。
為進一步分析PBZ 處理導致薯塊產量增加的具體原因,對單薯鮮重和單株結薯數進行測定。結果表明,PBZ 處理顯著提高定植后60 d 的單薯鮮重,但在其他3 個時期,PBZ 處理和對照之間均無顯著差異(圖1C)。此外,PBZ 處理對單株結薯數無顯著影響(圖1D)。上述數據表明,PBZ處理對甘薯塊根的增產效果主要是通過增加單薯鮮重來實現。
2.2 PBZ 對甘薯塊根中可溶性糖與淀粉含量的影響
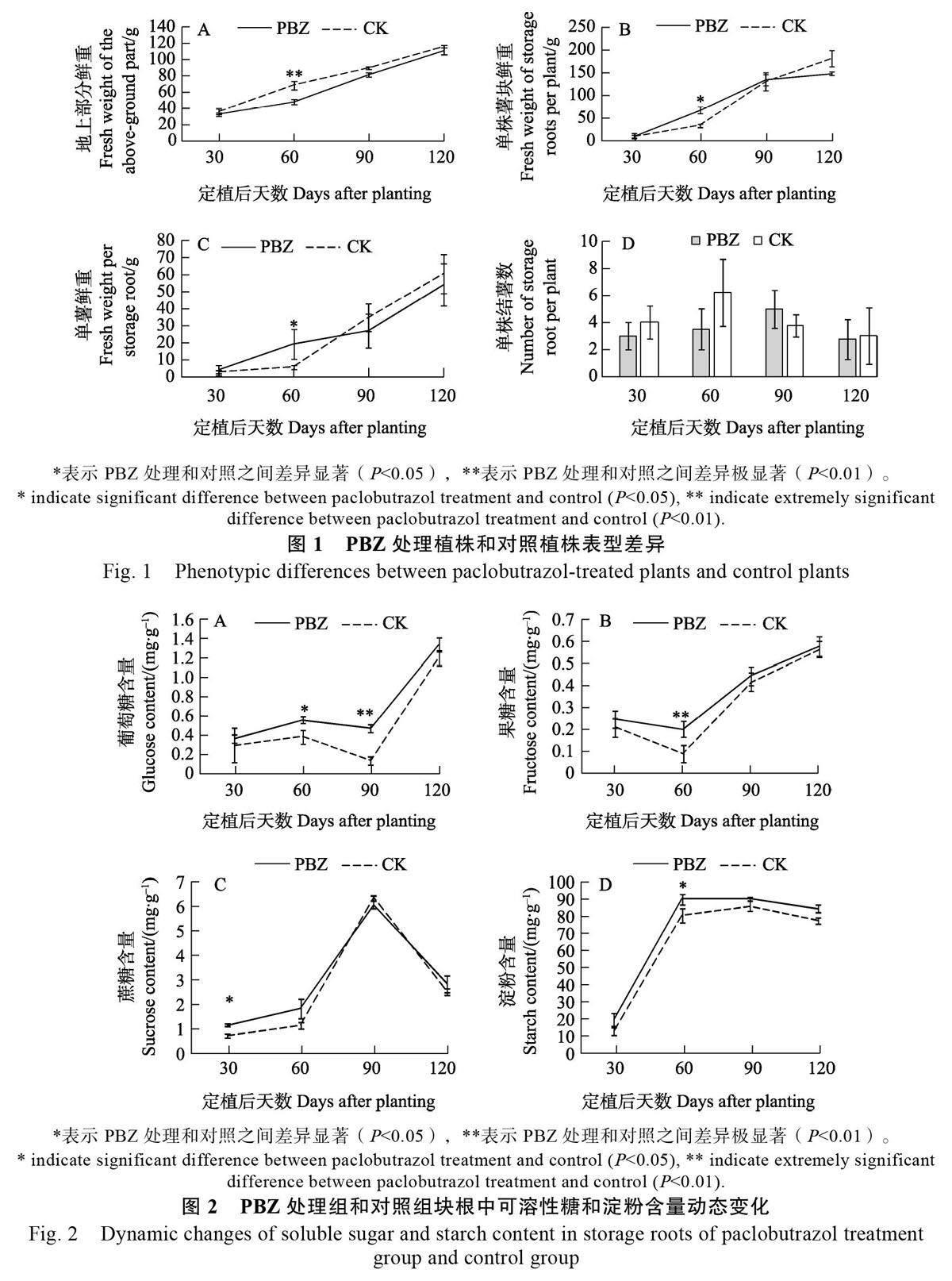
為進一步研究PBZ 處理對甘薯品質的影響,對定植后30、60、90、120 d 甘薯塊根中可溶性糖(葡萄糖、果糖、蔗糖)和淀粉含量的動態變化進行測定。結果表明,PBZ 處理可提高塊根中的葡萄糖和果糖含量(圖2A、圖2B),特別是PBZ 處理組的葡萄糖含量在定植后60 d 顯著高于對照組,在定植后90 d 極顯著高于對照組;此外,PBZ 處理組的果糖含量在定植后60 d 極顯著高于對照組。PBZ 處理塊根中蔗糖含量在定植后30 d顯著高于對照,而在其他3 個時期則均無顯著差異(圖2C)。
對淀粉含量的測定表明,塊根中的淀粉含量在定植后30~60 d 均迅速增加,在發育后期(定植后90~120 d)淀粉含量呈現略微下降趨勢(圖2D)。與可溶性糖含量類似,PBZ 處理塊根中淀粉含量在4 個時期均高于對照組塊根,并且在定植后60 d 顯著高于對照組。總體來看,PBZ 處理可在短時間內(定植后30~60 d)顯著提高塊根中的可溶性糖和淀粉含量,但隨著時間的延長,PBZ處理的效應逐漸消失。
2.3 PBZ 對甘薯塊根中蔗糖分解酶活性的影響
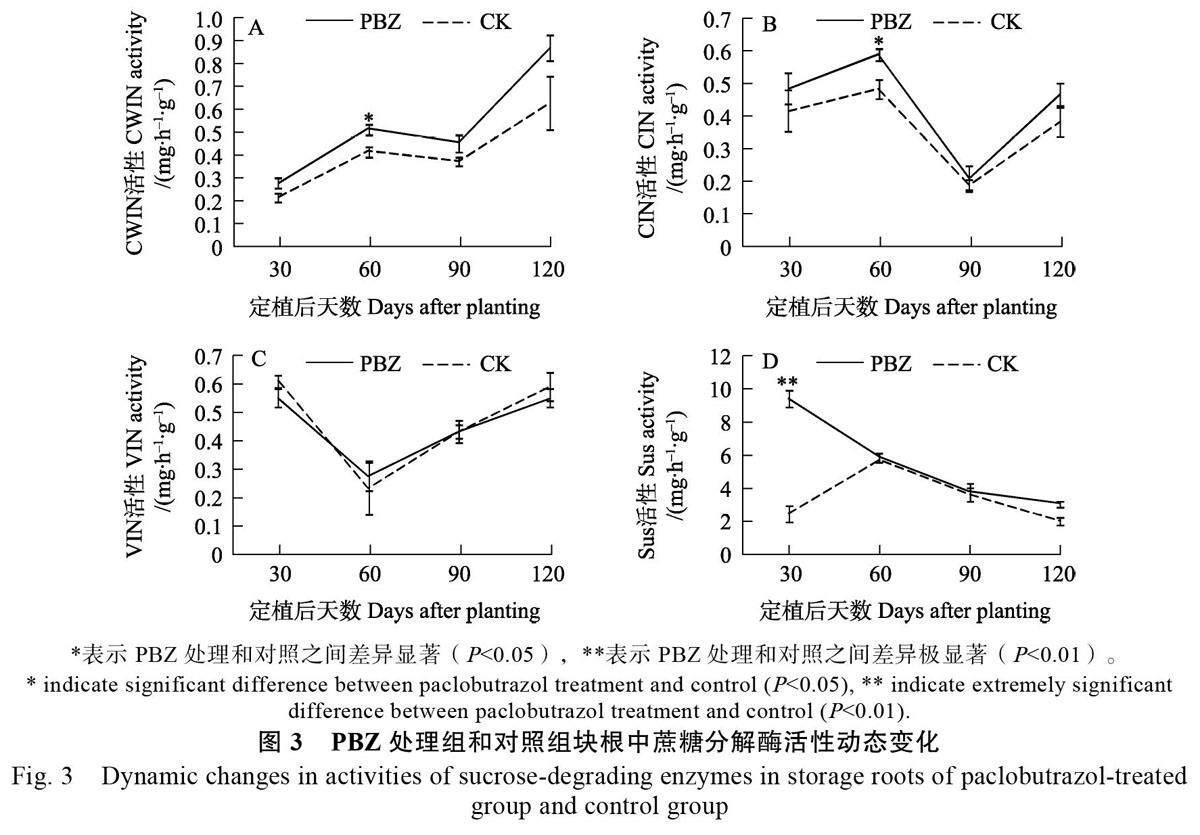
可溶性糖含量測定表明,與對照相比,PBZ處理提高了塊根中的可溶性糖和淀粉含量,且在處理后的短時期內表現為顯著增加,推測這可能是由于PBZ 處理對蔗糖分解酶活性的影響而導致的。對甘薯塊根中蔗糖合成酶和3 種蔗糖轉化酶活性的測定表明,塊根中Sus 的活性要遠高于3種轉化酶中的任何一種,且PBZ 確實影響了蔗糖分解酶的活性(圖3)。具體來看,PBZ 處理導致塊根中的CWIN 活性上升,特別是在定植后60 d,PBZ 處理塊根中的CWIN 活性顯著高于對照組塊根;而在其他3 個時期,處理組的CWIN 活性雖然高于對照組,但差異不顯著(圖3A)。PBZ 處理組的CIN 活性總體呈現出高于對照組的趨勢,且在定植后60 d 顯著高于對照組,在其余3 個時期無顯著差異(圖3B)。
此外,與CWIN 和CIN 不同,PBZ 處理組對VIN 活性的影響無明顯規律,且PBZ 處理組和對照組的VIN 活性在4 個時期均無顯著差異(圖3C)。PBZ 處理組的Sus 活性在4 個時期均高于對照組(圖3D),且在定植后30 d 極顯著高于對照組;而在其余3 個時期,處理組和對照組之間均無顯著差異。綜上所述,PBZ 處理總體上呈現出提高塊根中CWIN、CIN 和Sus 活性的作用,特別是在處理后的短時間內(定植后30~60 d)可顯著提高CWIN、CIN 和Sus 的活性;但是,PBZ 處理在4 個時期內對VIN 活性均無顯著影響。
在受到PBZ 誘導的3 種蔗糖分解酶中,Sus受到PBZ 誘導的時間(定植后30 d)要早于CWIN和CIN 活性受到誘導的時間(定植后60 d),而且Sus 活性受PBZ 誘導的幅度要遠高于CWIN 和CIN 活性受誘導的幅度。此外,PBZ 處理組和對照組的3 種轉化酶活性在4 個時期的動態變化趨勢一致,即PBZ 處理并未對3 種轉化酶活性隨塊根發育的動態變化規律產生影響。然而,PBZ 處理對Sus 活性隨塊根發育的動態變化規律產生了顯著的影響,對照組的Sus 活性呈先上升后下降的趨勢(圖3D),而PBZ 處理組的Sus 活性始終呈現不斷下降的趨勢。上述Sus 與CWIN/CIN 之間的差異表明Sus 可能在PBZ 調控甘薯塊根發育中發揮著更為重要的作用。
2.4 PBZ 處理對蔗糖分解酶基因家族成員表達的影響
上述蔗糖分解酶活性的測定表明,PBZ 處理可通過提高塊根中蔗糖分解酶(CWIN、CIN 和Sus)的活性來增加塊根中的可溶性糖和淀粉含量,因為高活性的蔗糖分解酶可增加塊根的庫強,加快蔗糖由光合葉片向塊根的轉運并合成淀粉進行儲藏。近期張文杰[25]從甘薯基因組中共鑒定出12 個CIN 基因和9 個Sus 基因,其中7 個CIN基因(IbCIN3、IbCIN4 和IbCIN7~11)和5 個Sus基因(IbSus2、IbSus5~7 和IbSus9)在塊根中高表達。此外,我們從甘薯基因組進一步鑒定出4個CWIN 基因(表1)。利用已發表的CIN 和Sus的引物以及自己設計CWIN 引物,我們研究了PBZ 處理對上述在塊根中高表達的CIN、Sus 和CWIN 基因家族成員表達水平的影響,以明確PBZ 是通過調控哪些特定基因家族成員來實現對甘薯塊根中糖分和淀粉含量的控制,從而最終影響到塊根的膨大和發育。
測定結果表明,在4 個CWIN 基因中,只有3 個CWIN 基因(IbCWIN1、IbCWIN2 和IbCWIN4)在塊根中有表達(圖4A~圖4C)。塊根中這3 個CWIN 基因的表達量均是在塊根發育早期(定植后30 d)最高,隨著塊根的發育(60~120 d)快速下降(圖4A~圖4C)。PBZ 處理顯著提高IbCWIN1 在30 d 的表達量(圖4A)。此外,PBZ 處理還顯著提高IbCWIN2 在60 d 以及IbCWIN4 在60 d 和90 d 的表達量(圖4B、圖4C)。7 個在塊根高表達的CIN 基因中,PBZ處理只對其中3 個CIN 基因(IbCIN3、IbCIN7和IbCIN8)表達量產生了顯著影響(圖4D~圖4F),對其他4 個CIN 基因并無顯著影響。具體來看,PBZ 顯著提高了IbCIN3、IbCIN7 和IbCIN8在定植后60 d 的表達量(圖4D~圖4F)。此外,PBZ 還顯著提高了IbCIN7 在90 d 的表達量(圖4E)。
與 CWIN 和CIN 類似,在塊根中高表達的5個Sus 基因的表達也均受到PBZ 處理的誘導。特別是在定植后30 d,IbSus2、IbSus5-7 和IbSus9的表達水平表現為PBZ 處理組顯著高于對照組,其中IbSus5、IbSus7、IbSus9 表現為極顯著差異(圖5A~圖5E)。此外,IbSus7 的表達在定植后90 d 也受到PBZ 的顯著誘導(圖5D)。
3 討論
前人的研究表明,PBZ 處理效果存在著不確定性,其可通過提高單薯重和結薯數2 種指標中的任何一種或同時提高2 種指標來增加甘薯產量[13-15]。這種不確定性可能和PBZ 處理的時間有關,目前已有的研究大多數PBZ 處理時間發生在植株封壟以后,大約在定植后50 d 左右[13-15]。此時,甘薯根60 d),且Sus 活性受PBZ 誘導的幅度要遠高于CWIN 和CIN 活性受誘導的幅度。Sus 與CWIN/IN在這2 個方面的差異表明Sus 可能在PBZ 調控甘薯塊根發育中發揮著更為重要的作用。研究表明,INV 和Sus 分別在植物器官發育的不同時期發揮著不同的功能。通常INV 活性在器官發育早期較高,主要和細胞分裂和器官的形成相關,而Sus活性在器官發育后期較高,主要和淀粉、纖維素的合成以及細胞和器官的膨大相關[28-31]。因此,INV 對植物生長發育的影響要大于Sus,前者可以影響庫器官的數量,而后者則主要影響庫器官的大小。例如,VIN 在棉花上的突變導致產生完全無纖維的棉花種子[30],而Sus 突變則主要抑制棉花種子上纖維的伸長生長而對纖維數量影響不大[31]。
在胡蘿卜上,VIN 和CWIN 的突變導致肉質根的膨大完全受到抑制,而Sus 的突變只是導致肉質根變小,并不影響其數量[18-19]。本研究結果表明,PBZ 處理改變了Sus 活性隨塊根發育的動態變化規律,且PBZ 處理導致塊根Sus 活性在定植后30 d 極顯著上升,增加幅度高達500%,這極大地促進塊根中淀粉的積累和塊根的膨大。雖然PBZ在定植后60 d 的CWIN 和CIN 活性也顯著上升,但上升幅度相對較小,最終表現為Sus 活性遠高于CWIN 和CIN 活性的總和。因此,推測較早受到PBZ 誘導且活性較高的Sus 可能會導致塊根中的蔗糖主要被Sus 分解,從而用于淀粉的合成和塊根的膨大,這會導致經過CWIN 和CIN 分解的蔗糖的量不足,最終抑制新塊根的形成。
雖然CWIN 和CIN 沒有對結薯數產生影響,但其可能對甘薯的品質產生一定的影響。和INV分解蔗糖產生葡萄糖和果糖不同,Sus 分解蔗糖產生的是UDP-葡萄糖和果糖,但不產生游離的葡萄糖。因此,PBZ 處理下塊根中游離葡萄糖含量的升高很可能是由于CWIN 和CIN 活性上升導致。此外,CWIN 和CIN 產生的葡萄糖和果糖還可以通過呼吸途徑為淀粉的合成和塊根的生長提供能量供應。因此,PBZ 處理下,Sus、CWIN 和CIN 三者共同作用的最終結果為PBZ 處理顯著增加了單薯鮮重和塊根中的可溶性糖和淀粉含量,但對單株薯塊數量無顯著影響。
甘薯基因組中共有9 個Sus 基因,其中5 個Sus 基因(IbSus2、IbSus5-7 和IbSus9)在塊根中高表達[25]。這5 個Sus 基因的表達量在定植后30 d均受到PBZ 處理的誘導,其中IbSus5、IbSus7、IbSus9 表現為極顯著差異,而IbSus2 和IbSus6 受到顯著誘導。這可以很好地解釋定植后30 d 甘薯塊根中Sus 活性受到PBZ 誘導的現象。
此外,甘薯基因組中共鑒定出12 個CIN 基因,其中7 個CIN 基因(IbCIN3、IbCIN4 和IbCIN7-11)在塊根中高表達[25]。這7 個CIN 基因中,PBZ 處理只對其中3 個CIN 基因(IbCIN3、IbCIN7 和IbCIN8)表達產生誘導作用,且誘導主要發生在定植后60 d,這和CIN 活性的誘導主要發生在60 d 相一致。從甘薯基因組中共鑒定出4個CWIN 基因中,其中只有3 個CWIN 基因(IbCWIN1、IbCWIN2 和IbCWIN4)在塊根中有表達。和CWIN 活性在定植后60 d 受到PBZ 誘導相對應,IbCWIN2 和IbCWIN4 表達量在定植后60 d 受到顯著誘導。和甘薯產量指標以及淀粉和可溶性糖含量指標類似,PBZ 對蔗糖分解酶活性及其基因表達水平的影響也主要發生在PBZ 處理后的短時期內(定植后30~60 d),而在定植后期(90~120 d)PBZ 的處理效果消失。
綜上所述,PBZ 處理下Sus 表達和活性增加的時間更早、幅度更大,這可以促進淀粉的快速積累和塊根的快速膨大。而隨后CWIN、CIN 表達和活性的上升則可為淀粉的合成和塊根的膨大提供能量供應,同時增加塊根中的可溶性糖含量,改善甘薯品質。本研究還鑒定出上述3 種蔗糖分解酶基因家族中受到PBZ 誘導表達的特定成員。PBZ 對甘薯塊根產量、品質、蔗糖分解酶活性及其基因表達水平的影響主要發生在PBZ 處理后的短時期內(即定植早期);隨著PBZ 處理時間的延長也就是在定植后期PBZ 對上述指標的影響逐漸消失。因此,生產上為了提高甘薯產量,有必要在全生育期進行PBZ 處理。上述研究結果可為下一步通過品種選育和栽培等措施實現甘薯優質、高產提供理論依據。
