葉下珠黃化植原體和叢枝植原體的分子鑒定
2024-06-09 15:21:47林兆威牛曉慶唐慶華王曄楠孟秀利宋薇薇
熱帶作物學(xué)報(bào) 2024年4期
林兆威 牛曉慶 唐慶華 王曄楠 孟秀利 宋薇薇

關(guān)鍵詞:葉下珠;黃化;叢枝;植原體;分子鑒定
中圖分類號(hào):S763.7 文獻(xiàn)標(biāo)志碼:A
葉下珠(Phyllanthus urinaria L.),又名珍珠草、夜合草,是大戟科(Euphorbiaceae Juss)葉下珠屬(Phyllanthus)一年生草本植物。在我國(guó)主要分布于廣西、廣東及海南等南方地區(qū)。帶根全草可入藥,具有很高的藥用價(jià)值。研究發(fā)現(xiàn),葉下珠在抗乙肝病毒(HBV)、抗腫瘤、抗菌、抗內(nèi)毒素、抗血栓形成及免疫調(diào)劑等方面具有較好的藥理作用[1]。近年來(lái),葉下珠常用于臨床上抗HBV 的研究,對(duì)乙型肝炎的治療有巨大的開發(fā)潛力。
植原體(phytoplasma)是存在于植物韌皮部的篩管細(xì)胞及介體昆蟲的腸道、淋巴及唾液腺等組織內(nèi)的一類專性菌,主要通過(guò)刺吸式介體昆蟲進(jìn)行植物間傳播。被植原體侵染的植物常引起叢枝、葉片黃化、花變?nèi)~及矮化等癥狀[2]。至今已有1000 余種植物被植原體侵染,我國(guó)已報(bào)道100多種植原體病害[3-4] , 其中棗瘋病( jujubewitches-broom, JWB)[5]、小麥藍(lán)矮病(wheat bluedwarf disease, WBD)[6]、甘蔗白葉病(sugarcanewhite leaf, SCWL)[7]及泡桐叢枝病(paulowniawitches-broom, PaWB)[8]等危害嚴(yán)重,對(duì)相關(guān)產(chǎn)業(yè)造成嚴(yán)重的經(jīng)濟(jì)損失。在海南,植原體侵染引起的檳榔黃化病(areca palm yellow leaf disease,YLD)是一種毀滅性病害[9],每年因該病造成的經(jīng)濟(jì)損失高達(dá)20 億元以上[10]。
2022 年8 月,本研究團(tuán)隊(duì)在海南省文昌市文城鎮(zhèn)的1 個(gè)檳榔園內(nèi)進(jìn)行黃化病調(diào)查,發(fā)現(xiàn)該園有葉片表現(xiàn)黃化的葉下珠和1 株表現(xiàn)叢枝的葉下珠,為典型的植原體侵染引起的癥狀。前期采用熒光定量PCR 檢測(cè)技術(shù)(專利申請(qǐng)?zhí)枺?02210933142.8),對(duì)葉下珠黃化和叢枝2 種癥狀進(jìn)行植原體檢測(cè),結(jié)果發(fā)現(xiàn)2 種癥狀均顯示植原體陽(yáng)性,且病原含量分別為6.8×103、4.6×108copies/μL。目前國(guó)內(nèi)外尚無(wú)葉下珠黃化植原體和叢枝植原體分類鑒定的相關(guān)報(bào)道,因此,本研究分別對(duì)葉下珠黃化、叢枝植原體16S rDNA 基因和核糖體蛋白基因(rp)進(jìn)行克隆,通過(guò)分子鑒定的手段明確葉下珠黃化植原體和叢枝植原體的分類地位,并與檳榔黃化植原體16S rDNA 進(jìn)行分析,為檳榔黃化病中間寄主的研究提供理論依據(jù)。
1 材料與方法
1.1 材料
1.1.1 供試材料 待測(cè)葉下珠黃化、叢枝樣品采自海南省文昌市文城鎮(zhèn)(19°33′13″N, 110°47′14″E)。葉下珠黃化癥狀表現(xiàn)為整株黃化,主要為葉肉黃化,葉肉葉脈黃綠相間明顯,后期發(fā)展為葉肉葉脈黃化(圖1A~圖1D),該樣品編號(hào)為HN-Pu2;葉下珠叢枝癥狀表現(xiàn)為整株叢枝,葉片變小(圖1E~圖1F),該樣品編號(hào)為HN-Pu1;健康樣品(圖1G ) 采自海南省文昌市文城鎮(zhèn)( 19°33′14″N,110°47′15″E)。
1.1.2 主要試劑與儀器 pMD18-T 載體購(gòu)自TaKaRa 公司;2×Taq PCR Mix、植物基因組DNA提取試劑盒及瓊脂糖凝膠回收試劑盒購(gòu)自TIANGEN 公司;DH5α 感受態(tài)細(xì)胞購(gòu)自上海唯地生物技術(shù)有限公司。主要儀器:1~1000 μL 移液器、GD120 恒溫水浴鍋、5810R 離心機(jī)及BiometraTOne 96G 梯度PCR 等。
1.2 方法
1.2.1 葉下珠葉片總DNA 提取 剪取葉片樣品0.1 g,參照植物基因組DNA 提取試劑盒說(shuō)明書,進(jìn)行樣品DNA提取,并將提取的DNA置于–20 ℃中保存,備用。
1.2.2 16S rDNA 和rp 基因的PCR 擴(kuò)增 以待測(cè)樣品葉片總DNA 為模板,健康樣品為陰性對(duì)照,H2O 為空白對(duì)照進(jìn)行PCR 擴(kuò)增。采用巢氏PCR技術(shù)檢測(cè)葉下珠黃化的植原體,采用一步PCR 檢測(cè)葉下珠叢枝的植原體。16S rDNA 和rp 基因擴(kuò)增引物序列見(jiàn)表1,16S rDNA 基因的巢氏PCR擴(kuò)增的第1 步反應(yīng)引物為P1/P7,第2 步反應(yīng)引物為R16F2n/R16R2;rp 基因的巢氏PCR 擴(kuò)增的第1 步反應(yīng)引物為rpF1/rpR1,第2 步反應(yīng)引物為rpF2/rpR2;其中,巢氏PCR 第1 步反應(yīng)結(jié)束后,需將PCR 產(chǎn)物稀釋50 倍作為模板進(jìn)行第2 步反應(yīng)。所需引物均由生工生物工程(上海)股份有限公司合成。擴(kuò)增反應(yīng)體系:2×Taq PCR Mix 12.5 μL,DNA 模板為2.0 μL,ddH2O 為8.5 μL,上、下游引物(10 μmol/L)各1.0 μL。引物P1/P7 擴(kuò)增程序?yàn)椋?5 ℃預(yù)變性5 min;95 ℃變性1 min,48 ℃退火1 min,72 ℃延伸2 min,共35 個(gè)循環(huán);72 ℃延伸10 min。引物R16F2n/R16R2 擴(kuò)增程序?yàn)椋?5 ℃預(yù)變性5 min;95 ℃變性45 s,55 ℃退火45 s,72 ℃延伸90 s,共35 個(gè)循環(huán);72 ℃延伸10 min。rp 基因的引物rpF1/rpR1 和rpF2/rpR2 擴(kuò)增程序參照林兆威等[11]的方法。
1.2.3 PCR 產(chǎn)物純化、克隆及序列測(cè)定 經(jīng)瓊脂糖凝膠回收試劑盒對(duì)PCR產(chǎn)物的目標(biāo)條帶進(jìn)行回收純化,并將目的片段與pMD18-T 載體連接后轉(zhuǎn)至DH5α 感受態(tài)細(xì)胞中,在抗生素平板中進(jìn)行涂板于37 ℃培養(yǎng),通過(guò)菌落PCR 進(jìn)行陽(yáng)性驗(yàn)證,將陽(yáng)性轉(zhuǎn)化子送深圳華大基因股份有限公司測(cè)序。
1.2.4 16S rDNA 和rp 基因序列一致性及系統(tǒng)發(fā)育樹分析 經(jīng)測(cè)序獲得的16S rDNA 和rp 基因通過(guò)NCBI 在線blastn 程序(https://www.ncbi.nlm.nih.gov/)進(jìn)行同源性檢索分析,在GenBank 數(shù)據(jù)庫(kù)中下載不同組或亞組的代表性植原體16 rDNA基因序列和rp 基因序列,利用MEGA 7.0 軟件,采用鄰接法(neighbor joining),bootstrap 復(fù)數(shù)設(shè)置為1000,分別構(gòu)建16S rDNA 和rp 基因的系統(tǒng)發(fā)育進(jìn)化樹,分析葉下珠黃化植原體和叢枝植原體分類地位,并將16S rDNA 和rp 基因序列提交至GenBank 數(shù)據(jù)庫(kù)。
1.2.5 植原體16S rDNA 序列虛擬RFLP 分析葉下珠黃化植原體和叢枝植原體的16S rDNA 基因通過(guò)植原體在線分類鑒定軟件iPhyClassifier( https://plantpathology.ba.ars.usda.gov/cgi-bin/resource/iphyclassifier.cgi)進(jìn)行相似性分析和虛擬RFLP 分析,確定植原體的分類地位[16]。
2 結(jié)果與分析
2.1 植原體16S rDNA 和rp 基因片段的獲得
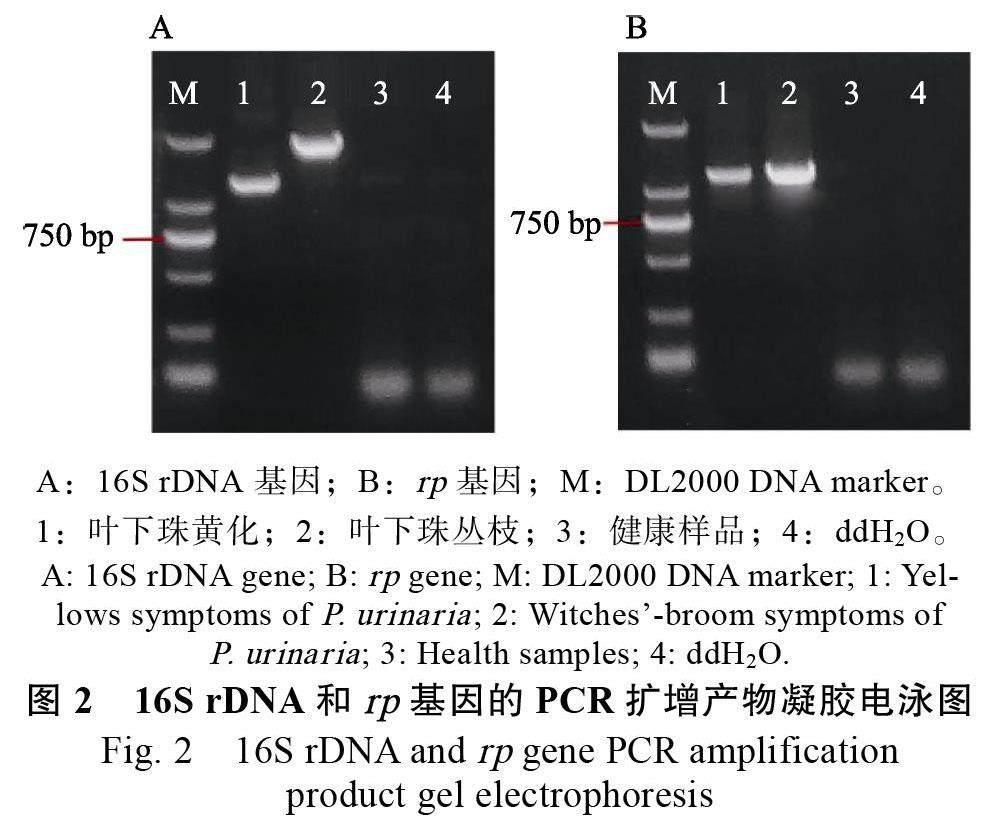
從葉下珠黃化樣品中擴(kuò)增植原體16S rDNA基因,獲得約1.2 kb 的片段;擴(kuò)增rp 基因,獲得約1.2 kb 的片段。從葉下珠叢枝樣品中擴(kuò)增植原體16S rDNA 基因,獲得約1.8 kb 的片段;擴(kuò)增rp 基因,獲得約1.2 kb 的片段(圖2)。獲得的片段大小均與預(yù)期目的片段大小基本一致,且健康樣品和ddH2O 中未擴(kuò)增出相應(yīng)片段。將以上片段進(jìn)行測(cè)序比對(duì),結(jié)果為植原體,因此將該葉下珠黃化和叢枝植原體分別暫命名為葉下珠黃化植原體海南株系(PuY-HN-2022)、葉下珠叢枝植原體海南株系(PuWB-HN-2022)。
2.2 16S rDNA、rp 基因序列一致性及系統(tǒng)發(fā)育樹分析
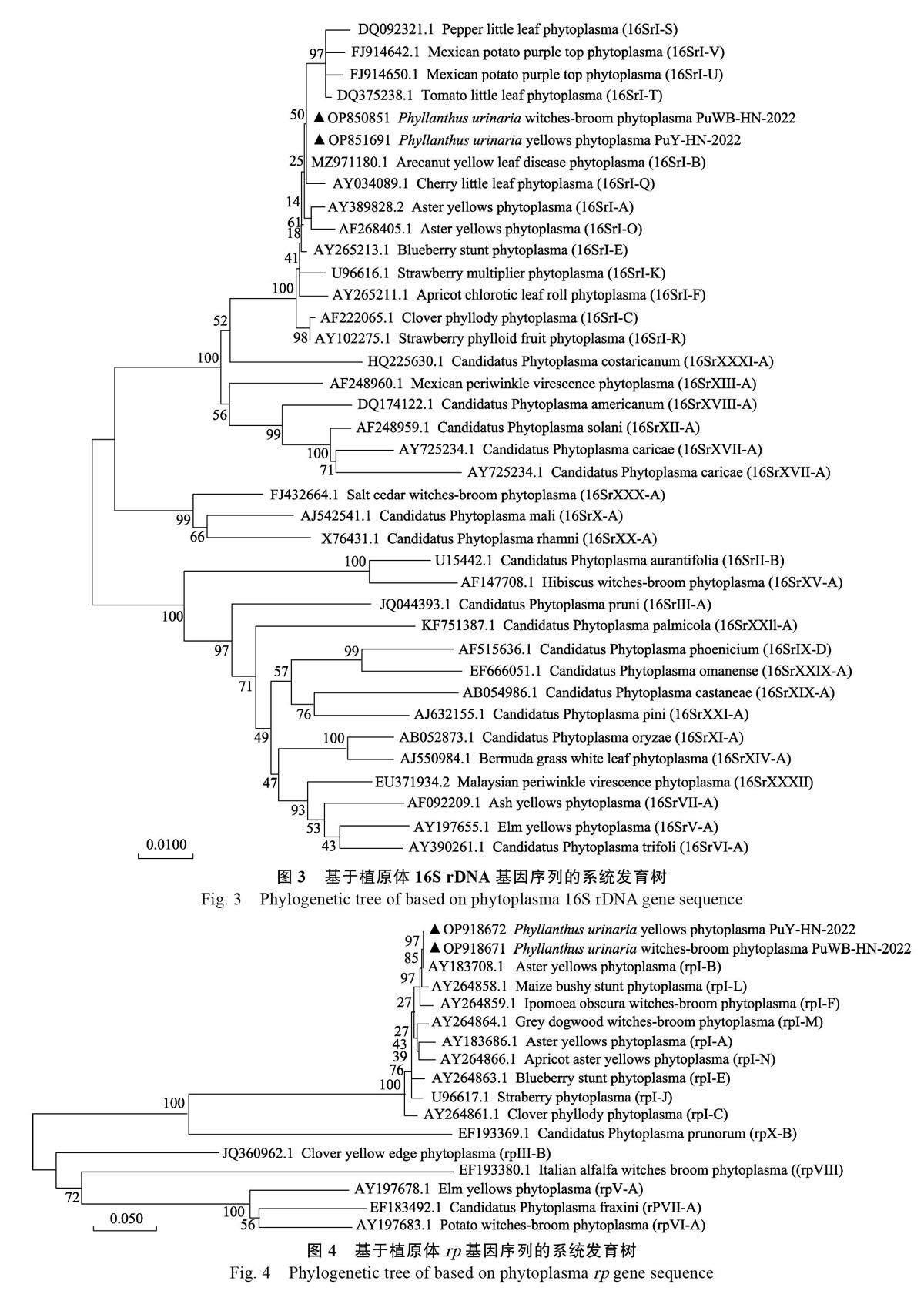
通過(guò)克隆基因目的片段,獲得葉下珠黃化植原體16S rDNA 片段1246 bp(登錄號(hào):OP851691),rp 基因片段1212 bp(登錄號(hào):OP918672);葉下珠叢枝植原體16S rDNA 片段1827 bp(登錄號(hào):OP850851), rp 基因片段1240 bp(登錄號(hào):OP918671)。經(jīng)基因序列一致性分析,葉下珠黃化植原體和叢枝植原體與16SrⅠ組植原體的16SrDNA 序列一致性高達(dá)98%以上。其中,葉下珠黃化植原體與歐洲油菜矮縮植原體(登錄號(hào):MG599470.1)的16S rDNA 序列一致性達(dá)100%;葉下珠叢枝植原體與多氈毛苧麻叢枝植原體(登錄號(hào):MT708485.1)、柳葉菜花變?nèi)~植原體(登錄號(hào):AY101386.1)的16S rDNA 序列一致性達(dá)99.84%;二者均與檳榔黃化植原體海南株系(登錄號(hào):MZ971180.1)的16S rDNA 序列一致性達(dá)100%。葉下珠黃化植原體、叢枝植原體的rp 基因序列均與rpⅠ組的植原體一致性高達(dá)99%以上,且與洋蔥簇生植原體(登錄號(hào):GU228514.1)一致性達(dá)100%。經(jīng)系統(tǒng)發(fā)育樹分析,葉下珠黃化植原體和叢枝原體16S rDNA 基因均與翠菊黃化組(16SrⅠ)植原體聚于一個(gè)大分支,且與16SrⅠ -B 亞組的檳榔黃化植原體( 登錄號(hào):MZ971180.1)聚于同一個(gè)小分支,親緣關(guān)系接近;與其他組植原體親緣關(guān)系較遠(yuǎn)(圖3)。葉下珠黃化植原體和叢枝植原體rp 基因均與翠菊黃化組(rpⅠ)植原體聚于一個(gè)大分支,且與rpⅠ-B 亞組的翠菊黃化植原體(登錄號(hào):AY183708.1)聚于同一個(gè)小分支,親緣關(guān)系接近;與其他組植原體親緣關(guān)系較遠(yuǎn)(圖4)。根據(jù)2004 年比較支原體學(xué)國(guó)際研究規(guī)劃(International Research Projectfor Comparative Mycoplasmology, IRPCM)對(duì)植原體暫定種的劃分規(guī)則[17],葉下珠黃化植原體和叢枝植原體劃為16SrⅠ組。
2.3 虛擬RFLP 分析
將葉下珠黃化植原體和叢枝植原體的16SrDNA 基因序列進(jìn)行虛擬限制性片段長(zhǎng)度多態(tài)性( restriction fragment length polymorphism,RFLP)分析,獲得的圖譜與16SrⅠ-B 的洋蔥黃化植原體(登錄號(hào):AP006628)的參考圖譜相同(圖5),且相似系數(shù)為1.00。綜上,該葉下珠黃化植原體海南株系和葉下珠叢枝植原體海南株系均屬于16SrⅠ-B 亞組成員。
3 討論
由于植原體極難人工分離培養(yǎng),在植原體的分類鑒定上難以通過(guò)形態(tài)觀察、生理生化指標(biāo)進(jìn)行分類鑒定,通常采用16S rDNA 基因、rp 基因及tuf 基因等植原體保守序列進(jìn)行一致性比較,且植原體的組/ 亞組的劃分標(biāo)準(zhǔn)依據(jù)為引物R16F2n/R16R2 擴(kuò)增16S rDNA 基因序列的17 種限制性內(nèi)切酶的RFLP 分析[18-19]。本研究通過(guò)克隆植原體16S rDNA 和rp 基因,并對(duì)基因序列進(jìn)行一致性分析、系統(tǒng)發(fā)育樹分析及16S rDNA 虛擬RFLP 分析,將葉下珠黃化植原體海南株系和葉下珠叢枝植原體海南株系鑒定為16SrⅠ-B 亞組成員。目前關(guān)于葉下珠屬植物受植原體侵染為害的報(bào)道較少,本文為首次報(bào)道植原體侵染為害葉下珠。本研究發(fā)現(xiàn),葉下珠均受16SrⅠ-B 亞組的植原體侵染,但表現(xiàn)出不同的癥狀,在前期的植原體檢測(cè)中發(fā)現(xiàn)這2 種癥狀的植原體含量有數(shù)量級(jí)的差異。因此推測(cè),導(dǎo)致葉下珠表現(xiàn)不同癥狀的原因與植原體侵染的含量差異有關(guān)。
在植原體的各組中,16SrⅠ組成員最多,為29 個(gè)亞組[20],16SrⅠ-B 亞組是翠菊黃化組植原體中寄主范圍最廣的亞組之一。其中,16SrⅠ-B 亞組的植原體侵染引起的檳榔黃化病在海南省發(fā)生面積占27.89%,對(duì)海南檳榔產(chǎn)量造成嚴(yán)重?fù)p失[21]。YU 等[22-23]研究發(fā)現(xiàn),檳榔黃化植原體株系與細(xì)圓藤叢枝植原體、辣椒黃化皺縮植原體的親緣關(guān)系很近,同源性可達(dá)100%,并推測(cè)細(xì)圓藤[Pericampylusglaucus (Lam.) Merr.]和辣椒(Capsicum annuumL.)可能是檳榔黃化植原體的中間寄主。本研究發(fā)現(xiàn),葉下珠黃化植原體和叢枝植原體的16SrDNA 基因序列與檳榔黃化病植原體海南株系16S rDNA 基因序列(登錄號(hào):MZ971180.1)的一致性達(dá)100%;在系統(tǒng)發(fā)育樹中,與檳榔黃化植原體的親緣關(guān)系接近;本研究團(tuán)隊(duì)前期對(duì)該檳榔園進(jìn)行檳榔黃化病檢測(cè)時(shí),發(fā)現(xiàn)該園的黃化病發(fā)生率高,葉下珠黃化和叢枝的發(fā)生也在該檳榔園內(nèi),因此,是否由感染黃化病檳榔作為傳染源引起葉下珠感染將作進(jìn)一步研究。另外,植原體通常通過(guò)刺吸式介體昆蟲傳播[24-25],目前發(fā)現(xiàn)檳榔園中椰子堅(jiān)蚜、長(zhǎng)尾粉蚧、黑刺粉虱、薊馬、棘緣蝽、白盾蚧6 種昆蟲攜帶植原體[26],因此,葉下珠是否會(huì)成為檳榔黃化病通過(guò)媒介昆蟲傳播的中間寄主也將作進(jìn)一步研究。
葉下珠是海南農(nóng)田、林地常見(jiàn)的草本植物,經(jīng)上述分析,葉下珠可能是檳榔黃化病田間的中間寄主。植原體寄主的多樣性可能會(huì)有利于同組或亞組的植原體適應(yīng)不同的環(huán)境,或?qū)⒉煌募闹髦参镒鳛橹虚g介體促進(jìn)植原體的傳播與病害的流行[27-28]。因此,在檳榔黃化病的防控中,不但要切斷媒介昆蟲的傳播,也應(yīng)鏟除中間寄主作為傳播途徑的毒源。
