兩個種源木荷幼苗對干旱-復水的生理生態響應
2024-06-27 10:38:16王德福趙楠邱萬濱桑棟鑫黃國敏劉文飛沈芳芳段洪浪
廣西植物 2024年5期
王德福 趙楠 邱萬濱 桑棟鑫 黃國敏 劉文飛 沈芳芳 段洪浪
DOI: 10.11931/guihaia.gxzw202205011
王德福, 趙楠, 邱萬濱, 等, 2024.
兩個種源木荷幼苗對干旱-復水的生理生態響應 [J].
廣西植物, 44(5): 873-884.
WANG DF, ZHAO N, QIU WB, et al., 2024.
Eco-physiological responses of Schima superba seedlings from two provenances to drought and rewatering [J].
Guihaia, 44(5): 873-884.
摘? 要:? 為探討不同種源樹木對干旱-復水的生理生態響應,該研究以廣東與福建種源木荷為對象,通過盆栽控水方式模擬干旱及復水條件,研究兩個種源木荷的水力及碳生理特征、脯氨酸(Pro)及超氧化物歧化酶(SOD)對干旱-復水的響應。結果表明:(1)對照情況下,廣東種源木荷的莖木質部水勢(Ψxylem)、葉片相對含水量(RWC)、光合速率(Asat)與氣孔導度(Gs)均低于福建種源木荷的。(2)兩個種源木荷的水力特征、Pro與SOD對干旱-復水的響應呈一致趨勢,其中Ψxylem、RWC與Pro均能較快恢復至對照水平,而莖木質部栓塞程度與SOD未恢復至對照水平。(3)福建種源木荷葉片的Asat對干旱的敏感性較廣東種源的高且復水后恢復至對照水平需要更長時間。(4)復水后福建種源木荷非結構性碳水化合物(NSC)的恢復速率高于廣東種源木荷。綜上認為,福建與廣東兩個種源木荷均不能通過短期復水(30 d)來修復被栓塞的木質部。盡管廣東種源木荷的光合速率能夠更快地恢復至對照水平,但其光合速率低于福建種源木荷,并且其NSC的恢復能力較福建種源荷的低。因此,在未來干旱加劇背景下,廣東種源木荷的生長及存活可能受到更大威脅。該研究結果有助于了解種源地氣候條件對樹木抗旱性的影響,為未來森林的經營與管理提供了理論參考。
關鍵詞: 干旱-復水, 光合特征, 水力特征, 木荷, 種內差異
中圖分類號:? Q945
文獻標識碼:? A
文章編號:? 1000-3142(2024)05-0873-12
收稿日期:? 2023-02-26? 接受日期: 2023-03-29
基金項目:? 國家自然科學基金 (31760111); 四川文理學院高層次人才科研啟動項目(2023RC002Z)。
第一作者: 王德福(1993—),博士,講師,研究方向為植物生理生態學,(E-mail)wangdefu16@163.com。
*通信作者:? 段洪浪,博士,教授,研究方向為植物生理生態學,(E-mail)hlduan@gzu.edu.cn。
Eco-physiological responses of Schima superba seedlings
from two provenances to drought and rewatering
WANG Defu1,2,3, ZHAO Nan1, QIU Wanbin1, SANG Dongxin1, HUANG Guomin1,
LIU Wenfei1, SHEN Fangfang1, DUAN Honglang1,4*
( 1. Jiangxi Provincial Key Laboratory for Restoration of Degraded Ecosystems & Watershed Ecohydrology, Nanchang Institute of Technology,
Nanchang 330099, China; 2. Key Laboratory of Vegetation Restoration and Management of Degraded Ecosystems, South China Botanical
Garden, Chinese Academy of Sciences, Guangzhou 510650, China; 3. Research Center for Sichuan Old Revolutionary Base Areas
Development, Sichuan University of Arts and Science, Dazhou 635000, Sichuan, China; 4. Institute for Forest
Resources and Environment of Guizhou, Guizhou University, Guiyang 550025, China )
Abstract:? In the context of global climate change, the research on eco-physiological responses to drought and rewatering among provenances of trees species will help to understand the effect of climate conditions of provenances on drought resistance of trees, further providing theoretical references for cultivation and management of forests in the future. In this study, Schima superba from two provenances in southern China were selected. Potted seedlings were subjected to simulated drought and rewatering conditions. Eco-physiological traits (photosynthesis traits, hydraulic traits, non-structural carbohydrates-NSC, proline concentrations-Pro and superoxide dismutase-SOD activity) of seedlings were examined. The results were as follows: (1) Under well-watered condition, stem xylem water potential (Ψxylem), leaf relative water content (RWC), photosynthetic rate (Asat) and stomatal conductance (Gs) were lower in seedlings from Guangdong provenance compared with Fujian provenance. (2) Responses of hydraulic traits, Pro and SOD to drought and rewatering were consistent between the two provenances. Ψxylem, RWC and Pro could recover to control values rapidly, while percent of stem xylem embolism and SOD could not recover to control values in the end. (3) Asat was more sensitive to drought from Fujian provenance than Guangdong provenance, and it took more time to recover to control in Fujian provenance. (4) The recovery rate of NSC from Fujian provenance was faster than Guangdong provenance. Collectively, seedlings from both provenances could not repair their embolized xylem in the short-term rewatering period (30 days). Although photosynthesis rate in Guangdong provenance could recover to control faster than from Fujian provenance, it was lower than that from Fujian provenance. Furthermore, the recovery rate of NSC was slower in Guangdong provenance than Fujian provenance, indicating that growth and survivals of seedlings from Guangdong provenance may experience greater risk in future characterized with increasing drought stress.
Key words: drought-rewatering, photosynthetic traits, hydraulic traits, Schima superba, intra-specific differences
全球氣候變化背景下,未來降水格局將會發生變化,干旱強度與持續時間均可能增加(Pachauri & Reisinger, 2014)。干旱脅迫會威脅樹木的存活,進而引起群落結構與功能的改變(Barros et al., 2019; Brodribb et al., 2020)。此外,在樹木的生命歷程中,經常會出現多次干旱及干旱-復水現象,樹木也會通過自身調節來應對干旱環境(段娜等,2019; Gessler et al., 2020)。但是,樹木響應干旱-復水的生理生態機制是否存在種內差異還具有很大的不確定性。因此,在全球降水格局變化背景下,研究樹木對干旱-復水生理生態響應策略的種內差異,可為研究樹木如何響應干旱-復水以及生態模型的優化提供基礎數據支撐,也為未來森林的經營與管理提供理論參考。
干旱脅迫通常會抑制樹木生長、導致葉片水勢與光合速率降低、進一步加劇木質部栓塞(Yan et al., 2017; Duan et al., 2019;呂朝燕等,2021),還會引起樹木葉片脯氨酸(Pro)的積累與超氧化物歧化酶(SOD)活性的增強,有助于緩解水分虧缺對植物細胞造成的損傷(Duan et al., 2020; Sun et al., 2020)。此外,干旱脅迫也會改變非結構性碳水化合物(non-structural carbohydrates, NSC)的含量(He et al., 2020)。He等(2020)研究表明干旱對樹木NSC的影響與干旱強度和持續時間有關,輕度干旱下葉片可溶性糖未發生顯著變化,而重度干旱使葉片可溶性糖顯著升高。
干旱脅迫后樹木生理生態指標的恢復能力是評價樹木適應干旱的重要依據。其中,木質部導水率是否能在短期內恢復以及恢復能力的種間差異已成為近年來的研究熱點。葉片水勢能夠較快地恢復至對照水平,而與葉片水勢相比,光合速率與木質部導水率恢復較慢(Duan et al., 2019; Ruehr et al., 2019)。此外,NSC在維持樹木水力功能及莖木質部栓塞修復過程中均發揮著重要作用,栓塞修復時往往伴隨NSC的消耗(Tomasella et al., 2019)。前人研究發現,樹木光合與水分生理指標對干旱-復水的響應不僅因物種不同而存在差異,同一物種不同分布地或種源對干旱-復水的響應也存在差異(劉菲等,2018;陳夢園等,2019);劉菲等(2018)研究發現福建種源油杉的抗旱性大于貴州種源的。然而,樹種不同種源生理生態指標的旱后恢復能力是否有差異仍不清楚,特別是木質部導水率恢復的種內差異仍鮮有報道。
木荷(Schima superba)是山茶科(Theaceae)木荷屬(Schima)大喬木,在我國福建、江西、湖南、廣東等南方地區廣泛分布,是亞熱帶常綠闊葉林的優勢樹種,也是綠化和造林的常用樹種。此外,木荷也是良好的防火樹種,既可與其他樹木混種,也可單獨種植作為防火林帶。然而,不同種源的木荷對干旱-復水的生理生態響應差異還不清楚。以往的研究發現,植物的抗旱能力與當地氣候條件(如年均降雨量)有關(Liang et al., 2019),說明植物對原生地氣候條件具有一定的適應性,并且不同種源的植物對干旱的響應可能存在差異。因此,本研究選取來自具有一定氣候差異的兩個種源木荷作為研究對象,通過盆栽控制干旱與復水條件,研究兩個種源木荷幼苗對干旱-復水的生理生態響應。我們提出的假設:兩個種源木荷的氣體交換與水力特征對干旱-復水的響應存在差異,其中降水量低的種源具有更強的抗旱性。
1? 材料與方法
1.1 材料和實驗設計
在南昌工程學院瑤湖校區(116°01′50.16″ E, 28°41′17.12″ N )開展本實驗。該地區屬亞熱帶季風氣候,年均降雨量1 600~1 700 mm(4—6月約占50%,7—9月約占19%)。從福建尤溪九阜山自然保護區(118°01′58″—118°07′16″ E、26°03′37″—26°06′59″ N)、廣東化陳禾洞自然保護區(113°49′30″—114°01′50″ E、23°43′02″—23°48′10″ N)兩地分別采集木荷種子,兩個種源地的年均降水量分別為1 665、1 801 mm,年均溫分別為19.6 ℃、22.8 ℃。2018年4月對種子進行播種,2019年3月將幼苗移栽至7.6 L的花盆中,每盆1株,盆栽所用土壤為磚紅壤。之后,將所有盆栽搬至遮雨棚(長×寬×高為20 m × 4 m × 3 m)下進行自然生長。遮雨棚頂部所用材料為透明PVC板,遮光率為15%,四周通風良好。實驗開始前所有盆栽的土壤保持在田間持水量,并且每周向土壤施可溶性營養肥料(施可得園藝肥料有限公司,武漢產,N≥30 g·L-1,P2O5≥ 14 g·L-1,K2O≥16 g·L-1,Fe≥0.14 g·L-1,Mn≥0.06 g·L-1)的稀釋液1次,每次200 mL,使幼苗保持良好生長。
待幼苗生長4個月后,對每個種源選取長勢良好、高度一致的幼苗50株作為本實驗的研究對象。實驗設置2個水分處理,即(1)對照:選取20株幼苗,實驗期間保持土壤濕度在田間持水量;(2)干旱-復水:每個種源剩余的30株幼苗,在實驗處理開始后不澆水,使土壤自然變干。期間,監測莖木質部水勢變化情況,當水勢達到前期研究測定的木質部導水率曲線預測以及莖木質部栓塞在88%[木質部導水率下降88%,通常表示嚴重干旱(Uril et al., 2013; Duan et al., 2021)]的水勢附近時,收割樣品,并測定莖木質部栓塞程度(方法見1.2.2)。當莖木質部栓塞程度為88%左右時,對所有幼苗進行復水,使土壤濕度達到并保持在田間持水量,直到實驗結束。
1.2 指標測定方法
1.2.1 葉片氣體交換參數測定? 使用Li-6400便攜式氣體交換系統(LI-Cor, lnc, Lincoln, NE, USA)測定植物的氣體交換參數。分別于干旱的第0天、第2天、第4天、第5天和第6 天及復水后的第3天、第7天和第15 天的上午9:00—11:00對氣體交換參數進行測定。每個種源的每個處理選取4株幼苗(每株選取1片當年生成熟葉片)測定葉片飽和光強光合速率(Asat, μmol·m-2·s-1)、氣孔導度(Gs, mol·m-2·s-1)、蒸騰速率(E, mmol·m-2·s-1)。光源采用人工紅藍光源(6400-2B),光強設置為1 500 μmol·m-2·s-1,二氧化碳濃度設為400 μmol·mol-1,空氣溫度控制在(31.1±0.2)℃,相對濕度控制在60%~80%。
1.2.2 水力特征參數測定
1.2.2.1 水勢? 于正午測定莖木質部水勢,測定之前用保鮮膜與錫箔紙包裹葉片(保鮮膜在內,錫箔紙在外)1 h以上,以保證葉片與木質部之間的水勢平衡(可以用此時的葉片水勢代替莖木質部水勢)。之后,采集葉片于自封袋中并放入保溫箱內保存,帶回實驗室。利用PMS-Model 1505D數顯便攜式植物水勢壓力室(PMS instruments, Corvalis, Oregon USA)測定干旱的第0天、第4天、第6天、第7天、第9天與復水后的第3天和第7天的莖木質部水勢(Ψxylem, MPa)。每個種源的每個處理選取4株幼苗(每株選取2片當年生成熟葉片)。
1.2.2.2 葉片相對含水量(relative water content, RWC)? 首先,將葉片從枝條上取下后,稱量葉片鮮重。然后,于水下用剪刀剪掉一段葉柄,在黑暗處將葉柄浸沒于水中12 h,待葉片充分吸收水分后,將葉片取出,用紙巾擦干表面水分,稱量葉片飽和鮮重。最后,將葉片放入70 ℃烘箱中烘至恒重, 稱取葉片干重。葉片相對含水量=(葉片鮮重-葉片干重)/(葉片飽和鮮重-葉片干重)×100%。葉片相對含水量的測定時間與水勢測定同步。
1.2.2.3 莖木質部栓塞程度(percentage loss of conductivity, PLC)? 先將盆栽放入裝滿水的水桶中,于水下用枝剪將莖取下,用封口膜將切口纏緊;再迅速將莖的下端浸沒于另一水桶中,并用黑色塑料袋罩住。為減少人為因素造成的木質部栓塞,將樣品帶到實驗室后,于水下從莖底部剪掉5 cm,并用黑色塑料袋罩住整個植株1 h左右(莖底部依然浸沒于水中),使木質部張力釋放(Wheeler et al., 2013)。待葉片水勢恢復至大于-1 MPa時,取5~10 cm長的莖(Creek et al., 2018),用XYLEM木質部栓塞測量儀測定莖木質部初始導水率(此過程中莖木質部始終被水浸沒)。隨后用100 kPa的壓力沖洗木質部(約30 min),當木質部末端不再有氣泡出現時停止沖洗,之后用XYLEM木質部栓塞測量儀測定莖木質部的最大導水率(此過程中莖木質部始終被水浸沒)。測定導水率與沖洗木質部所用溶液為2 mmol·L-1的KCl溶液,測定導水率時的壓力梯度為5.4 kPa。莖木質部栓塞程度=(最大導水率-初始導水率)/最大導水率×100%,具體實驗參照Cochard等(2002)的方法。本研究分別測定了復水后的第0天與第30天的莖木質部栓塞程度,每個種源的每個處理選取3~4株幼苗的莖進行測定。
1.2.3 生化指標測定? 非結構性碳水化合物(NSC)[包括可溶性糖(soluble sugar, SS)與淀粉(starch, ST)]含量的測定采用恩酮法。將復水后第0天(干旱臨界點)與第30天的植物各器官樣品烘干并磨成粉末,分別稱取0.05 g于15 mL 離心管中,加入4 mL 80%的酒精,漩渦振蕩。之后置于80 ℃水浴鍋中加熱30 min,待冷卻后以10 000 r·min-1離心8 min,收集上清液于15 mL 離心管中(重復3次)。上層清液用于可溶性糖測定,下層殘渣用于淀粉測定(王德福,2019)。每個種源的每個處理選取4片當年生成熟葉片進行測定。
脯氨酸(proline, Pro)含量測定采用酸性茚三酮顯色法(李合生等,2000)。超氧化物歧化酶(superoxide dismutase, SOD)活性測定采用氮藍四唑比色法(Giannopolitis & Ries, 1977)。每個種源的每個處理選取4片當年生成熟葉片進行測定。
1.3 數據處理
首先,對數據進行正態性及方差齊性檢驗,如果不符合,則進行數據轉換。然后,利用SPSS 19.0 (SPSS Inc. USA)對指標進行單因素方差分析,通過Duncan法進行數據之間的差異顯著性檢驗。最后,利用重復度量來檢驗種源、水分與時間對各指標的影響。所有結果中P<0.05即顯著,采用Sigmaplot 12.5作圖。圖中所有結果均為平均值±標準誤(x±sx)。
2? 結果與分析
2.1 兩個種源木荷幼苗對干旱脅迫的生理生態響應
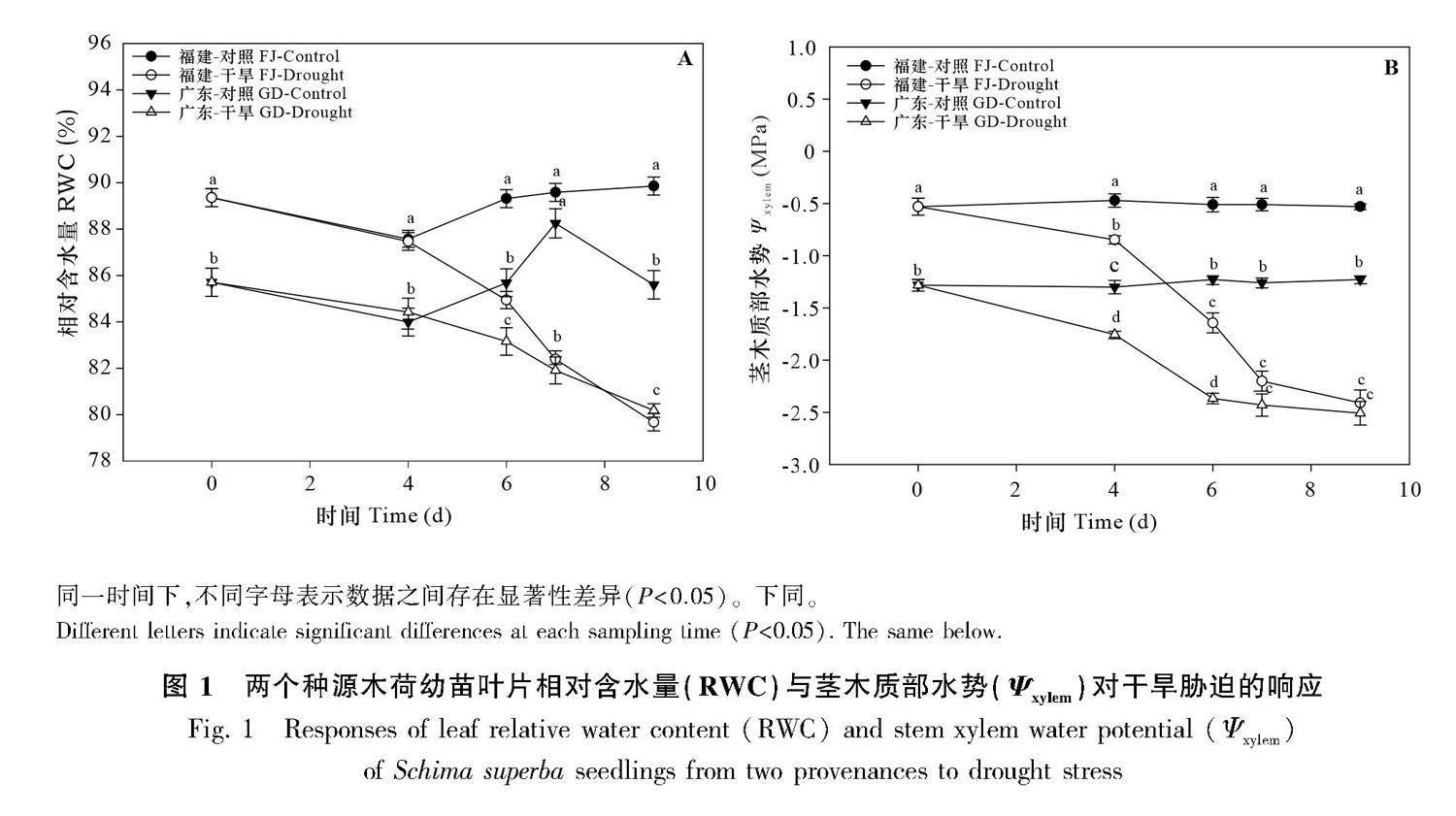
2.1.1水力特征? 由圖1可知,干旱脅迫下,兩個種源木荷葉片相對含水量(RWC)與莖木質部水勢(Ψxylem)均呈下降趨勢。在干旱的第4天,Ψxylem均顯著低于對照水平。干旱的第6 天,RWC均顯著低于對照水平。干旱的第9天(復水0 d)時,Ψxylem均已遠低于對照水平,其中福建種源木荷的Ψxylem降至-2.4 MPa,廣東種源木荷的Ψxylem降至-2.5 MPa。此外,干旱的第9 天,福建與廣東兩個種源木荷的莖木質部栓塞程度(PLC)分別為94%、90%,均已達到88%,達到復水臨界點(圖4)。在對照條件下,廣東種源木荷的Ψxylem與RWC比福建的低。
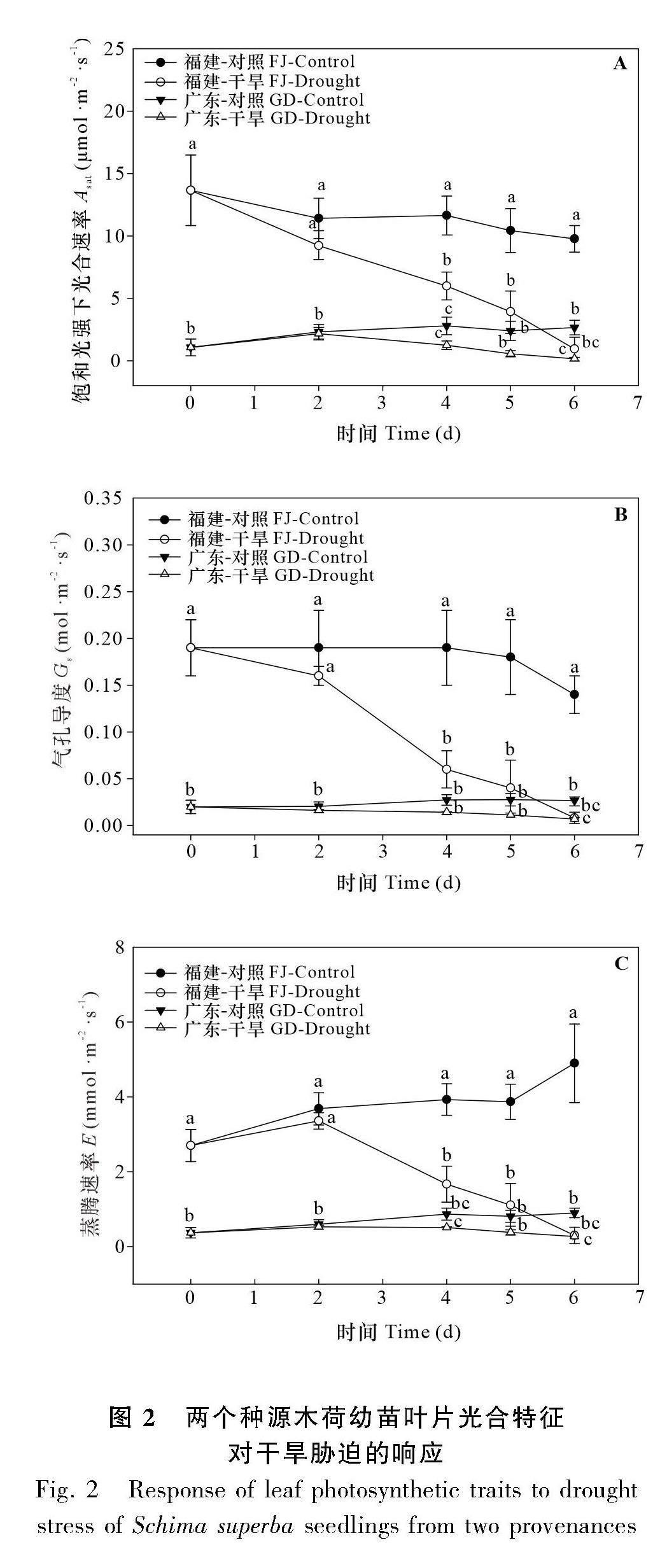
2.1.2 光合特征? 由圖2可知,干旱脅迫下,福建種源木荷的光合速率(Asat)、氣孔導度(Gs)及蒸騰速率(E)較廣東種源的先下降,于干旱脅迫的第4天起,葉片的Asat、Gs、E(5.99、0.06、1.67)均顯著低于對照水平(11.64、0.19、3.93)。廣東種源的木荷,于干旱脅迫的第6天起,葉片的Asat、Gs、E(0.16、0.01、0.27)均顯著低于對照水平(2.66、0.03、0.90)(圖2)。此外,在對照條件下,廣東種源木荷葉片的Asat、Gs始終處于較低水平,并遠低于福建種源木荷的。
2.2 兩個種源木荷幼苗對旱后復水的生理生態響應
2.2.1 水力特征? 復水后,兩個種源木荷的Ψxylem與RWC均升高,于復水后的第3天起,Ψxylem與RWC均恢復至對照水平(圖3)。廣東種源木荷的Ψxylem與RWC均顯著低于福建種源木荷的。種源對Ψxylem與RWC產生顯著影響(表1)。
相對于Ψxylem與RWC,PLC恢復較慢。復水的第30天,福建與廣東種源木荷的PLC(分別為83%、93%)仍顯著高于對照水平(分別為23%、30%),表明莖木質部栓塞未恢復(圖4)。復水的第0天與第30天,在對照與復水條件下,兩個種源木荷的PLC之間均不存在顯著差異。
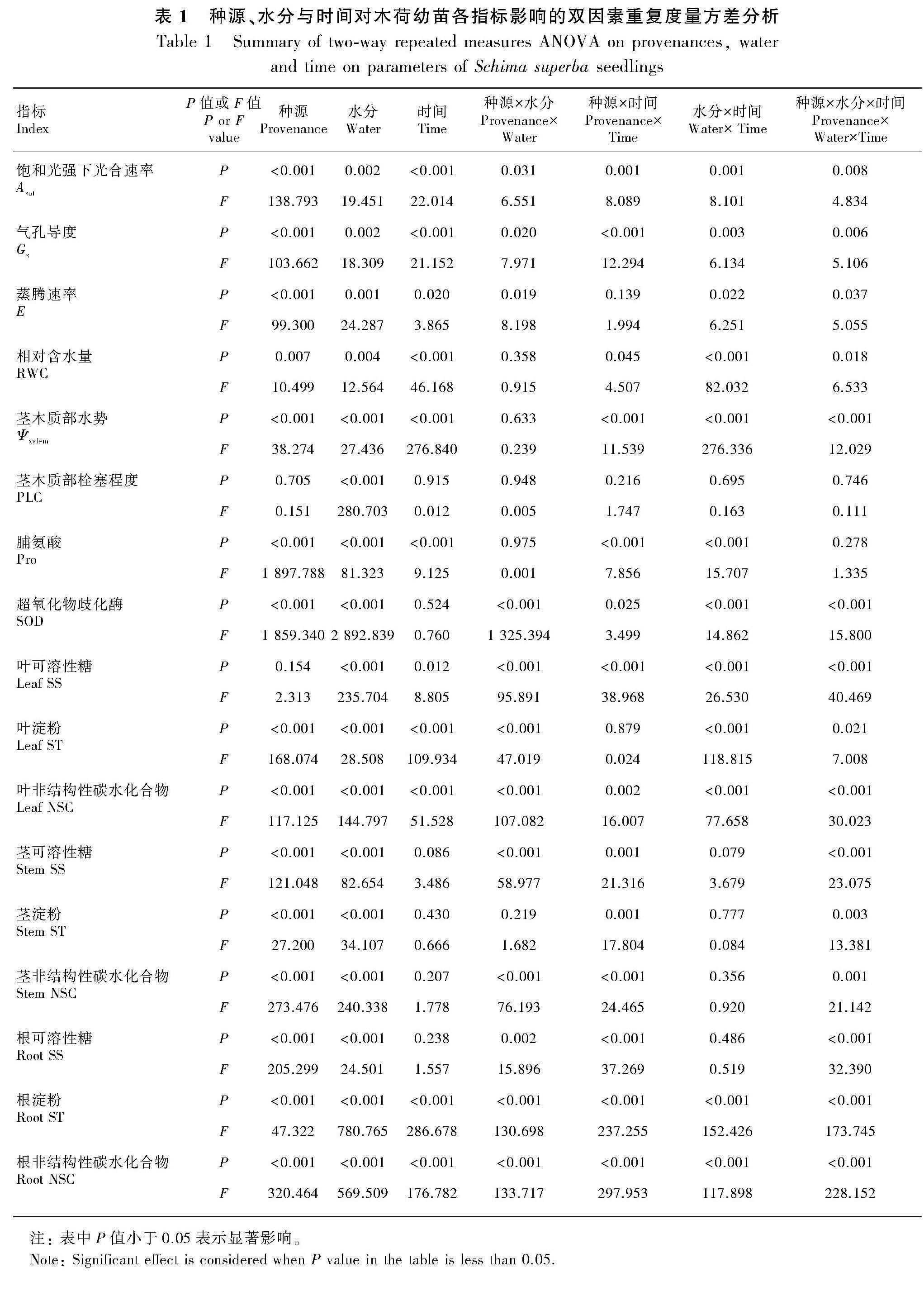
2.2.2 光合特征? 復水后,福建種源木荷葉片的Asat、Gs、E均呈上升趨勢,于第15天均恢復至對照水平(圖5)。對于廣東種源的木荷,復水的第3天,葉片的Asat、Gs、E均恢復至對照水平,但隨著復水的進行,葉片的Asat、Gs出現小幅波動(圖5)。總體分析,在兩個種源木荷中,廣東種源木荷葉片的光合特征恢復較快。這說明種源對葉片Asat、Gs和E產生顯著影響(表1)。
2.2.3 生化特征? 由圖6可知,復水后,福建種源木荷的根、莖與葉的可溶性糖(SS)、淀粉(ST)以及NSC含量均升高,并恢復至或高于對照水平。與福建種源木荷不同,廣東種源木荷莖與根的SS、ST以及NSC的含量均降低,葉片的ST與NSC含量呈升高趨勢(圖6),但各器官NSC含量仍顯著低于對照水平。這說明種源對葉片ST、NSC及根莖SS、ST和NSC產生顯著影響(表1)。
復水后,兩個種源木荷葉片的超氧化物歧化酶(SOD)與脯氨酸(Pro)表現出一致的變化。其中,SOD活性雖逐漸升高,但仍顯著低于對照水平;Pro含量逐步降低,于第15天恢復至對照水平(圖7)。這說明種源對葉片Pro和SOD產生顯著影響(表1)。
3? 討論與結論
3.1 兩個種源木荷幼苗對干旱脅迫的生理生態響應
通常在干旱脅迫下,植物水勢與相對含水量會降低,莖木質部栓塞程度顯著升高(José et al., 2018; Romy et al., 2020)。本研究也發現,干旱脅迫顯著降低了兩個種源木荷莖木質部水勢與葉片相對含水量,莖木質部栓塞程度顯著升高。此外,木荷葉片的氣孔導度、光合速率與蒸騰速率也隨干旱的進行而降低,與以往其他的研究結果類似(陳夢圓等,2019;鄧秀秀等,2020)。但是,在本研究中,福建種源木荷的光合速率、氣孔導度及蒸騰速率對干旱更敏感,較廣東種源的先下降,有助于減少水分的散失。兩個種源木荷的光合生理對干旱的響應存在差異,可能與兩個種源地的年均降雨量(福建的年均降雨量低于廣東地區)有關,是植物適應生境的一種策略(Liang et al., 2019)。非結構性碳水化合物能夠反映植物的碳平衡狀態(鄭云普等,2014)。在干旱脅迫過程中,當樹木呼吸消耗的能量高于光合速率累積的能量時,則會引起樹木體內非結構性碳水化合物的降低(Mcdowell, 2011),進一步干旱可能會導致碳饑餓的發生。本研究中,干旱脅迫顯著降低了兩個種源木荷葉片、莖和根的可溶性糖、淀粉及非結構性碳水化合物濃度,表明干旱脅迫下植物體處于負的碳平衡狀態。在各器官中,福建種源木荷根的非結構碳水化合物降低最多,而廣東種源木荷葉的非結構性碳水化合物降低最多,說明干旱脅迫對樹木非結構性碳水化合物的影響因器官、種源的不同而存在差異(Li et al., 2018)。
當植物遭受干旱脅迫時,細胞內的脯氨酸含量通常會升高以調節滲透,并對酶、蛋白質起保護作用(Sun et al., 2020)。王曦等(2018)研究發現,干旱脅迫下楨楠(Phoebe zhennan)幼苗葉片的脯氨酸含量顯著升高,本研究也發現兩個種源木荷的葉片脯氨酸含量均顯著升高以應對干旱脅迫。干旱脅迫還會引起植物體活性氧的增加,活性氧的存在不僅會對植物的光合系統產生不利影響,還會造成細胞的脂質過氧化與細胞死亡(Xu et al., 2010; Sun et al., 2020)。然而,植物會通過增強活性氧清除酶的活性,如超氧化物歧化酶可清除過多活性氧,減少對植物的傷害(Xu et al., 2010; 劉菲等,2018),但也有研究發現干旱脅迫使超氧化物歧化酶活性降低(Zhang et al., 2017; 吳玲等,2017)。本研究中,干旱脅迫下葉片的超氧化物歧化酶活性顯著降低,可能是木荷葉片超氧化物歧化酶的活性容易遭到活性氧的破壞而下降(吳志華等,2004; 吳玲等,2017)。
3.2 兩個種源木荷幼苗對旱后復水的生理生態響應
干旱復水后,植物水分狀態與細胞膨壓恢復,有助于后期代謝的恢復(Ruehr et al., 2019)。本研究中,兩個種源木荷的莖木質部水勢與葉片相對含水量均于復水后第3天恢復至對照水平,表明兩個種源木荷的枝條與葉片具有同樣的恢復水分狀態的能力(Yan et al., 2017)。復水后葉片相對含水量快速恢復,表明木質部仍具有水分運輸能力(Ruehr et al., 2019)。相比于相對含水量與水勢而言,兩個種源木荷葉片的光合速率、氣孔導度的恢復較為滯后,可能是由脫落酸(Duan et al., 2020)、乙烯(Yao et al., 2020)的抑制作用引起的。本研究發現,廣東種源木荷葉片的光合速率恢復速率高于福建種源木荷的,表明不同種源木荷葉片的光合速率對復水的響應存在差異。兩個種源木荷葉片氣體交換參數恢復速率不一致,可能與以下原因有關;第一,以往研究發現葉片導水率在旱后的恢復能力會影響氣體交換參數的恢復能力(Blackman et al., 2009),因此我們推測本研究中兩個種源木荷氣體交換參數的旱后恢復能力差異與其葉片導水率的恢復有關。第二,脫落酸與乙酸會對氣體交換參數的恢復產生影響(Duan et al., 2020; Yao et al., 2020),本研究中兩個種源木荷氣體交換參數的旱后恢復能力差異可能與植物體內的激素水平有關。因此,在未來的研究中,需要考慮葉片導水率與激素(如脫落酸)水平在復水后的變化情況,為解釋植物氣體交換對復水響應的種間差異提供新的機制。
干旱復水后,雖然植物可以通過木質部栓塞修復或長出新的木質部進而恢復導水率(Cardoso et al., 2020),但木質部栓塞修復所需時間往往高于長出新的木質部所需時間(Brodribb et al., 2010; Martorell et al., 2014)。本研究發現復水30 d后,兩個種源木荷莖木質部栓塞程度仍顯著高于對照水平,表明復水后莖木質部栓塞未修復至對照水平,可能需要更長時間的修復或通過長出新的木質部進行導水率的恢復。此外,復水30 d后福建種源木荷各器官的非結構性碳水化合物逐步恢復至或超過對照水平,而廣東種源木荷的則呈相反趨勢。非結構性碳水化合物對復水后導水率的恢復具有重要貢獻,可以提供滲透調節物質與能量(Tomasella et al., 2019)。因此,相比于福建種源木荷,廣東種源木荷較低的非結構性碳水化合物濃度可能不利于后期莖木質部導水率的恢復。復水后,隨著葉片相對含水量的提高,植物體內滲透調節能力逐漸降低(周歡歡等,2019)。本研究發現,復水后兩個種源木荷的葉片脯氨酸含量逐步降低,并恢復至對照水平;而兩個種源木荷葉片的超氧化物歧化酶活性均逐步升高,可能原因是活性氧在復水后減少,進而對超氧化物歧化酶的破壞降低(吳志華等,2004; Naya et al., 2007)。
本研究測定了兩個種源木荷的生理生態(光合特征、水力特征、NSC、脯氨酸與超氧化物歧化酶)在極端干旱(莖木質部導水率下降88%左右)及復水中的變化情況。研究結果表明:(1)兩個種源木荷均通過關閉氣孔與提高脯氨酸含量來響應干旱脅迫;(2)兩個種源木荷的莖木質部水勢、葉片RWC、Pro、SOD以及莖PLC對干旱復水的響應呈一致趨勢,并且莖PLC在復水30 d后未恢復至對照水平;(3)福建種源木荷的光合速率較廣東種源的先下降且復水后恢復至對照水平需要更長時間;(4)福建種源木荷的NSC恢復速率高于廣東種源木荷的。綜上所述,在未來干旱加劇背景下,福建與廣東兩個種源木荷均不能通過短期復水(30 d)來修復被栓塞的木質部。盡管廣東種源木荷的光合速率能夠更快地恢復至對照水平,但其光合速率低于福建種源木荷的,并且其NSC的恢復能力較福建種源的低。因此,在未來干旱加劇背景下,廣東種源木荷的生長及存活可能受到更大威脅。
參考文獻:
BARROS FDV, BITTENCOURT PRL, BRUM M, et al., 2019. Hydraulic traits explain differential responses of Amazonian forests to the 2015 El Nio-induced drought? [J]. New Phytol, 223(3): 1253-1266.
BLACKMAN CJ, BRODRIBB TJ, JORDAN, GJ, 2009. Leaf hydraulics and drought stress: response, recovery and survivorship in four woody temperate plant species? [J]. Plant Cell Environ, 32(11): 1584-1595.
BRODRIBB TJ, BOWMAN D, NICHOLS S, et al., 2010. Xylem function and growth rate interact to determine recovery rates after exposure to extreme water deficit? [J]. New Phytol, 188(2): 533-542.
BRODRIBB TJ, POWERS J, COCHARD H, et al., 2020. Hanging by a thread?forests and drought? [J]. Science, 368(6488): 261-266.
CARDOSO AA, BILLON LA, BORGES AF, et al., 2020. New developments in understanding plant water transport under drought stress? [J]. New Phytol, 227(4): 1025-1027.
CHEN MY, LI YC, WANG LB, et al., 2019. Photosynthetic responses to drought and subsequent re-watering in seedlings from two different provenances of Quercus variabilis Bl? [J]. Chin J Ecol, 38(10): 2950-2958.? [陳夢園, 李迎超, 王利兵, 等, 2019. 2個種源栓皮櫟對干旱及復水的光合生理響應 [J]. 生態學雜志, 38(10): 2950-2958.]
COCHARD H, COLL L, ROUX XL, et al., 2002. Unraveling the effects of plant hydraulics on stomatal closure during water stress in walnut? [J]. Plant Physiol, 128(1): 282-290.
CREEK D, BLACKMAN C, BRODRIBB TJ, et al., 2018. Coordination between leaf, stem, and root hydraulics and gas exchange in three arid-zone angiosperms during severe drought and recovery? [J]. Plant Cell Environ, 41(12): 2869-2881.
DENG XX, SHI Z, XIAO WF, et al., 2020. Effects of drought and shading on growth and photosynthetic characteristics of Pinus massoniana seedlings? [J]. Acta Ecol Sin, 40(8): 2735-2742.? [鄧秀秀, 施征, 肖文發, 等, 2020. 干旱和遮蔭對馬尾松幼苗生長和光合特性的影響 [J]. 生態學報, 40(8): 2735-2742.]
DUAN HL, DE DIOS VR, WANG DF, et al., 2021. Testing the limits of plant drought stress and subsequent recovery in four provenances of a widely distributed subtropical tree species? [J]. Plant Cell Environ, 45(4): 1187-1203.
DUAN HL, LI YY, XU Y, et al., 2019. Contrasting drought sensitivity and post-drought resilience among three co-occurring tree species in subtropical China? [J]. Agric For Meteorol, 272/273: 55-68.
DUAN HL, WANG DF, WEI, XH, et al., 2020. The decoupling between gas exchange and water potential of Cinnamomum camphora seedlings during drought recovery and its relation to ABA accumulation in leaves? [J]. J Plant Ecol, 13(6): 683-692.
DUAN N, XU J, CHEN HL, et al., 2019. Effects of drought stress on phenotypic plasticity of Cerasus humilis? [J]. Guihaia, 39(9): 1159-1165.? [段娜, 徐軍, 陳海玲, 等, 2019. 干旱脅迫對歐李幼苗表型可塑性的影響 [J]. 廣西植物, 39(9): 1159-1165.]
GESSLER A, BOTTERO A, MARSHALL J, et al., 2020. The way back: recovery of trees from drought and its implication for acclimation? [J]. New Phytol, 228(6): 1704-1709.
GIANNOPOLITIS CN, RIES SK, 1977. Superoxide dismutases, 1:occurrence in higher plants? [Corn, oats, peas]? [J]. Plant Physiol, 59(2): 309-314.
HE W, LIU H, QI Y, et al., 2020. Patterns in nonstructural carbohydrate contents at the tree organ level in response to drought duration? [J]. Glob Change Biol, 26(6): 3627-3638.
JOS PP, SCAR MH, EUSTAQUIO GP, et al., 2018. Cavitation limits the recovery of gas exchange after severe drought stress in Holm Oak (Quercus ilex L.)? [J]. Forests, 9(8): 443-455.
LIU F, ZHOU LT, JIANG Y, et al., 2018. Physiological response from different provenances of Keteleeria fortunei seedlings to drought stress? [J]. J Centr S Univ For Technol, 38(11): 35-45.? [劉菲, 周隆騰, 蔣燚, 等, 2018. 不同種源江南油杉幼苗對干旱脅迫的生理響應 [J]. 中南林業科技大學學報, 38(11): 35-45.]
LI HS, SUN Q, ZHAO SJ, et al., 2000. Principle and technology of plant physiological and biochemical experiment? [M]. Beijing: Higher Education Press: 164-261.? [李合生, 孫群, 趙世杰, 等, 2000. 植物生理生化實驗原理和技術 [M]. 北京: 高等教育出版社: 164-261.]
LI W, HENRIK H, ADAMS HD, et al., 2018. The sweet side of global change-dynamic responses of non-structural carbohydrates to drought, elevated CO2 and nitrogen fertilization in tree species? [J]. Tree Physiol, 38(11): 11-28.
LIANG XY, HE PC, LIU H, et al., 2019. Precipitation has dominant influences on the variation of plant hydraulics of the native Castanopsis fargesii (Fagaceae) in subtropical China? [J]. Agric For Meteorol, 271: 83-91.
L CY, GAO ZX, YAN Y, et al., 2021. Effects of drought-rewatering on leaf water potential of two Dendrobium plants? [J]. Guihaia, 41(2): 177-182.? [呂朝燕, 高智席, 嚴羽, 等, 2021. 干旱-復水對兩種石斛屬植物葉水勢的影響 [J]. 廣西植物, 41(2): 177-182.]
MARTORELL S, DIAZ-ESPEJO A, MEDRANO H, et al., 2014. Rapid hydraulic recovery in Eucalyptus pauciflora after drought: linkages between stem hydraulics and leaf gas exchange? [J]. Plant Cell Environ, 37(3): 617-626.
MCDOWELL NG, 2011. Mechanisms linking drought, hydraulics, carbon metabolism, and vegetation mortality? [J]. Plant Physiol, 155(3): 1051-1059.
NAYA L, LADREA R, RAMOS J, et al., 2007. The response of carbon metabolism and antioxidant defenses of Alfalfa nodules to drought stress and to the subsequent recovery of plants? [J]. Plant Physiol, 144(2): 1104-1114.
PACHAURI R, REISINGER A, 2014. Climate change 2014: synthesis report. Contribution of working groups I, II and III to the fifth assessment report of the intergovernmental panel on climate change? [J]. J Romance Stud, 4(2): 85-88.
ROMY R, ANGELICA C, MARCUS Z, et al., 2020. Drought-induced xylem embolism limits the recovery of leaf gas exchange in Scots pine? [J]. Plant Physiol, 184(2): 852-864.
RUEHR NK, RDIGER G, STEFAN M, et al., 2019. Beyond the extreme: recovery of carbon and water relations in woody plants following heat and drought stress? [J]. Tree Physiol, 1(8): 1-15.
SUN Y, WANG C, CHEN HYH, et al., 2020. Response of plants to water stress: a meta-analysis? [J]. Front Plant Sci, 11(4): 978-985.
TOMASELLA M, PETRUSSA E, PETRUZZELLIS F, et al., 2019. The possible role of non-structural carbohydrates in the regulation of tree hydraulics? [J]. Int J Mol Sci, 21(1): 144-163.
URIL M, PORTE AJ, COCHARD H, et al., 2013. Xylem embolism threshold for catastrophic hydraulic failure in angiosperm trees? [J]. Tree Physiol, 33(7): 672-683.
WANG DF, 2019. The effect of drought and water-logging stresses on eco-physiology of Cinnamomum camphora seedlings? [D]. Nanchang: Nanchang Institute of Technology: 10-11.? [王德福, 2019. 干旱與水淹脅迫對樟樹幼苗生理生態特征的影響 [D]. 南昌: 南昌工程學院: 10-11.]
WANG X, HU HL, HU TX, et al., 2018. Effects of drought stress on the osmotic adjustment and active oxygen metabolism of Phoebe zhennan seedlings and its alleviation by nitrogen application? [J]. Chin J Plant Ecol, 42(2): 240-251.? [王曦, 胡紅玲, 胡庭興, 等, 2018. 干旱脅迫對楨楠幼樹滲透調節與活性氧代謝的影響及施氮的緩解效應 [J]. 植物生態學報,? 42(2): 240-251.]
WHEELER JK, HUGGETT BA, TOFTE AN, et al., 2013. Cutting xylem under tension or supersaturated with gas can generate PLC and the appearance of rapid recovery from embolism? [J]. Plant Cell Environ, 36(11): 1938-1949.
WU L, LI ZH, WU JY, et al., 2017. Response of drought stress on cholorophyll content and anti-oxydant enzyme of Cyclobalanopsis glauca provenance clone seedlings? [J]. J Centr S Univ For Technol, 37(6): 51-55.? [吳玲, 李志輝, 吳際友, 等, 2017. 干旱脅迫對青岡櫟種源葉綠素含量與抗氧化酶活性的影響 [J]. 中南林業科技大學學報, 37(6): 51-55.]
WU ZH, ZENG FH, MA SJ, et al., 2004. A review of advances in active oxygen metabolism in plants under water stress? [J]. Subtrop Plant Sci, 33(2): 77-80.? [吳志華, 曾富華, 馬生健, 等, 2004. 水分脅迫下植物活性氧代謝研究進展(綜述1) [J]. 亞熱帶植物科學, 33(2): 77-80.]
XU Z, ZHOU G, SHIMIZU H, 2010, Plant responses to drought and rewatering? [J].
Plant Signal Behav, 5(6): 649-654.
YAN W, ZHENG S, ZHONG Y, et al., 2017. Contrasting dynamics of leaf potential and gas exchange during progressive drought cycles and recovery in Amorpha fruticosa and Robinia pseudoacacia? [J]. Sci Rep, 7(1): 4470-4481.
YAO GQ, LI FP, NIE ZF, et al., 2020. Ethylene, not ABA, is closely linked to the recovery of gas exchange after drought in four Caragana species? [J]. Plant Cell Environ, 44(2): 399-411.
ZHANG C, LI X, HE YF, et al., 2017. Physiological investigation of C4-phosphoenolpyruvate-carboxylase-introduced rice line shows that sucrose metabolism is involved in the improved drought tolerance? [J]. Plant Physiol Biochem, 115: 328-342.
ZHENG YP, WANG HX, LOU X, et al., 2014. Changes of non-structural carbohydrates and its impact factors in trees: A review? [J]. Chin J Appl Ecol, 25(4): 1188-1196.? [鄭云普, 王賀新,? 婁鑫, 等, 2014. 木本植物非結構性碳水化合物變化及其影響因子研究進展 [J]. 應用生態學報, 25(4): 1188-1196.]
ZHOU HH, FU LC, MA L, et al., 2019. Physiological characteristics of Osmanthus fragrans ‘Boyejingui with drought stress and rewatering? [J]. J Zhejiang A & F Univ, 36(4): 687-696.? [周歡歡, 傅盧成, 馬玲, 等, 2019. 干旱脅迫及復水對‘波葉金桂生理特性的影響 [J]. 浙江農林大學學報, 36(4): 687-696.]
(責任編輯? 蔣巧媛? 王登惠)
