人工老化對水稻種子活力的影響
2024-07-04 13:07:48付坤
中國農業文摘·農業工程 2024年3期
關鍵詞:影響
付坤

摘 要:【目的】研究人工老化對水稻種子活力的影響。【方法】在相對濕度為95%、溫度為40℃的高溫高濕環境內對水稻種子進行人工老化,比較不同老化時間下水稻種子發芽指標、幼苗形態指標及生理生化指標。【結果】在短期高溫高濕老化處理下(0-1d),水稻種子各指標無明顯變化。隨著老化時間的延長,水稻種子活力不斷降低,相對電導率、丙二醛含量升高,可溶性糖含量降低,SOD活性、POD活性先升高后降低,CAT活性持續降低。【結論】實際生產過程中,在高溫高濕條件下,人工老化1天不會對水稻種子造成影響。
關鍵詞:人工老化;水稻種子;活力;影響
水稻作為一種常見的禾谷類作物,在亞洲各國種植極為廣泛,是亞洲人最喜愛的一種主食,水稻增產增收一直備受人們關注[1]。水稻是我國主要糧食作物之一,其高產與種子活力密切相關[2]。種子活力直接決定著種子出苗及成苗的質量性狀,活力越高其抗逆性越強,種子出苗速度、整齊度及發芽率越高,具備優質高產潛能,因而,種子活力會直接影響作物生長及最終產量[3]。近年來,隨著水稻機械化生產水平的不斷提升,高質量種子尤其是高活力種子對水稻優良群體形態及產量提高的作用愈發凸顯[4]。
種子老化是指種子達到生理成熟時種子活力達到最高點,接著種子活力開始降低直至活力喪失甚至死亡的變化過程,是隨著種子貯藏時間的增加而發生的一種自然不可逆過程[5]。水稻種子老化不僅會對種子萌發及幼苗生長造成影響,而且決定著植株后期生長、產量及品質[6]。因此,水稻種子的耐貯藏性是影響水稻種子質量的重要因素之一[7]。
老化方法包括自然老化和人工老化兩種。自然老化是衡量大豆耐儲藏性的直接指標,是利用種子在自然儲藏或種質保存的條件下發芽力逐漸喪失的特征對其進行耐儲藏性研究的方法,但是其歷時較長,很難廣泛應用于實際研究[8]。人工老化是在高溫高濕作用下加快種子老化速度以研究其耐儲藏性的方法[9]。近年來,已有部分學者研究了人工老化對種子活力的影響。張詩慧等[10]以白刺花種子為材料,采用高溫、高濕人工加速老化的方法探究種子貯藏過程中生理生化指標及種子活力變化,發現老化處理6d的白刺花種子發芽勢、發芽指數、活力指數和SOD活性顯著升高(p<0.05),丙二醛、可溶性蛋白含量顯著下降(p<0.05);幼苗長勢以處理2d最優。祝煜中等[11]以甜玉米品種“農甜 88”和“農甜 99”為試驗材料,研究了人工老化處理對甜玉米種子活力、生理特性和遺傳多樣性的影響,發現隨老化時間的增加,2個甜玉米品種種子的活力以及DHA、POD活性逐漸降低,REC和MDA含量逐漸升高;種子老化處理降低了種子遺傳物質的多樣性。目前,有關水稻種子人工老化的研究報道較少。基于此,本試驗采用人工老化處理方法,研究不同老化時間對水稻種子活力及生理生化特性的影響,希望能夠為延緩種子老化進程和延長種子壽命提供理論依據。
1 材料與方法
1.1 試驗材料
試驗用水稻種子為稻花香1號,2020年11月購自當地種子公司。購買來的種子在4℃冰箱內儲存,5個月后進行人工老化試驗。在人工老化處理前,將種子置于室溫下平衡24h。
1.2 試驗方法
1.2.1 種子處理
在相對濕度為95%、溫度為40℃的高溫、高濕環境內對水稻種子進行人工老化。選取干凈飽滿的水稻種子置于網紗袋內,均勻放置于已平衡好的老化箱內,分別對種子處理1-8d,分期取出種子后,測量種子發芽指標、幼苗形態指標及生理生化指標。將貯藏于-20℃冰箱內未經老化處理的種子作為對照處理。
1.2.2 發芽試驗
隨機選擇經人工老化處理的水稻種子,置于45℃溫水內浸種12h后置于發芽盒(蛭石基質)內進行發芽試驗,每個處理設置3個重復,每個重復50粒種子,于人工培養箱內培養(溫度為25℃,每天光照時間為8h)。在發芽試驗開始后,每日透氣30min,及時為種子補入充足的水分。21d后結束發芽試驗,注意將發霉種子剔除,觀察發芽種子數。
1.2.3 測量指標及方法
(1)發芽指標
在發芽試驗結束后,按照下式計算水稻種子發芽率、發芽指數、活力指數。
發芽率Gr(%)=(ΣGt/NT)×100%
發芽勢Ge(%)=高峰時發芽種子數/種子總粒數×100%
發芽指數Gi=Σ(Gt/Dt)
活力指數Vi=S×Gi
上式中,Gt為在第t天發芽的種子數;NT為50(每個重復種子總數);S為幼苗平均干質量。
(2)幼苗形態指標
在發芽試驗結束后,每個重復隨機選擇10株幼苗(老化時間較長時,幼苗萌發數量較少,以實際測量株數為準進行形態指標測量),采用直尺測量水稻幼苗苗高、根長等指標。
(3)生理生化指標
各處理分別采用老化處理后的水稻種子50粒,參照屈煜瑩等[12]的方法測量水稻種子相對電導率;采用蒽酮法[13]測量可溶性糖含量;硫代巴比妥酸法[14]測量丙二醛含量;考馬斯亮藍法測量可溶性蛋白含量;參照蔡慶生主編的《植物生理學實驗》[15]一書中的方法測量超氧化物歧化酶(SOD)、過氧化物酶(POD)和過氧化氫酶(CAT)活性。
1.3 數據處理
采用Excel 2007軟件對試驗數據進行整理,SPSS20.0軟件對試驗數據進行統計分析。
2 結果與分析
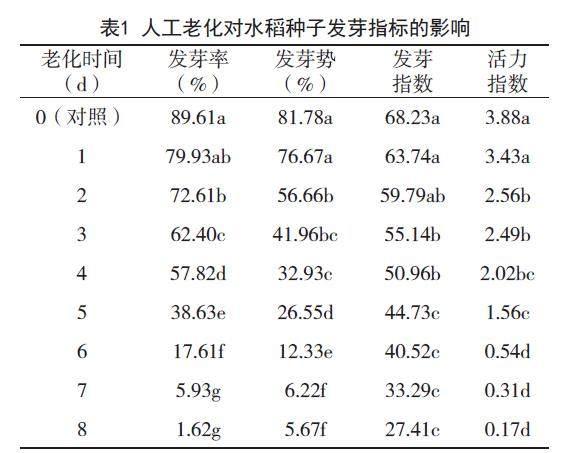
2.1 人工老化對水稻種子發芽指標的影響
不同人工老化時間對水稻種子各發芽指標的影響,見表1。
由表1可知,在人工老化條件下,水稻種子發芽率、發芽勢、發芽指數、活力指數均呈現出逐漸降低趨勢。在老化初期(即老化1-4天時間內),水稻種子發芽率降低較為緩慢,由0d的89.61%逐漸降低至老化4d的57.82%,隨著老化時間的進一步延長,發芽率快速降低,在老化8d時水稻種子發芽率僅1.62%,其中,老化0d和老化1d水稻種子發芽率差異不顯著,老化1d和老化2d水稻種子發芽率差異不顯著,老化7d和老化8d水稻種子發芽率差異不顯著;從水稻種子發芽勢情況來看,在老化1d時,水稻種子發芽勢與0d時無明顯差異,這表明老化1d對水稻種子的發芽勢不存在顯著影響,隨著老化時間的逐漸延長,水稻種子發芽勢開始降低,由76.67%降低至5.67%;在本試驗條件下,水稻種子發芽指數在27.41%-68.23%之間,其中,老化0d時水稻種子發芽指數最高,而老化8d時水稻種子發芽指數最低,老化0d、1d、2d時水稻種子發芽指數差異不顯著,老化2d、3d、4d時水稻種子發芽指數差異不顯著,老化5d、6d、7d、8d時水稻種子發芽指數差異不顯著;從不同老化時間下水稻種子活力指數情況來看,各老化處理水稻種子活力指數均低于未老化處理,相較于老化0d,老化1d、2d、3d、4d、5d、6d、7d、8d水稻種子活力指數分別降低了11.60%、34.02%、35.82%、47.94%、59.79%、86.08%、92.01%、95.62%。
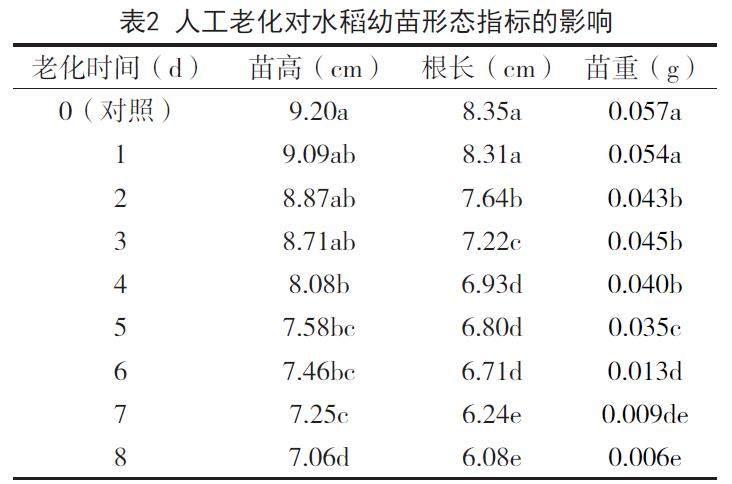
2.2 人工老化對水稻幼苗形態指標的影響
人工老化對水稻幼苗形態指標的影響,見表2。
由表2可知,隨著人工老化時間的延長,水稻幼苗苗高呈逐漸降低趨勢,由老化0d的9.20cm逐漸降低至老化8d的7.06cm,其中,老化0d、1d、2d、3d處理水稻苗高差異不顯著,老化1d、2d、3d、4d、5d、6d處理水稻苗高差異不顯著,老化5d、6d、7d水稻苗高差異不顯著;從水稻幼苗根長情況來看,在本試驗條件下各處理水稻幼苗根長在6.08-8.38cm之間,以老化0d處理水稻根長為最長,老化8d水稻幼苗根長為最小,老化1d處理水稻幼苗根長與老化0d處理差異不顯著,老化7d處理水稻幼苗根長與老化8d處理差異不顯著;從水稻幼苗苗重情況來看,在老化初期,水稻幼苗苗重呈緩慢降低趨勢,由0.057g逐漸降低至0.035g,其中,老化0d處理與老化1d處理水稻幼苗苗重差異不顯著,老化2d、老化3d、老化4d處理水稻幼苗苗重差異不顯著,從老化5d到老化6d,水稻幼苗苗重顯著降低,由0.035g降低至0.013g。
2.3 人工老化對水稻幼苗生理生化指標的影響
人工老化對水稻滲透調節物質含量的影響,見表3。
由表3可知,不同老化時間下水稻種子電導率、可溶性糖含量、丙二醛含量差異顯著,可溶性蛋白質含量差異不顯著。具體而言,隨著老化時間的增加,水稻種子相對電導率、丙二醛含量呈現出逐漸增長的趨勢,可溶性糖含量呈現出逐漸降低趨勢。從相對電導率情況來看,老化7d處理最高,達到了65.66%,但與老化8d水稻種子相對電導率差異不顯著;老化0d處理最低,僅49.75%,與老化1d、老化2d處理差異不顯著,其余處理水稻種子相對電導率居中。從水稻種子可溶性糖含量情況來看,在老化時間小于2d內時,水稻種子可溶性糖含量差異不顯著,維持在9.35-9.72mg/g之間,隨著老化時間的延長,可溶性糖含量有所降低,老化時間在3-5d時,水稻種子可溶性糖含量維持在8.33-8.62mg/g之間,三者差異不顯著。在老化時間為6-8d時,水稻種子可溶性糖含量差異不顯著,維持在7.03-7.40mg/g之間。在本試驗條件下,水稻種子可溶性蛋白質含量維持在4.33-5.83mg/g之間,這表明老化時間對水稻種子可溶性蛋白質含量不存在顯著影響。從水稻種子丙二醛含量情況來看,隨著老化時間的延長,丙二醛含量明顯增加,老化8d、7d、6d、5d、4d、3d、2d、1d處理水稻種子丙二醛含量分別較對照增加了70.43%、64.79%、56.33%、52.43%、37.97%、31.19%、15.66%、3.96%,其中,老化0d、老化1d處理水稻種子丙二醛含量差異不顯著,老化2d、3d、4d、5d處理水稻種子丙二醛含量差異不顯著,老化4d、5d、6d、7d、8d處理水稻種子丙二醛含量差異不顯著。
2.4 人工老化對水稻種子酶活性的影響
不同人工老化時間對水稻種子酶活性的影響,見表4。
由表4可知,不同老化時間對水稻種子SOD活性、POD活性、CAT活性均存在顯著影響。具體而言,隨著老化時間的延長,水稻種子SOD活性呈現出先升高、后降低的趨勢,由老化0d處理的175.37U/g逐漸升高至老化4d處理的275.79U/g,老化時間的進一步增加,不會導致SOD活性的進一步增加,反而會導致SOD活性的降低,最終SOD活性降低為老化8d處理的180.78U/g,其中,老化0d、1d、2d、8d處理水稻種子SOD活性差異不顯著,老化3d、4d、5d、6d處理水稻種子SOD活性差異不顯著,老化3d、5d、6d、7d處理水稻種子SOD活性差異不顯著;從水稻種子POD活性情況來看,其由高到低排序依次為老化3d、2d、1d、0d、4d、5d、6d、7d、8d,其中,老化3d處理水稻種子POD活性為126.35U/g,老化8d處理水稻種子POD活性僅64.05U/g,老化3d、2d、1d、0d四個處理水稻種子POD活性差異不顯著,老化1d、0d、4d處理水稻種子POD活性差異不顯著,老化5d、6d、7d、8d處理水稻種子POD活性差異不顯著;隨著老化時間的延長,水稻種子CAT活性呈逐漸降低趨勢,老化1d、2d、3d、4d、5d、6d、7d、8d處理水稻CAT活性分別較老化0d處理降低了97.94%、93.04%、91.56%、82.01%、73.67%、66.28%、64.35%、55.29%,其中,老化0d、1d、2d、3d四個處理水稻種子CAT活性差異不顯著。
3 討論
在種子生理成熟后,其活力開始降低,種子萌發狀況是種子活力水平的重要反饋指標之一。本研究發現,在老化過程中,水稻種子發芽率、發芽勢、發芽指數及活力指數均呈現出逐漸降低的趨勢,這與朱世東等[16]的研究結果相一致,這表明在經過人工老化后水稻種子的活力有所衰退。
在經過老化處理后,不同作物種子其幼苗所發生的變化存在著一定差異。王玉嬌等[17]發現,在對春小麥種子進行老化處理后,幼苗的生長發育有所延緩,同時幼苗根系受到傷害;何學青等[18]發現,老化對柳枝稷幼苗的生長沒有顯著影響,同時可提升幼苗根冠比。本研究發現,經過老化處理的水稻幼苗發生了變化。老化時間較長時,水稻幼苗苗長、根長、苗重均小于對照,這說明水稻種子在老化后,其幼苗生長受到了一定影響。
本研究發現,隨著老化時間的增加,水稻種子相對電導率、丙二醛含量呈現出逐漸增長的趨勢,可溶性糖含量呈現出逐漸降低趨勢。這主要是由于在種子老化過程中,細胞膜極容易受到活性氧的攻擊而造成脂質過氧化反應,這會導致細胞膜受損,同時產生大量的脂質自由基及有毒產物丙二醛,丙二醛能夠與抗氧化酶相結合以降低抗氧化酶活性,導致消除自由基能力的減弱,因自由基的積累而導致生物膜受損的進一步加速,致使大量膜內無機物質及礦物質外滲,最終造成浸出液電導率的升高[19]。
SOD、POD、CAT是植物細胞重要的抗氧化酶,抗氧化酶可消除自由基以及過氧化物,使自由基的產生及消除處于平衡狀態。本研究發現,隨著老化時間的延長,SOD活性及POD活性呈現出先升高、后降低的趨勢,這可能是由于在高溫、高濕條件下,種子產生了大量的自由基,SOD、POD產生了應激反應,通過迅速提升活性以清除自由基,隨著老化時間的延長,大量活性氧產生,抗氧化酶無法及時清除活性氧,導致細胞內過氧化物的產生及消除平衡遭到破壞,最終SOD、POD活性開始降低。
4 結論
綜上所述,在短期高溫高濕老化處理下(1d以內),水稻種子活力指標無明顯變化。隨著老化時間的延長,水稻種子活力不斷降低,相對電導率、丙二醛含量升高,可溶性糖含量降低,SOD活性、POD活性先升高后降低,CAT活性持續降低。因此,在高溫、高濕條件下,人工老化1天不會對水稻種子造成影響。
參考文獻
[1] 金文玲,曹乃亮,朱明東,等.基于近紅外超連續激光光譜的水稻種子活力無損分級檢測研究[J].中國光學,2020,13(5):1032-1043.
[2] 楊翠,田宏躍,覃廷英,等.不同類型水稻種子活力的比較研究[J].種子,2019,38(11):12-15.
[3] 秦潔.水稻種子活力研究進展及工作展望[J].種子科技,2021,39(13):15-16.
[4] 余亞瑩,段永紅,唐瀟,等.人工老化處理對水稻兩系不育系及其雜交組合種子活力的影響[J].雜交水稻,2021,36(6):64-69.
[5] 高華偉,滿強,潘晶,等.大豆人工老化與自然老化的種子活力差異研究[J].種子,2015,34(1):14-18.
[6] 劉明久,王鐵固,陳士林,等.玉米種子人工老化過程中生理特性與種子活力的變化[J].核農學報,2008,22(4):510-513.
[7] 吳聚蘭,周小梅,范玲娟,等.人工老化對大豆種子活力和生理生化特性的影響[J].中國油料作物學報,2011(6):582-587.
[8] 許惠濱,魏毅東,連玲,等.水稻種子人工老化與自然老化的分析比較[J].分子植物育種,2013,11(5):552-556.
[9] 曾大力,錢前,國廣泰史,等.稻谷儲藏特性及其與秈粳特性的關系研究[J].作物學報,2002,28(4):551-554.
[10] 張詩慧,吳文榮,歐順,等.不同老化處理對‘白刺花種子活力的影響[J].熱帶農業科學,2020,40(6):16-22.
[11] 祝煜中,夏黎明,竺思儀,等.人工老化的甜玉米種子活力、生理特性和遺傳多樣性的變化[J].華南農業大學學報,2018,39(1): 25-30.
[12] 屈煜瑩,黃鑫,徐強輝,等.人工老化處理對芥藍種子生理生化特性的影響[J].種子,2020,39(8):24-29.
[13] 艾希珍,于賢昌,王紹輝,等.低溫脅迫下黃瓜嫁接苗與自根苗某些物質含量的變化(簡報)[J].植物生理學通訊,1999,35(1):26-28.
[14] 趙世杰,許長成,鄒琦,等.植物組織中丙二醛測定方法的改進[J].植物生理學通訊,1994(3):207-210.
[15] 蔡慶生.植物生理學實驗[M].北京:中國農業大學出版社,2013.
[16] 朱世東,斐孝伯,張志偉.香椿種子衰老機理[J].安徽農業科學,1999,27(1):62-63.
[17] 王玉嬌,吳薇,郭忠軍,等.小麥種子老化處理對發芽指標及根系的影響[J].核農學報,2018,32(12):2423-2430.
[18] 何學青,陶奇波,沙亞·海拉提,等.柳枝稷種子活力及幼苗生長對人工老化處理的響應[J].草地學報,2016,24(4):834-840.
[19] 張海波,楊桂娟,高衛東,等.香椿種子特定貯藏條件下活力變化的研究[J].林業科學研究,2019,32(2):152-159.
習近平《 論“三農”工作》
把鄉村振興戰略這篇大文章做好⑨
(二〇一八年九月二十一日)
要始終把解決好“三農”問題作為全黨工作重中之重。我們一直強調,對“三農“要多予少取放活,但實際工作中“三農”工作“說起來重要、千起來次要、忙起來不要”的問題還比較突出。我們要扭轉這種傾向,在資金投入、要素配置、公共服務、干部配備等方面采取有力舉措,加快補齊農業農村發展短板,不斷縮小城鄉差距,讓農業成為有奔頭的產業,讓農民成為有吸引力的職業,讓農村成為安居樂業的家園。
——節選自習近平同志主持中共十九屆中央政治局第八次集體學習時講話,《論“三農”工作》,中央文獻出版社,2022年6月第1版
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
家庭影院技術(2020年10期)2020-12-14 07:54:18
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
知識經濟·中國直銷(2016年3期)2016-02-27 16:15:49
現代檢驗醫學雜志(2014年6期)2014-02-02 03:02:04
閱讀與作文(小學低年級版)(2011年3期)2011-01-01 00:00:00
