
中國近海漁業資源的結構與質量研究
2024-07-13 04:48:59張瑛李忠輝張力
中國人口·資源與環境 2024年5期
張瑛 李忠輝 張力

摘要 海洋漁業資源對于保障食品安全、維護生態系統穩定、促進經濟發展等方面起著關鍵作用,科學管理和可持續利用近海漁業資源,對于推動經濟與環境協調發展至關重要。為了探究中國近海漁業資源的結構與質量變化情況,該研究基于1990—2020年中國近海捕撈統計數據和海表溫度(sea surface temperature, SST)變化數據,通過計算分析中國近海各漁區(渤黃海漁區、東海漁區和南海漁區)漁獲物的平均營養級(mean trophic level,MTL)、各漁區漁業均衡指數(fishing?in?balance index,FIB指數)與人為捕撈活動、SST變化之間的關聯性,探究人為捕撈活動、SST變化對中國近海漁業資源結構與質量變化、漁業生態系統健康的影響。結果表明:過去30年間,中國近海各漁區的漁獲量和FIB指數均呈現先上升后下降的趨勢,渤黃海和東海漁區的MTL呈現顯著下降趨勢,南海漁區的MTL呈現微弱增長趨勢。人為過度捕撈已影響到渤黃海和東海漁區的漁業資源結構與質量,導致低營養級魚類占比逐漸升高,漁業資源質量顯著降低,但對南海漁區影響不大。受到氣候變化影響,在渤黃海漁區5—6月和東海漁區9—11月,海溫增暖抑制高營養級魚類生長,導致漁業資源出現結構向低營養級魚類變化、質量大幅下降的趨勢,但在南海漁區,海溫變化對MTL沒有影響。人為捕撈對中國近海漁業生態系統健康影響不大,主要是由于漁獲量的增加起到了一定的平衡作用。海溫變化對渤黃海、東海漁區的漁業生態系統健康影響也較小,南海秋季海溫增暖則有利于其漁業生態系統健康發展。基于研究結果,作者提出了改善中國近海各漁區漁業資源結構,提高漁業資源質量,提升漁業生態系統健康的相關建議,為中國近海漁業資源的可持續利用提供了管理借鑒。
關鍵詞 近海;漁業資源;人為捕撈;氣候變化
中圖分類號 P74;S931 文獻標志碼 A 文章編號 1002-2104(2024)05-0058-11 DOI:10. 12062/cpre. 20231021
海洋漁業資源是一種重要的自然資源,為人類提供了高質量的食物來源,并且為將近70%的世界人口提供了40%的蛋白質[1]。海洋漁業資源種類繁多,包括魚類、甲殼類、貝類、藻類、足類和其他類,其中魚類和甲殼類的品種眾多,數量大。海洋漁業資源的合理利用對維護海洋資源的生態平衡、推動海洋經濟朝著健康可持續的方向發展有著舉足輕重的意義[2-3]。中國大陸海岸線綿長,同時擁有渤海、黃海、東海和南海等邊緣海,海洋漁業資源豐富。但從20世紀中期開始,由于市場上對高質量漁業資源的需求不斷擴大,許多先進的捕撈技術也就應運而生,捕撈力度不斷加大,加之氣候變化、海洋環境污染、管理制度等原因,中國近海漁業資源可持續利用面臨巨大的挑戰[4-9]。因此,對海洋漁業資源動態變化的有效掌握,將有利于及時發現其開發利用過程中出現的問題,推動中國海洋漁業健康發展[10]。本研究聚焦人為捕撈和氣候變化雙重影響,對中國近海漁業資源結構與質量變化進行對比分析與歸因研究,以期為中國近海漁業資源高質量管理提供決策參考。
1 文獻綜述
近海漁業資源是海洋漁業資源重要的組成部分。近10年,中國近海捕撈漁業迅速發展的同時,也導致了其與資源環境沖突問題愈來愈嚴重。一方面,由于人為影響,隨著捕撈技術的提升和捕撈政策的開放,捕撈強度呈上升趨勢,且居高不下,致使近海捕撈長期處于飽和的危險狀態。同時,捕撈強度增加使得近海漁業資源衰退嚴重,近海主要經濟魚類資源大幅衰減[4,7]。另一方面,氣候變化對漁業資源波動也影響深遠。研究表明,氣候變化主要通過改變海水溫度、上層洋流、海洋層結等過程,影響海水酸堿度、營養鹽分布等狀況,進一步對漁業資源的多時空尺度變化產生影響[8,11]。
針對目前的海洋漁業資源利用現狀,國內外學者從不同的方向進行了研究[12-18]。已有研究在影響因素的考量方面,就人為捕撈因素對漁業資源可持續利用產生的影響,從宏觀系統角度開展了較多研究。劉子飛[14-15]分析了中國近海捕撈漁業管理政策對漁業資源狀態的影響,指出隨著專業捕撈人員、捕撈漁船功率噸位及非選擇性漁具的不斷增加,中國近海漁業資源減少是很難避免的,不合理的漁具使用和超強度的捕撈會破壞漁業資源生存的“三場一通道”,對漁業資源的可持續利用形成負面影響。這些研究從宏觀角度揭示了高強度的人為捕撈導致了中國近海漁業資源減少的現象,但尚未就人為捕撈對中國近海漁業資源結構與質量進行量化分析。
為衡量漁業資源結構與質量變化,國內外相關研究中廣泛使用漁獲物營養層級變化作為重要的參考指標[2]。漁獲物平均營養級(mean trophic level,MTL)主要用來反映漁獲物種群結構的變化情況[19-20]。當高營養級的捕食性魚類被過度捕撈時,平均營養級會降低,可能會導致整個漁業生態系統平衡被打破。除MTL 外,Pauly等[21]又提出漁業均衡指數(Fishing?in?Balance index,FIB指數),主要聚焦食物鏈中底層的小型食餌魚類的生物量。這些小型生物是大型捕食性魚類的主要食物來源,過度捕撈會影響到上層食物鏈的魚類。MTL和FIB指數結合可以更全面反映漁業生態系統中不同營養級的動態變化[22-24]。國內外學者針對特定海域漁獲物營養級變化開展了卓有成效的研究。例如,吳佳穎等[12]對渤黃海漁區主要漁獲物的平均營養級進行了研究,指出渤黃海MTL在近50年間整體呈下降趨勢,中低營養級漁獲物的漁獲量增加,而高營養級漁獲物的漁獲量減少。李繼龍等[13]根據中國漁船漁獲統計數據,對中國東部海域和南部海域的漁業資源利用水平進行了研究,指出東部海域MTL在近30年間呈現下降趨勢,其漁業資源利用已經呈現出一種不可持續性,而南部海域MTL無顯著變化趨勢,并未出現漁業資源利用的衰退現象。Milessi等[25]聚焦烏拉圭海域,通過分析烏拉圭漁場60種漁獲物的營養級,發現MTL以每10年0. 28的速度下降,烏拉圭漁場漁業資源利用呈現出不可持續性。
除了人類的捕撈活動外,氣候變化是影響漁業資源整體結構與質量的另一個關鍵因素。氣候變化對MTL變化的影響存在持續性,其關鍵影響路徑是通過改變海洋環境要素進而對漁業資源結構與質量產生影響。海溫與海洋物種的分布、豐度及其變化有著顯著的聯系,適宜的海溫有利于海洋生物的新陳代謝,促進海洋生物的生長和繁殖[26],異常的海溫會影響海洋生物的生理功能,導致新陳代謝紊亂,從而影響其生長發育[27]。而海洋生物的生長發育又影響著自身的體長,一般來說,海洋生物的體長與自身營養級之間存在著聯系[28-29]。因此,氣候變化通過影響海溫,進而對海洋生物的體長和營養級產生影響。陳爽等[30]針對東北大西洋海域的MTL和多樣性指數進行了探究,分析結果表明,這兩個指標與海洋環境因子的變化有著顯著聯系,隨著海溫和海水鹽度不斷升高,多樣性指數和MTL呈現下降趨勢。另外,海溫變化與海洋生物的資源量變化,以及群落格局的代際變化也有聯系[31-32]。基于氣候變化的不斷加劇,其所帶來的海洋環境變化對漁業資源的影響是漁業管理重要的研究方向之一[33]。對于中國近海,海溫呈現快速增暖趨勢,其速率大約是全球平均的3~5倍[11],這種快速增溫如何影響漁業資源結構與質量亟待評估。
本研究圍繞人為捕撈和氣候變化對中國近海漁業資源結構與質量變化的雙重影響,利用1990—2020年的國內外相關數據系統分析中國近海3個漁區(渤黃海漁區、東海漁區、南海漁區)的MTL和FIB指數的變化趨勢,重點分析捕撈漁船不同作業類型、海溫變化對各漁區MTL和FIB指數的影響,進一步對各漁區的漁業資源結構與質量變化進行對比分析與歸因研究。本研究中近海漁業資源主要考慮海水中具有經濟價值的動物,以魚類和甲殼類為主。
2 數據來源與研究方法
2. 1 數據來源
本研究所采用的渤黃海、東海、南海1990—2020年的漁獲量數據,包括按品種和漁具分類的各地區海洋捕撈產量、按海域劃分的各海域海洋捕撈產量均來自《中國漁業統計年鑒》國內海洋捕撈生產統計。結合數據的可獲取性,分別利用遼寧省、河北省、天津市、山東省、江蘇省的漁獲統計數據對渤黃海漁區的漁獲物進行分析;利用浙江省、上海市、福建省的漁獲統計數據對東海漁區的漁獲物進行分析;利用廣東省、廣西壯族自治區、海南省的漁獲統計數據對南海漁區的漁獲物進行分析。上述漁獲物的種類主要包括鳀魚、小黃魚、鮐魚、梭子蟹、鷹爪蝦等26種。相關漁獲物種類營養級的確定主要借鑒孫明等[34]對遼東灣海域主要漁業生物營養級的研究,白懷宇等[35]對黃海主要魚類營養級和食性的研究,高春霞等[36]對浙江南部近海主要漁業生物營養級的研究,黃佳興等[29]對南海中西部漁場主要漁業生物營養級的研究。
中國近海SST數據來自美國國家海洋和大氣管理局(NOAA),水平分辨率為1°×1°;水平覆蓋范圍為105°E~131°E,4°N~40°N;覆蓋時段為1990 年1 月—2020 年12月。
2. 2 研究方法
與個體小、壽命短、營養級低的漁業生物相比,個體大、壽命長、營養級高的肉食性漁業生物更容易受到過度捕撈的影響,這會使得海域內整體漁業資源的營養級水平呈下降趨勢,而MTL指標可以用來確定某漁區內高營養級漁業資源的衰退情況。同時,FIB指數可以與MTL指標相結合,從而更加全面地評估漁業生態系統的均衡狀況。另外,為了確定人為捕撈活動和海溫變化對MTL和FIB指數的具體影響,本研究采取一元相關分析方法進行計算分析。
2. 2. 1 計算漁獲物平均營養級
漁獲物平均營養級概念是由Pauly等[20]首先提出的,主要用來反映漁獲物種群結構的變化情況,判斷多物種漁業生物的承受力,以衡量漁業生物多樣性和可持續性水平,其計算公式為:
另外,借鑒鄧景耀等[37]對中國近海漁獲物營養等級劃分的研究,本研究依據營養級指數將漁獲物營養等級分為3 級,即低營養級種類(2. 0~2. 8)、中營養級種類(2. 9~3. 4)和高營養級種類(3. 5~4. 0)。這種劃分有助于觀測海洋生物營養層級構成變化,高營養級種類漁獲量比重的下降,可以幫助確定某海域是否出現了捕撈對象沿著海洋食物網向下移動的現象。
2. 2. 2 計算漁業均衡指數
Pauly等[21]認為,捕撈活動對于漁業生態系統的影響是多方面的,只計算MTL這一個指標的變化,可能會存在“刻意降低食物網”的現象。因此,他又提出了另一個指標,即FIB指數,該指數是用來衡量相對于基準年的總捕撈量與MTL的抵消情況。FIB指數意在考慮漁業轉向捕撈低營養級物種時可能出現的捕撈量增加的情況,其計算公式為:
2. 2. 3 一元相關分析
一元相關分析是一種用于研究兩個連續型變量之間關系的統計方法,以便于了解變量之間的趨勢和相關程度。在這個分析中,主要關注皮爾遜相關系數,該系數的取值范圍介于-1到1之間,為正值時表示正相關,負值時表示負相關,而接近于0則意味著較弱的相關性。具體計算公式如下:
為研究人為捕撈活動對漁業資源變化的影響,本研究將統計范圍內捕撈漁船作業類型劃分為5種,包括選擇性漁具(圍網、釣具和流刺網)和非選擇性漁具(拖網和定置網),將捕撈漁船不同作業類型的漁獲量分別與MTL、各營養等級漁獲量占比、FIB指數進行一元相關分析。為研究氣候變化對漁業資源變化的影響,本研究著重分析中國近海各漁區每個月SST變化趨勢對MTL和FIB指數的影響。
3 結果分析
3. 1 中國近海各漁區漁獲量、漁獲物平均營養級與漁業均衡指數的變化
就漁獲量而言,1990—2020年渤黃海和東海漁區的年漁獲量呈現基本相同的變化趨勢,具體表現為在1990—1998年期間增長迅速,達到歷史最高漁獲量,之后基本保持穩定,2017年開始下降。南海漁區的漁獲量在1990—2005年持續增長,2006—2016年漁獲量穩定,之后逐年下降。綜合30年間的漁獲總量,東海最高,渤黃海次之,南海最低(圖1(a))。
就MTL而言,渤黃海漁區的MTL在1990—2002年呈現整體下降趨勢,2003—2007年則呈現上升趨勢,2007—2010年又逐漸下降,之后穩定在3. 04左右。東海漁區的MTL 在1990—2002 年呈現整體下降趨勢,之后穩定在3. 27左右。南海漁區的MTL在1990—2003年呈現微弱增長趨勢,2004—2006年逐漸上升,之后穩定在3. 2左右(圖1(b))。
就FIB指數而言,渤黃海和東海漁區的FIB指數整體上呈現出了先上升后下降的趨勢。具體來說,1990—1998年FIB指數呈現上升趨勢,1999—2020年FIB指數呈現下降趨勢。在南海漁區,FIB指數總體上呈現上升趨勢,2017年后有所下降(圖1(c))。
從各營養等級漁獲量占比來看,在渤黃海漁區,低營養級漁獲量占比最高,基本穩定在40%~50%;中營養級漁獲量占比整體呈現上升趨勢,在2006年后穩定在33%左右;高營養級漁獲量占比呈現下降趨勢,由1990年的34%下降至2020年的20%(圖2(a))。在東海漁區,高營養級漁獲量占比最高,呈現顯著的下降趨勢,由1900—2001年的52%迅速下降到2002—2020年的40%;中營養級漁獲量占比整體呈現顯著的上升趨勢,由1990—2000年的30%上升到2003—2020年的41%;低營養級漁獲量占比最小,基本上穩定在20%左右(圖2(b))。在南海漁區,中營養級漁獲量占比最高,在1998年前呈現下降趨勢,之后穩定在40%;低營養級漁獲量占比整體穩定在20%左右;高營養級漁獲量占比呈現上升趨勢,由1990年的20%上升到2020年的35%(圖2(c))。
為進一步分析漁獲量與MTL、漁獲量與FIB指數之間的關系,本研究分別計算了不同漁區漁獲量與MTL、FIB指數的相關系數(表1)。結果顯示,渤黃海漁區的漁獲量與MTL相關系數為-0. 75,呈現顯著的負相關關系;東海漁區的漁獲量與MTL相關系數為-0. 34,統計學上依然顯著。以上結果表明,在渤黃海和東海漁區,人為捕撈對漁業資源的結構與質量變化有顯著的影響,過度捕撈會帶來營養級降低。南海漁區的漁獲量與MTL無相關關系,這表明人為捕撈未對漁業資源的結構與質量產生顯著的影響。從FIB指數來看,3個漁區的漁獲量與FIB指數均呈現顯著的正相關關系,這表明漁獲量的增加能夠抵消MTL降低帶來的影響,從而導致FIB指數上升。漁獲量與MTL、FIB指數之間的相互影響,使得漁業生態系統在結構和功能上保持著相對良好的健康狀態。
3. 2 中國近海捕撈漁船不同作業類型對MTL及FIB指數的影響
圖3 展示了1990—2020 年中國近海不同作業類型的漁獲量變化情況。由圖3可以看出,在3個漁區,不同作業類型中拖網的漁獲量均是最多,但渤黃海和東海漁區拖網的漁獲量要遠遠大于南海漁區拖網的漁獲量。在渤黃海漁區,流刺網和定置網的漁獲量大致相當,圍網和釣具的漁獲量則相對較少;在東海漁區,定置網的漁獲量緊隨在拖網的漁獲量之后,流刺網、圍網和釣具的漁獲量則相對較低;在南海漁區,流刺網的漁獲量僅次于拖網的漁獲量,并且流刺網的漁獲量占比在3個漁區中最高。
各作業類型的漁獲量主要是由漁船的數量、噸位、功率所決定。本研究以2002、2011、2020年為例,分析上述因素對不同作業類型漁獲量的影響程度。由圖4可以看出,在渤黃海漁區,漁船的數量主要以流刺網和拖網為主,其中流刺網占比大于拖網;在漁船的噸位和功率方面,拖網占比遠超過流刺網。在東海漁區,漁船的數量以拖網、流刺網和定置網為主,三者所占比例相差不大;在漁船的噸位和功率方面,拖網占比遠大于其他4種作業類型占比,約占55%。在南海漁區,流刺網在漁船的數量中占比最大,高達70%左右;在漁船的噸位和功率方面,主要以拖網和流刺網為主,其中拖網占比稍高于流刺網。
為分析各漁區不同作業類型對漁業資源結構與質量的影響,本研究分別計算了不同作業類型的漁獲量與MTL、各營養等級漁獲量占比、FIB指數的相關系數(表2—表4。由表2可知,在渤黃海漁區,拖網漁船漁獲量與低營養級漁獲量占比呈現顯著的正相關關系,與高營養級漁獲量占比呈現顯著的負相關關系。這表明,隨著拖網漁船作業類型的漁獲量增加,渤黃海漁區的高營養級漁獲量占比將減少,而低營養級漁獲量占比將增加,這是渤黃海漁區的MTL呈現下降趨勢的原因之一(圖1(b))。由表3可知,在東海漁區,拖網漁船漁獲量與中營養級漁獲量占比呈現顯著的正相關關系,與高營養級漁獲量占比呈現顯著的負相關關系。這表明,隨著拖網漁船作業類型的漁獲量增加,東海漁區的中營養級漁獲量占比將增加,而高營養級漁獲量占比將會減少,這是東海漁區的MTL呈現下降趨勢的原因之一(圖1(b))。由表4可知,在南海漁區,拖網的漁獲量雖然最高,但與高營養級漁獲量占比下降的相關關系不顯著;流刺網的漁獲量與高營養級漁獲量占比呈現顯著的正相關關系。這表明,隨著流刺網作業類型的漁獲量增加,南海漁區的中、高營養級漁獲量占比將增加,這是南海漁區的MTL呈現上升趨勢的原因之一(圖1(b))。
就FIB指數而言,渤黃海與東海漁區的漁獲量中占比最大的拖網漁船與FIB指數呈現正相關關系,但南海漁區兩者的相關關系卻不顯著。在南海漁區,流刺網的漁獲量與FIB指數呈現顯著的正相關關系。這表明,3個漁區主要作業類型的漁獲量增加能抵消MTL 下降的負面效應,以維持健康的漁業生態系統結構與功能。
3. 3 海溫變化對MTL及FIB指數的影響
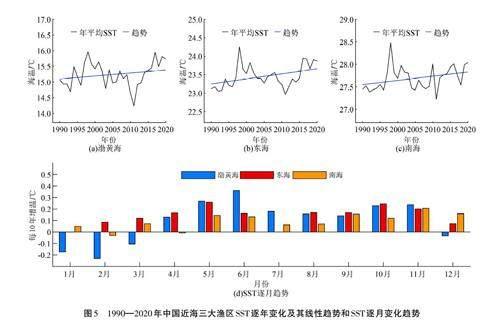
由圖5可以看出,中國近海各漁區SST整體均呈上升趨勢,其中東海增溫最為顯著,南海較弱。1998年,3個漁區的SST均達到最高值,是由于1997/1998年厄爾尼諾現象異常強烈,厄爾尼諾現象發生期間會導致中國近海的SST異常增暖,尤其是在厄爾尼諾現象發生的冬季和春季,對中國近海的SST升高影響最為顯著[39]。2011—2012年,3個漁區的SST均達到最低值,其原因在于2011年拉尼娜現象強度較往年大[40],拉尼娜現象發生期間會導致中國近海的SST異常下降。2011年之后,中國近海各漁區SST上升趨勢增加。南海漁區SST變化相對較弱是由于南海漁區海水溫度和鹽度的垂向結構變化非常復雜[41],同時受到南海季風[42]、黑潮入侵[43]、印度洋偶極子[44]等多因素復雜影響,因此南海漁區SST變化有一定的獨特性[45]。
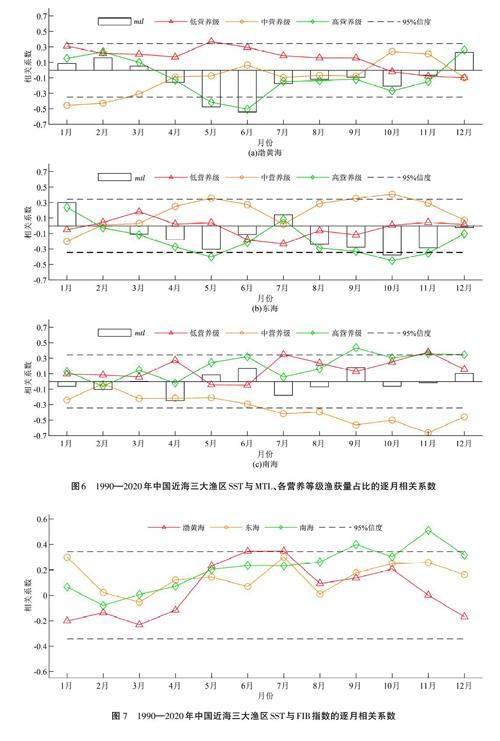
SST變化會影響到海洋生物的生長和發育[46-48],進而影響到營養級的變化[ 28]。在渤黃海漁區,SST與MTL整體上呈現負相關關系,這種關系主要由5—6月的SST與高營養級魚類呈現顯著的負相關關系所決定(圖6(a))。就增溫趨勢而言,渤黃海漁區增溫趨勢最為顯著的也是在5—6月,分別達到每10年0. 27 oC和0. 36 oC(圖5(d)),從而抑制了同期高營養級魚類的生長,這是渤黃海漁區的MTL呈現下降趨勢的原因之一。在東海漁區,SST與MTL整體上呈現負相關關系,主要是由于秋季海溫下降抑制了高營養級魚類的生長,以9—11月最為顯著;同時,SST與中營養級魚類呈現正相關關系,一定程度上削弱了MTL的下降趨勢(圖6(b)),也表明東海漁區有中營養級魚類的生長期在秋季。從圖5(d)可以看出,東海漁區秋季增暖趨勢也比較明顯,基本上在每10年0. 17 oC~0. 24oC,表明東海漁區秋季增暖趨勢抑制了高營養級魚類的生長,從而對其MTL下降趨勢有一定的貢獻度。在南海漁區,SST變化與MTL整體上沒有呈現顯著的相關關系,但各營養級魚類呈現不同的響應,尤其是在7—12月(圖6(c)),SST與中營養級魚類呈現顯著的負相關關系,與高、低營養級魚類呈正相關關系。這表明,SST異常增暖抑制了中營養級魚類的生長,但促進了高、低營養級魚類的生長,兩種效應相互抵消,導致SST與MTL無顯著的相關關系。由圖5(d)可以看出,雖然南海漁區7—12月海溫增暖也比較明顯,基本上在每10年0. 06 oC~0. 2 oC范圍內波動,秋季最強,但對南海漁區MTL 上升趨勢沒有貢獻。
與MTL不同,SST對渤黃海和東海漁區的FIB指數沒有顯著影響,但對南海漁區的影響較為顯著,主要是秋季SST與FIB指數呈現顯著的正相關關系(圖7)。因此,南海漁區FIB指數的增加與秋季海溫增暖有關,即南海秋季海溫增暖對于局地漁業生態系統持續健康有一定的貢獻。
4 結論與建議
4. 1 主要結論
本研究聚焦中國近海漁業資源結構與質量變化,以漁獲物平均營養級和漁業均衡指數為指標,較為系統地研究了人為捕撈和局地海溫變化對渤黃海、東海及南海漁業資源結構與質量的影響,得出如下主要結論。
(1)渤黃海和東海漁區,分別以低營養級和高營養級魚類占比最高,MTL呈下降趨勢;南海漁區以中營養級魚類占比最高,MTL無明顯變化趨勢。渤黃海和東海漁區的漁獲量與MTL呈現顯著的負相關關系,這表明人為過度捕撈已影響到漁業資源結構與質量。在南海漁區,漁獲量與MTL之間并沒有顯著相關性,這表明人為捕撈活動未對南海漁區漁業資源的結構與質量產生顯著影響。
(2)FIB指數在渤黃海和東海漁區的總體趨勢均是先上升后下降,在南海則表現出微弱的上升趨勢。3個漁區的漁獲量與FIB指數均呈現顯著的正相關關系,這表明漁獲量的增長可以抵消MTL降低帶來的影響,進而導致FIB指數上升,這種上升有助于維持漁業生態系統結構與功能的健康狀態。
(3)在渤黃海和東海漁區,拖網漁船作業是導致MTL下降的一個重要原因,但在南海漁區,拖網漁船作業對MTL影響不大。就FIB指數而言,在渤黃海和東海漁區,拖網的漁獲量占比最大,均與FIB指數呈現顯著的正相關關系,但在南海漁區,兩者的相關關系卻不顯著,南海漁區主要是流刺網的漁獲量與FIB指數呈現顯著的正相關關系,這表明3個漁區主要作業類型的漁獲量抵消了MTL下降的負面效應,使得漁業生態系統結構與功能得以維持。
(4)在渤黃海漁區,MTL 的下降趨勢是由于該區域5—6月份海溫的快速增暖,這種現象對高營養級魚類的生長產生了抑制作用。在東海漁區,秋季海溫增暖對MTL下降趨勢有一定的貢獻度。但在南海漁區,MTL與SST變化沒有呈現顯著的相關關系。渤黃海和東海漁區的FIB指數受SST變化影響較小,但南海漁區FIB指數的增加與秋季海溫增暖有關。
4. 2 建議
本研究著重分析了人為捕撈和氣候變化對中國近海漁業資源結構與質量的影響,因此,各漁區在開展漁業活動時要綜合考量這兩個因素。①對于渤黃海漁區來說,建議合理控制捕撈量,根據海洋漁業資源最大產量的可持續性,確定適宜的捕撈漁船數量和功率,繼續落實漁船“雙控”政策;加強對拖網等非選擇性漁具的管理和控制,積極發展流刺網、釣具等作業方式,加大政策扶持,促進拖改流工作取得良好成效。②對于東海漁區來說,要嚴格控制近海捕撈強度,壓縮機動漁船,特別是小功率漁船的數量,采取合理的價格收購居民漁船并給予一定補償,使部分專業漁民從海洋捕撈中退出;同時減少非選擇性漁具(如拖網)的使用,優化漁具的選擇,實施負責任的捕撈方式。③基于渤黃海和東海漁區海溫增暖對MTL下降的顯著作用,應采用遙感技術、GIS技術等,加強對海溫的動態監測和評估,增進觀測資料的共享;同時通過馴化手段提高一些經濟魚類的耐高溫能力,以更好地適應外部環境的變化。④對于南海漁區來說,雖然漁區內整體高營養級漁獲物的漁獲量并沒有出現下降現象,但是北部灣等近海大陸架內的漁獲物由于受到過度捕撈的影響,已經出現了壽命長、高營養級的魚類數量不斷下降,而壽命短、低營養級的魚類數量上升的現象,因此,在南海漁區,可適當拓展捕撈作業區,發展遠洋捕撈,調整完善外海漁業結構,減輕近海捕撈壓力。
本研究使用漁業統計數據對中國近海漁業資源的結構與質量進行探究,但由于漁區數據有限,故采用沿海不同省份的漁業統計數據表征各漁區數據,這可能會使分析結果的精確度降低。同時,技術進步、政策法規等因素也會對中國近海漁業資源的結構與質量產生影響,在以后的研究中需要綜合多種因素進行評價。
參考文獻
[1] 陳新軍. 海洋漁業資源可持續利用評價[D]. 南京:南京農業大
學,2001.
[2] 魯泉,蘇雪,方舟,等. 基于漁獲量平均營養級的東印度洋漁業
資源可持續利用評價[J]. 海洋學報,2021,43(8):118-127.
[3] 孫吉亭. 中國海洋漁業可持續發展研究[D]. 青島:中國海洋大
學,2003.
[4] 朱玉貴,趙麗麗,劉燕飛. 海洋漁業資源可持續利用研究[J]. 中
國人口·資源與環境,2009,19(2):166-169.
[5] 李紅艷,王穎,劉天紅,等. 基于生態足跡的我國海洋水產資源
可持續利用研究[J]. 漁業研究,2021,43(5):509-516.
[6] 劉瑞玉. 關于我國海洋生物資源的可持續利用[J]. 科技導報,
2004,22(11):28-31.
[7] 盧秀容. 中國海洋漁業資源可持續利用和有效管理研究[D]. 武
漢:華中農業大學,2005.
[8] 蔡榕碩,劉克修,譚紅建. 氣候變化對中國海洋和海岸帶的影
響、風險與適應對策[J]. 中國人口·資源與環境,2020,30(9):
1-8.
[9] 徐連章. 新制度經濟學視角下的我國海洋漁業資源可持續利用
研究[D]. 青島:中國海洋大學,2010.
[10] 孫康,周曉靜,蘇子曉,等. 中國海洋漁業資源可持續利用的動
態評價與空間分異[J]. 地理科學,2016,36(8):1172-1179.
[11] WU L X,CAI W J,ZHANG L P,et al. Enhanced warming over the
global subtropical western boundary currents[J]. Nature climate
change,2012,2(3):161-166.
[12] 吳佳穎,薛瑩,劉笑笑,等. 黃、渤海漁業生物平均營養級的長
期變動[J]. 中國海洋大學學報(自然科學版),2017,47(11):
53-60.
[13] 李繼龍,曹坤,丁放,等. 基于漁獲物統計的中國近海魚類營養
級結構變換及其與捕撈作業的關系[J]. 中國水產科學,2017,
24(1):109-119.
[14] 劉子飛. 我國近海捕撈漁業管理政策困境、邏輯與取向[J]. 生
態經濟,2018,34(11):47-53.
[15] 劉子飛. 中國近海漁場荒漠化:評價、原因與治理[J]. 農業經
濟問題,2019,40(6):105-116.
[16] 閆文彥,蔣楊徽,蔡立煌,等. 舟山市不同作業類型漁船捕撈產
量的灰色分析[J]. 浙江海洋大學學報(自然科學版),2018,37
(5):468-474.
[17] 宋倫,王年斌,董婧,等. 捕撈對長山群島海域漁業生態系統的
影響[J]. 生態學雜志,2010,29(8):1578-1584.
[18] LIANG C,PAULY D. Fisheries impacts on China's coastal ecosystems:
unmasking a pervasive‘ fishing down effect[J]. PLoS one,
2017,12(3):e0173296.
[19] LINDEMAN R L. The trophic?dynamic aspect of ecology[J]. Bulletin
of mathematical biology,1991,53(1):167-191.
[20] PAULY D, CHRISTENSEN V, DALSGAARD J, et al. Fishing
down marine food webs[J]. Science,1998,279(5352):860-863.
[21] PAULY D, CHRISTENSEN V, WALTERS C. Ecopath, ecosim,
and ecospace as tools for evaluating ecosystem impact of fisheries
[J]. ICES journal of marine science, 2000, 57(3): 697-706.
[22] DUNLOP E S, GOTO D, JACKSON D A. Fishing down then up
the food web of an invaded Lake[J]. Proceedings of the National
Academy of Sciences of the United States of America, 2019, 116
(40): 19995-20001.
[23] 張忠,楊文波,陳新軍,等. 基于漁獲量平均營養級的東南大西
洋漁業資源狀況分析[J]. 海洋漁業,2015,37(3):197-207.
[24] 丁琪,陳新軍,李綱,等. 基于漁獲統計的西北太平洋漁業資源
可持續利用評價[J]. 資源科學,2013,35(10):2032-2040.
[25] MILESSI A C, ARANCIBIA H. The mean trophic level of Uruguayan
landings during the period 1990-2001[J]. Fisheries research,
2005, 74(1-3): 223-231.
[26] 張瑛,李大海,耿濤. 氣候變化背景下我國深藍漁業的發展戰
略研究[J]. 山東大學學報(哲學社會科學版),2018(6):
121-129.
[27] ASSIS J, TYBERGHEIN L, BOSCH S, et al. Bio?ORACLE
v2. 0: extending marine data layers for bioclimatic modelling[J].
Global ecology and biogeography, 2018, 27(3): 277-284.
[28] 許友卿,曹占旺,丁兆坤,等. 高溫對魚類的影響及其預防研究
[J]. 水產科學,2010,29(4):235-242.
[29] 黃佳興,龔玉艷,徐姍楠,等. 南海中西部漁場主要漁業生物碳
氮穩定同位素特征[J]. 熱帶海洋學報,2019,38(1):76-84.
[30] 陳爽,陳新軍. 氣候變化對東北大西洋漁獲物組成、多樣性和
營養級的影響[J]. 海洋學報,2020,42(10):100-109.
[31] 劉尊雷,陳誠,袁興偉,等. 基于調查數據的東海小黃魚資源變
化模式及評價[J]. 中國水產科學,2018,25(3):632-641.
[32] 吳紹洪,趙東升. 中國氣候變化影響、風險與適應研究新進展
[J]. 中國人口·資源與環境,2020,30(6):1-9.
[33] 肖啟華,黃碩琳. 氣候變化對東南太平洋智利竹筴魚漁獲量的
影響[J]. 中國水產科學,2021,28(8):1020-1029.
[34] 孫明,劉修澤,李軼平,等. 應用氮穩定同位素技術研究遼東灣
海域主要漁業生物的營養級[J]. 中國水產科學,2013,20(1):
190-198.
[35] 白懷宇,王玉堃,張廷廷,等. 基于碳、氮穩定同位素研究黃海
及東海北部主要魚類的營養級和食性[J]. 漁業科學進展,
2021,42(2):10-17.
[36] 高春霞,戴小杰,田思泉,等. 基于穩定同位素技術的浙江南部
近海主要漁業生物營養級[J]. 中國水產科學,2020,27(4):
438-453.
[37] 鄧景耀,姜衛民,楊紀明,等. 渤海主要生物種間關系及食物網
的研究[J]. 中國水產科學,1997,4(4):1-7.
[38] PAULY D,WATSON R. Background and interpretation of the
‘Marine Trophic Index as a measure of biodiversity[J]. Philosophical
transactions of the Royal Society B:biological sciences,
2005,360(1454):415-423.
[39] 鮑穎,蘭健,王毅. 南海在1997/1998年El Ni?o事件后的異常變
化[J]. 地球科學進展,2008,23(10):1027-1036.
[40] 陳明誠,黃玉蓉. 典型拉尼娜事件與1988—1989年拉尼娜事件
生命史演變差異性成因分析[J]. 大氣科學學報,2021,44(3):
428-440.
[41]WANG G H, XIE S P, QU T D, et al. Deep South China Sea circulation
[J]. Geophysical research letters, 2011, 38(5): L05601.
[42]WANG B, HUANG F, WU Z W, et al. Multi?scale climate variability
of the South China Sea monsoon: a review[J]. Dynamics of
atmospheres and oceans, 2009, 47(1-3): 15-37.
[43]ZHAO Y H, LIANG X S, YANG Y. The Kuroshio intrusion into
the South China Sea at Luzon Strait can be remotely influenced by
the downstream intrusion into the East China Sea[J]. Journal of
geophysical research?oceans, 2023, 128(8): e2023JC019868.
[44]ZHANG L, REN X Y, CAI W J, et al. Weakened western Indian
Ocean dominance on Antarctic sea ice variability in a changing climate
[J]. Nature communications, 2024, 15(1): 3261.
[45] 徐策,張力,余靜,等. 氣候變化對中國近海捕撈漁業的影響:
以太平洋年代際濤動為例[J]. 資源科學,2022,44(2):
386-400.
[46]MANTUA N J, HARE S R, ZHANG Y, et al. A Pacific interdecadal
climate oscillation with impacts on salmon production[J].
Bulletin of the American Meteorological Society, 1997, 78(6):
1069-1079.
[47]王萬良,張忭忭,周建設,等. 不同水溫對亞東鮭幼魚生長及存
活的影響[J]. 水產科技情報,2019,46(1):24-27.
[48]POLOVINA J J, MITCHUM G T, GRAHAM N E, et al. Physical
and biological consequences of a climate event in the Central North
Pacific[J]. Fisheries oceanography, 2010, 3(1): 15-21.
(責任編輯:田紅)
