4種夏櫟對低溫脅迫的生理響應及抗寒性評價
2024-07-18 00:00:00姜恒張志剛楊建軍李斌王興勝張曉紅
河北大學學報(自然科學版) 2024年4期


DOI:10.3969/j.issn.10001565.2024.04.009
摘要:為探究低溫脅迫下夏櫟(Quercus robur)生理指標的響應及抗寒性,以普通夏櫟、速生夏櫟、裂葉夏櫟和垂枝夏櫟為實驗材料,測定在不同溫度下細胞膜通透性、超氧化物歧化酶(SOD)、過氧化物酶(POD)、可溶性蛋白等指標,并利用擬合Logistic方程計算半致死溫度,結合隸屬函數法對4種夏櫟綜合評價.結果表明:4種夏櫟細胞膜通透性均隨著溫度降低而增加,SOD和POD活性以及可溶性蛋白、脯氨酸和丙二醛(MDA)含量均逐漸下降;4種夏櫟半致死溫度為-41.9~-38.2 ℃,其中普通夏櫟半致死溫度最低,速生夏櫟半致死溫度最高;4種夏櫟抗寒性強弱為普通夏櫟gt;垂枝夏櫟gt;裂葉夏櫟gt;速生夏櫟.綜上可知,普通夏櫟抗寒性最強,在不同地區推廣和繁育具有顯著優勢.
關鍵詞:夏櫟;抗寒性;隸屬函數法;生理指標;半致死溫度
中圖分類號:Q945.79文獻標志碼:A文章編號:10001565(2024)04040608
Physiological response and cold resistance evaluation of four
species of Quercus robur to low temperature stress
JIANG Heng1,2, ZHANG Zhigang3 , YANG Jianjun1,2, LI Bin4, WANG Xingsheng4, ZHANG Xiaohong4
(1. Key Laboratory of Oasis Ecology of Education Ministry, College of Ecology and Environment,
Xinjiang University,Urumqi" 830017,China; 2.Technology Innovation Center for Ecological
Monitoring and Restoration of Desert-Oasis," Ministry of Natural Resources (MNR), Urumqi "830002,China;
3. Key Laboratory of Forest Resources and Utilization in Xinjiang of National Forestry and Grassland
Administration, Institute of Afforestation and Desertification Control, Xinjiang Academy of Forestry,
Urumqi" 830000,China; 4. Yili Prefecture Forest Tree Breeding Experiment Center, Chabchal" 835311, China)
Abstract: In order to explore the response of Quercus robur" physiological indicators and the strength of cold resistance under low temperature stress, the common Q. robur, fast-growing Q. robur, split-leaf Q. robur and weeping Q. robur were used as experimental materials. The cell membrane permeability, SOD activity, POD activity and soluble protein at different temperatures were measured, and the fitted Logistic
收稿日期:20240109;修回日期:20240508
基金項目:新疆三農骨干人才培養項目(2022SNGGNT082);新疆維吾爾自治區2023年度重大科技專項(2023A02008)
第一作者:姜恒(1998—),男,新疆大學在讀碩士研究生,主要從事生態修復研究.E-mail:670551763@qq.com
通信作者:楊建軍(1978—),男,新疆大學教授,博士生導師,主要從事干旱區水土保持與荒漠化防治研究.
E-mail:yjj@xiu.edu.cn
張曉紅(1972—),女,伊犁州林木良種繁育試驗中心高級工程師,主要從事林木良種選育與推廣研究.
E-mail:1436787731@qq.com
equation was used to calculate the half-lethal temperature. And the function method was used for comprehensive evaluation of the four Q. robur species. The results showed that the cell membrane permeability of the four species of Q. robur showed an increasing trend as the temperature decreased, and the SOD activity, POD activity, soluble protein content, proline content and MDA content showed a decreasing trend as a whole, and the cell membrane permeability was calculated by combining the Logistic equation. The semi-lethal temperature of four species of Q. robur ranged from -41.9 ℃ to -38.2 ℃. The common Q. robur had the highest semi-lethal temperature and the fast-growing Q. robur had the lowest semi-lethal temperature. This is consistent with the evaluation by the membership function method. The four species of Q. robur had strong cold resistance. The cold resistance ranking of the four Q. robur species was common Q. roburgt; weeping Q. roburgt;split-leaf Q. robur gt; fast growing Q. robur. The study show that common Q. robur is the most resistant to cold and has significant advantages for promotion and breeding in different regions.
Key words: Quercus robur; cold resistance; membership function method; physiological index; half-lethal temperature
夏櫟(Quercus robur)隸屬殼斗科(Fagaceae)櫟屬(Quercus),落葉喬木,也稱夏橡、英國櫟,其樹體高大通直,成年后樹高可達40 m,在中國的新疆、北京和山東均有栽培,具有材質好、壽命長、樹冠開闊和樹形優美等優點,對生長環境要求較低,生長范圍十分廣泛,是十分理想的造林樹種,在干旱和半干旱區林業造林中具有良好的推廣前景[1-2].冬季低溫是制約植物能否安全越冬的重要因素[3],受全球氣候變化影響,春季氣候多變,寒潮、冰雹、低溫霜凍等災害性天氣頻發,嚴重阻礙了植物的生長發育[4],因此,研究夏櫟在低溫脅迫下的生理響應及抗寒性對夏櫟推廣栽培具有重要意義.
低溫脅迫下,植物體內會發生一系列生理生化反應來適應外界低溫,單一的指標無法對植物的抗寒性進行評價.李呈呈等[5]對6個彩葉桂品種的研究發現,細胞膜通透性、超氧化物歧化酶(SOD)、過氧化物酶(POD)和可溶性蛋白等指標對植物抵御低溫脅迫有重要作用,可作為植物抗寒性評價指標;牟開萍等[6]以20個月季品種為研究對象,測定了相對電導率(REC)、丙二醛(MDA)和可溶性蛋白等,并結合主成分分析、聚類分析和隸屬函數法對其進行了綜合評價;賈金輝等[7]以8種釀酒葡萄為研究對象,測定了低溫脅迫下枝條萌芽率和相關生理指標,發現隸屬函數法和半致死溫度評價結果的一致性;楊振亞等[8]研究了娜塔櫟、沼生櫟、舒瑪櫟、雙色櫟在低溫脅迫下的響應,并通過半致死溫度確定了其抗寒性.因此,選擇合適的指標和評價方法尤為重要.目前關于夏櫟的研究多集中在育苗移栽和白粉病等方面[9-11],而對夏櫟抗寒性的研究較少.
本研究選取垂枝夏櫟、速生夏櫟、裂葉夏櫟和普通夏櫟1年生枝條為實驗材料,在人工模擬低溫的條件下,通過測定細胞膜通透性、可溶性蛋白和MDA含量及SOD和POD活性,并擬合Logistic方程計算半致死溫度,采用隸屬函數法對4種夏櫟進行綜合評價,篩選出抗寒性強的夏櫟品系,為夏櫟在不同地區推廣和繁育提供理論依據.
1材料與方法
1.1實驗材料
實驗材料來自伊犁州林木良種繁育試驗中心,品系為垂枝夏櫟、速生夏櫟、裂葉夏櫟和普通夏櫟.先使用自來水清洗枝條表面灰塵,再用去離子水沖洗3次,用紗布吸干表面水分.枝條清洗完畢后,對其進行剪切(避開芽孢,每段至少留存2個芽孢),每段長10 cm,將剪切好的枝條裝入自封袋,并貼好標簽,標記好品系名稱.
1.2實驗方法
實驗開始后將全部枝條同時放入低溫冰柜內.實驗處理設置為6個溫度梯度,分別為5、-20、-25、-30、-35、-40 ℃,以5 ℃為對照,溫度調整為每8 h下降0.6 ℃,達到對應溫度后保持24 h,隨后取出樣條測定抗寒生理指標并進行水培實驗.
將低溫脅迫的枝條用去離子水沖洗3次,將枝條下端1 cm剪除,置于三角瓶中水培,每個處理10段枝條,將三角瓶放置于組培架上,陽光培養12 h,每隔2 d換水、觀測枝條復蘇情況并統計枝條成活率(芽鱗開裂、膨大為成活).根據枝條恢復后存活枝的成活率評價各品系的抗寒性.
枝條成活率=成活枝數/水插枝總數×100%.
1.3指標測定及方法
生理指標測定:SOD活性采用氮藍四唑還原法測定[12];POD活性采用愈創木酚法測定[12];可溶性蛋白含量采用考馬斯亮藍法測定[12];MDA采用硫代巴比妥酸法測定[12];脯氨酸采用茚三酮顯色法測定[13].
細胞膜通透性測定:細胞膜通透性采用電導率法測定[12].對不同溫度下各品系夏櫟細胞膜通透性進行Logistic擬合,計算出各品系的半致死溫度(LT50)[14].
擬合方程為Y=K/(1+ae-bx),(1)
LT50=-(ln a)/b,(2)式中:Y為相對電導率;K為常數,代表電解質滲出的最大量;e為自然底數;a、b均為方程參數;x為處理溫度;LT50為曲線的拐點溫度(半致死溫度).
1.4數據處理及分析
采用隸屬函數法對不同品系夏櫟進行評價,與抗寒性呈正相關指標(SOD、POD、可溶性蛋白、脯氨酸),計算公式為U(Xij)=(Xij-Xjmin)/(Xjmax-Xjmin),(3)與抗寒性呈負相關指標(相對電導率和丙二醛含量),計算公式為U(Xij)=1-(Xij-Xjmin)/(Xjmax-Xjmin),(4)式中:U(Xij)為各指標的平均值;Xij為第i個品系下第j個指標的隸屬度值;Xjmin和Xjmax分別為第j個指標不同溫度下最小值和最大值.
對各品系在不同溫度下的隸屬度值求均值,按照隸屬度值排序即為不同品系夏櫟的抗寒能力.使用Microsoft Office和Origin 2022進行數據整理,SPSS 27.0進行單因素方差分析和顯著性分析(Plt;0.05).
2結果與分析
2.1低溫脅迫對夏櫟細胞膜通透性的影響
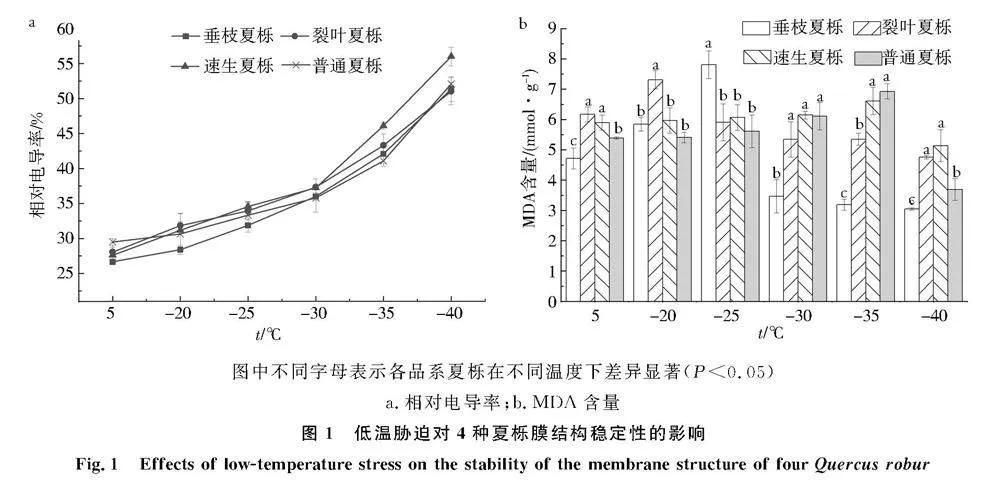
低溫脅迫下夏櫟各品系細胞膜通透性變化如圖1a所示,隨著溫度的降低,各品系細胞膜通透性均呈上升趨勢,其中垂枝夏櫟和普通夏櫟的變化趨勢較為一致.對照組4種夏櫟枝條相對電導率均保持在28%~33%,并在-40 ℃時均達到最大值.對照組垂枝夏櫟細胞膜通透性最小,為28.39%,其次是裂葉夏櫟,為30.65%,當溫度為-30~-25 ℃時,速生夏櫟的細胞膜通透性變化最大,而普通夏櫟的變化最小,其余2種均在二者之間,4種夏櫟葉片細胞膜通透性達到最大值時,相對電導率依次為速生夏櫟gt;裂葉夏櫟gt;普通夏櫟gt;垂枝夏櫟.
2.2低溫脅迫對夏櫟MDA的影響
低溫脅迫下夏櫟各品系MDA含量變化如圖1b所示,隨著溫度的降低,各品系的MDA含量均呈現先上升后下降的趨勢,其中速生夏櫟和普通夏櫟MDA含量的變化趨勢較為一致.經過-35 ℃處理后,速生夏櫟和普通夏櫟枝條的MDA含量均達到最大值,分別為6.6 mmol/g和6.9 mmol/g,而裂葉夏櫟和垂枝夏櫟分別在-20 ℃和-25 ℃處理下達到最大值,分別為7.3 mmol/g和7.8 mmol/g.垂枝夏櫟的MDA含量在低溫脅迫下均與其他品系存在顯著差異.與對照組相比,4種夏櫟枝條受到低溫脅迫后MDA含量整體表現為速生夏櫟gt;裂葉夏櫟gt;垂枝夏櫟gt;普通夏櫟.
圖中不同字母表示各品系夏櫟在不同溫度下差異顯著(Plt;0.05)
a.相對電導率;b.MDA含量
2.3Lotistic方程的建立及LT50的確定
低溫脅迫下對4種夏櫟相對電導率進行方程擬合,結果如表1所示,低溫脅迫處理下各品系夏櫟的相對電導率和溫度之間的關系可以用公式(1)和公式(2)擬合,R2值為0.891~0.979,表明溫度和相對電導率有很強的線性關系,擬合結果具有可靠性.4種夏櫟的LT50為-41.9~-38.2 ℃,根據LT50大小可得夏櫟抗寒性為普通夏櫟gt;裂葉夏櫟gt;垂枝夏櫟gt;速生夏櫟.表1低溫脅迫下4種夏櫟的LT50及擬合方程
Tab.1LT50 and fitting equations of four Quercus robur under low-temperature stress品系擬合方程R2t/℃垂枝夏櫟Y=100/(1+6.997e-0.048)0.915-40.5速生夏櫟Y=100/(1+5.957e-0.047)0.891-38.2裂葉夏櫟Y=100/(1+5.781e-0.043)0.908-41.1普通夏櫟Y=100/(1+5.269e-0.04)0.979-41.9
2.4低溫脅迫對夏櫟SOD的影響
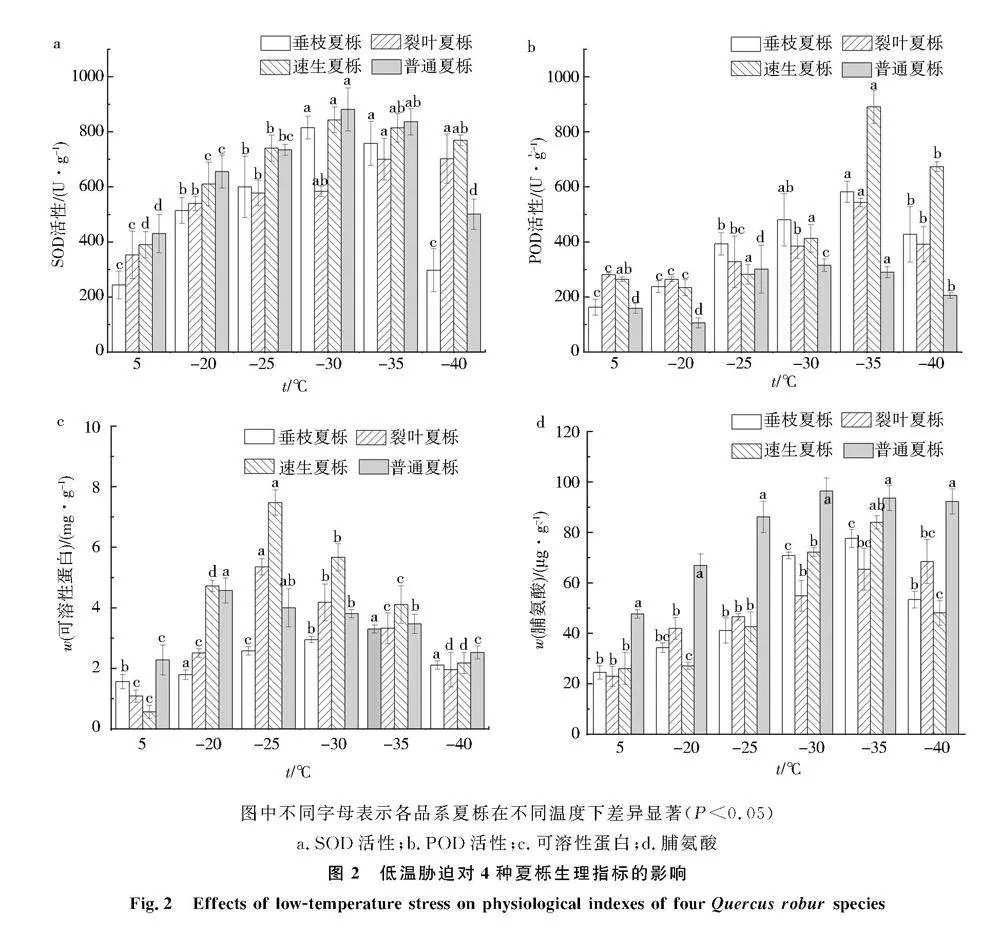
低溫脅迫下夏櫟SOD酶活性變化如圖2a所示,隨著溫度的降低,4種夏櫟1年生枝條SOD酶活性呈現先升高后降低的趨勢.4種夏櫟SOD酶活性最大值均不相同,其中垂枝夏櫟、速生夏櫟和普通夏櫟均在-30 ℃時枝條的SOD活性達到最大值,分別為815.01、843.28、881.19 U/g,而裂葉夏櫟的SOD活性在-35 ℃達到最大值,為702.15 U/g.與對照組相比,垂枝夏櫟的SOD含量增加最多,為235%,裂葉夏櫟的SOD含量增加最少,為98.55%.4種夏櫟枝條受到低溫脅迫后SOD活性增加幅度為垂枝夏櫟gt;速生夏櫟gt;普通夏櫟gt;裂葉夏櫟.
2.5低溫脅迫對夏櫟POD的影響
低溫脅迫后4種夏櫟的POD活性如圖2b所示,其中,普通夏櫟對照組的POD活性為159 U/g,當經過-20 ℃處理后,POD活性有所下降,但與對照組差異不顯著,隨著溫度的降低,POD活性逐漸增加,到-30 ℃時達到最大值,為315.33 U/g,比對照組增加了98.3%,之后隨著溫度的降低,POD活性隨之降低,但均顯著高于對照組.垂枝夏櫟、裂葉夏櫟和速生夏櫟的POD活性整體呈現出先增加后下降的趨勢,變化較為相似,在-35 ℃低溫脅迫下,垂枝夏櫟、裂葉夏櫟和速生夏櫟的POD活性均達到最大值,分別為582、543、890.67 U/g,且速生夏櫟POD活性顯著高于其他3種品系,比對照組分別增加了257.1%、93.2%和236.95%,之后隨著溫度的增加POD活性隨之下降.4種夏櫟POD活性低溫脅迫后整體上升幅度為速生夏櫟gt;垂枝夏櫟gt;裂葉夏櫟gt;普通夏櫟.
2.6低溫脅迫對夏櫟可溶性蛋白含量的影響
低溫脅迫下4種夏櫟的可溶性蛋白含量(質量分數)如圖2c所示,對照組4種夏櫟可溶性蛋白含量均處于最小值,其中速生夏櫟含量最低,其次為普通夏櫟,而裂葉夏櫟的可溶性蛋白含量最高,且顯著高于其他3種品系.隨著溫度的降低,各品系夏櫟呈現出先增加后下降的趨勢,4種夏櫟達到最大值的可溶性蛋白含量各不相同,-35 ℃處理后除速生夏櫟的可溶性蛋白含量與對照組無顯著差異外,其他溫度處理下均存在顯著差異.普通夏櫟的可溶性蛋白含量在-20 ℃處理下達到最大值,相較于對照組增加了321.3%,裂葉夏櫟在-25 ℃處理下達到最大值,增加了135.2%,4種夏櫟受到低溫脅迫后可溶性蛋白含量增加幅度為速生夏櫟gt;裂葉夏櫟gt;普通夏櫟gt;垂枝夏櫟.
2.7低溫脅迫對夏櫟脯氨酸含量的影響
低溫脅迫下4種夏櫟的脯氨酸含量(質量分數)如圖2d所示,隨著溫度的降低,速生夏櫟、垂枝夏櫟和普通夏櫟的脯氨酸含量呈現先增加后下降的趨勢,而裂葉夏櫟的脯氨酸含量呈現出持續增大的趨勢.在-35 ℃處理下,垂枝夏櫟和速生夏櫟的脯氨酸含量達到最大值,普通夏櫟的脯氨酸含量在-30 ℃處理下達到最大值,相較于對照組增加了102.3%.與對照組相比,4種夏櫟受到低溫脅迫后脯氨酸含量達到最大值時增加幅度整體為垂枝夏櫟gt;速生夏櫟gt;裂葉夏櫟gt;普通夏櫟.圖中不同字母表示各品系夏櫟在不同溫度下差異顯著(Plt;0.05)
a.SOD活性;b.POD活性;c.可溶性蛋白;d.脯氨酸
2.8低溫脅迫下不同品系夏櫟枝條水培萌發情況
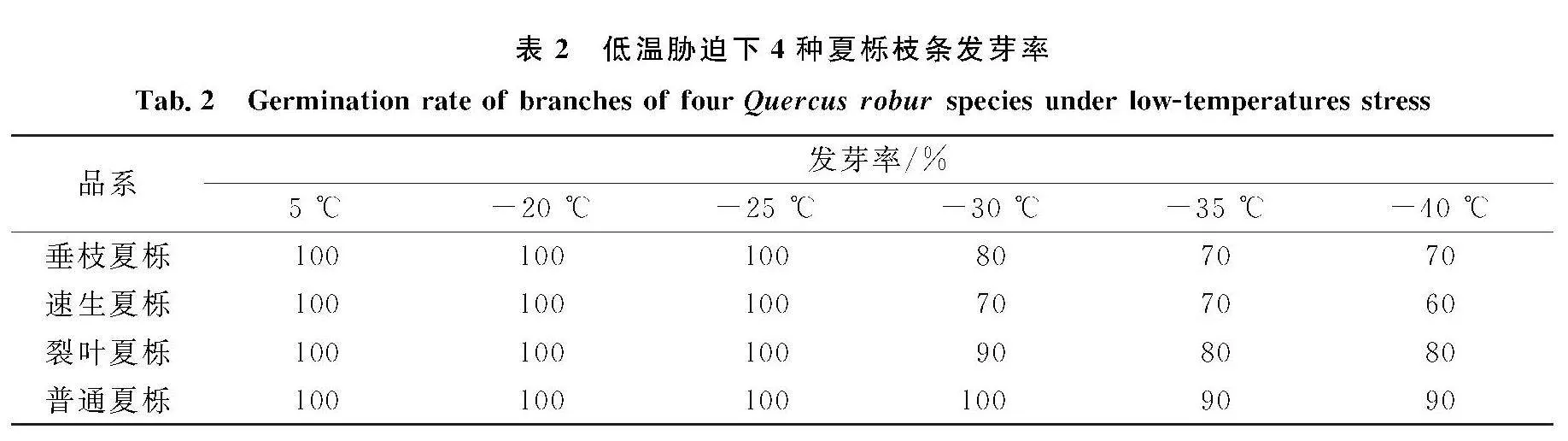
水培恢復法是驗證植物在抵御低溫脅迫后是否遭受凍害最簡單有效的方法.低溫脅迫后不同品系水培萌芽情況如表2所示,隨著溫度的降低,不同夏櫟發芽率也有所不同,在-25~-20 ℃處理后,4種品系夏櫟水培發芽率均達到100%;-30 ℃處理后,垂枝夏櫟、裂葉夏櫟和速生夏櫟的發芽率均有所降低,分別為80%、70%和90%.隨著溫度的下降,普通夏櫟、裂葉夏櫟和垂枝夏櫟的發芽率較為穩定,經過-40 ℃處理后,速生夏櫟枝條的發芽率最低,為60%;普通夏櫟的發芽率最高,為90%,表明4種品系夏櫟均能在-40 ℃低溫下存活,并且普通夏櫟抗寒性強于垂枝夏櫟、裂葉夏櫟和速生夏櫟.
2.9不同品系夏櫟的抗寒性分析
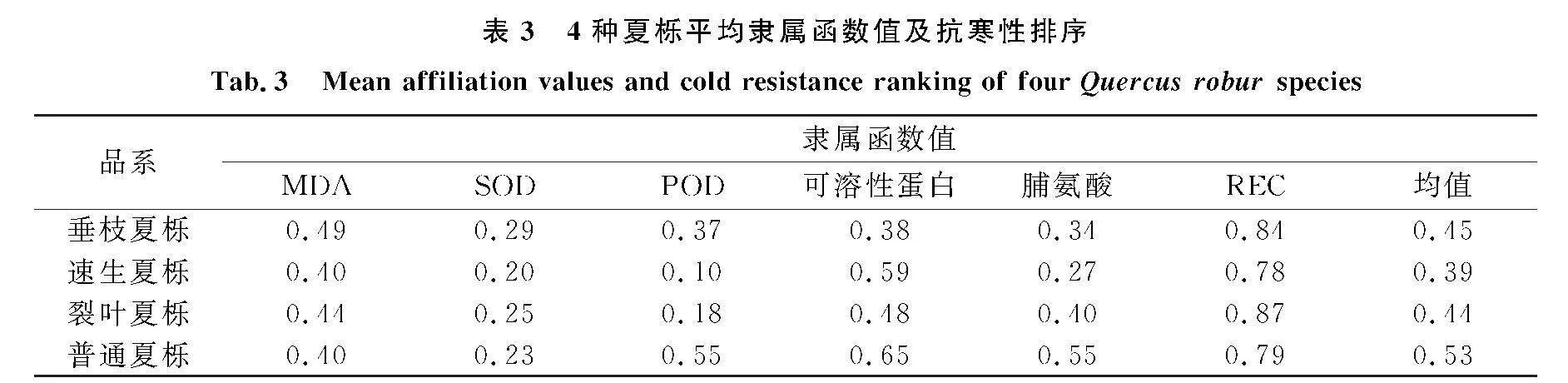
對低溫脅迫下4種品系夏櫟的細胞膜通透性等生理指標采用隸屬函數進行綜合分析及評價.隸屬函數值與抗寒性存在正相關關系,隸屬函數值越大,其抗寒性越強,反之抗寒性越弱.
由表3可知,4種夏櫟中普通夏櫟抗寒性最強,速生夏櫟的抗寒性最差,4種夏櫟抗寒性由大到小依次為普通夏櫟gt;垂枝夏櫟gt;裂葉夏櫟gt;速生夏櫟,這一結果與水培復蘇實驗的結果基本一致.
3討論與結論
細胞膜是植物受到低溫脅迫后細胞反應最直接的部位,也是容易受到低溫脅迫傷害的部位[15-17].低溫脅迫下植物細胞膜結構和功能發生改變,細胞內離子和糖類物質滲出,通透性增大,從而導致胞外電導率增大.因此,電導率值可用來表示植物在低溫脅迫下受影響的程度,且相對電導率越大,表明植物越容易受到低溫脅迫的影響,抗寒性越弱[18].本研究中,4種夏櫟相對電導率隨著溫度的降低呈增加趨勢,這與王金鑫等[19]在研究低溫脅迫對不同果型翅果油樹細胞膜通透性變化表現一致.溫度高于-35 ℃時,4種夏櫟細胞膜通透性表現相當;溫度低于-35 ℃時,速生夏櫟的細胞膜通透性迅速增大,表明速生夏櫟的抗寒性低于其他3種夏櫟.LT50表示植物抗寒能力和耐低溫的極限,LT50越低,表明植物的抗寒性越強[20].本研究中,4種夏櫟抗寒性從小到大排序為速生夏櫟<垂枝夏櫟<裂葉夏櫟<普通夏櫟,LT50依次為-38.2、-40.5、-41.1、-41.9 ℃.
MDA是細胞膜過氧化的最終產物,已經成為植物抗寒評價的重要指標之一.MDA在植物體內的積累量與植物的抗寒性有較強的相關性,MDA含量越高,說明細胞膜過氧化越嚴重,即抗寒性越弱[21-22].4種夏櫟的MDA含量在低溫脅迫初期隨著溫度的降低呈上升趨勢,隨著低溫脅迫的下降,4種夏櫟在低溫脅迫下MDA含量有所降低,當溫度低于-25 ℃時,垂枝夏櫟MDA含量顯著下降,并顯著低于其他品系,表明垂枝夏櫟在低溫初期抗寒性要弱于其他3種夏櫟,這與溫日宇等[23]在研究不同低溫脅迫下不同藜麥幼苗的MDA含量變化與抗寒性評價結果相一致.隨著低溫脅迫的加劇,普通夏櫟和垂枝夏櫟的MDA含量顯著低于速生夏櫟和裂葉夏櫟,即普通夏櫟和垂枝夏櫟的抗寒性高于裂葉夏櫟和速生夏櫟,說明低溫在脅迫初期導致4種夏櫟細胞膜損害加重,但隨著低溫的加劇細胞膜起到了一定的保護作用,這與LT50計算出的4種夏櫟的抗寒性結果相一致.本實驗中,4種夏櫟在低溫脅迫后枝條復蘇率與LT50結果基本一致.
低溫脅迫下細胞膜脂氧化加重,體內超氧自由基陰離子、過氧化氫、羥基自由基等活性氧含量增加,降低細胞膜的功能,嚴重時導致植物細胞功能紊亂[24-25].SOD和POD是植物體抗氧化系統中重要的酶,能夠清除體內產生過量的活性氧,對維持植物正常的生理功能和抵御低溫脅迫具有重要意義,因此在低溫脅迫下,SOD和POD活性含量與植物的抗寒性有較強的關系,這與李桂榮等[26]在研究6種山葡萄在低溫脅迫下抗氧化酶的結果一致.本研究中4種夏櫟的SOD和POD含量在低溫脅迫初期有所下降,之后呈上升趨勢,這可能是由于低溫脅迫初期植物體內保護酶活性降低或分解有關,再次升高可能是由于植物已經適應了低溫脅迫從而導致抗氧化酶升高.在溫度脅迫下,速生夏櫟POD含量在最大值時增加顯著高于其他夏櫟,相反普通夏櫟的變化相對較小,這說明速生夏櫟對低溫脅迫的變化較為敏感,而普通夏櫟對低溫脅迫有較強的適應能力.隨著溫度降低,抗氧化酶活性降低,推測是因為植物細胞膜氧化加劇,活性氧產生超過保護酶的清除能力.
植物細胞滲透調節物質在應對環境脅迫和維持細胞內外水分平衡方面發揮著重要作用[27],低溫脅迫條件下,植物細胞會積累可溶性蛋白和脯氨酸,以增加細胞內的滲透濃度,從而緩解低溫脅迫對植物造成的損害[28-29].本研究發現抗寒性強的普通夏櫟脯氨酸含量高于其他夏櫟,即脯氨酸含量與夏櫟抗寒性呈正相關,雷夢瑤等[30]對石榴枝條對低溫脅迫的響應的結果顯示,抗寒性強的千層花和紅雙喜脯氨酸含量高于另外2種夏櫟,與本實驗的研究結果一致.從可溶性蛋白含量變化發現,抗寒性差的速生夏櫟可溶性蛋白在低溫脅迫初期增加最多并顯著高于其他3種夏櫟,可能是因為速生夏櫟對低溫變化較為敏感,細胞持水能力下降,需要增加大量的可溶性蛋白來降低細胞冰點,保護植物免受低溫損傷.反而抗寒性強的普通夏櫟可溶性蛋白含量相較于抗寒性弱的速生夏櫟更加穩定,這與李桂榮等[31]的研究結果一致.推測的原因可能是因為普通夏櫟體內本身含有較高的可溶性蛋白且本身耐寒性較好,受到低溫脅迫后體內可溶性蛋白或僅需小幅度提高即可應對外界脅迫帶來的損傷.
評價植物抗寒性的指標有很多,在低溫脅迫過程中單一指標無法客觀判斷植物的抗寒能力[32-33],除了結合多指標對植物的抗寒性進行評價,還需要通過水培條件下不同夏櫟的萌芽率來確定植物的抗寒能力.本文采用隸屬函數法并結合水培條件下枝條萌芽率對4種夏櫟進行綜合評價,結果發現隸屬函數法與枝條萌芽率結果基本一致.普通夏櫟在低溫脅迫下表現最好,基本確定普通夏櫟可作為在不同地區推廣和繁育的品系,4種夏櫟抗寒性由強到弱依次為普通夏櫟gt;垂枝夏櫟gt;裂葉夏櫟gt;速生夏櫟.
參考文獻:
[1]白永強,李永華,朱強,等.夏橡高效育苗技術試驗[J].林業科技通訊, 2022(10): 70. DOI:10.13456/j.cnki.lykt.2022.01.21.0001.
[2]石海燕,張志剛,李宏,等.IAA對幼苗期夏橡生長及光合特性的影響[J].河北大學學報(自然科學版), 2022, 42(5): 519-529. DOI:10.3969/j.issn.1000-1565.2022.05.011.
[3]李文明,辛建攀,魏馳宇,等.植物抗寒性研究進展[J].江蘇農業科學, 2017, 45(12): 6-11. DOI:10.15889/j.issn.1002-1302.2017.12.002.
[4]HU L, XIAOFENG W, SHUJIANG C, et al. Dynamic analysis and evaluation of Xinjiang forest resources: based on RS and GIS[J]. Journal of Geographical Sciences, 2005, 15(3): 346-352. DOI:10.1007/BF02837522.
[5]李呈呈,吳其超,馬燕,等.6個彩葉桂品種對低溫脅迫的生理響應及抗寒性評價[J].浙江農林大學學報, 2021, 38(4): 828-836. DOI:10.11833/j.issn.2095-0756.20200606.
[6]牟開萍,李維芳,楊文新,等.20個月季品種的抗寒性綜合評價[J].草原與草坪, 2021, 41(6): 58-66. DOI:10.13817/j.cnki.cyycp.2021.06.009.
[7]賈金輝,徐凌,劉慧純,等.8個釀酒葡萄品種的抗寒性比較[J].中國果樹,2021(7):39-44.DOI:10.16626/j.cnki.issn1000-8047.2021.07.008.
[8]楊振亞,臺秀國,曹振玉,等.4個引種櫟類樹種苗期抗寒性初步研究[J].山東大學學報(理學版), 2016, 51(5): 43-47,60. DOI:10.6040/j.issn.1671-9352.0.2015.573.
[9]李素瓊,史開奇,張曉紅.夏橡白粉病防治[J].農業與技術, 2024, 44(3): 55-57. DOI:10.19754/j.nyyjs.20240215013.
[10]郭英,李開花.珍貴樹種夏橡育苗栽培技術[J].農村科技,2016(4): 69-70. DOI:10.3969/j.issn.1002-6193.2016.04.041.
[11]單興宇.夏橡的育苗和栽培技術[J].現代農機, 2021(3): 109-110. DOI:10.3969/j.issn.1674-5604.2021.03.060.
[12]楊梅花,黃祥,韓彥奇,等.8種睡蓮對低溫脅迫的生理響應及抗寒性評價[J].江蘇農業科學, 2022, 50(23): 123-128. DOI:10.15889/j.issn.1002-1302.2022.23.018.
[13]井俊麗,劉銘瀟,魏欣,等.幾種蘋果中間砧的抗寒性評價[J].果樹學報, 2022, 39(6): 970-981. DOI:10.13925/j.cnki.gsxb.20210444.
[14]劉曉東,任偉嘉,何淼.苦水玫瑰和冷香玫瑰抗寒性[J].東北林業大學學報, 2012, 40(11): 28-30,34. DOI:10.3969/j.issn.1000-5382.2012.11.007.
[15]井俊麗,劉銘瀟,高美娜,等.不同蘋果中間砧枝條在越冬期間的生理特性變化及抗寒性比較[J].河北農業大學學報, 2022, 45(4): 25-31. DOI:10.13320/j.cnki.jauh.2022.0056.
[16]王睿哲,張蓓,郭鐵群,等.5個梨品種在低溫脅迫下的生理響應及抗寒性評價[J].山東農業科學, 2023, 55(2): 57-63. DOI:10.14083/j.issn.1001-4942.2023.02.008.
[17]王偉強,田新會,杜文華.甘農4號小黑麥苗期抗寒性評價[J].甘肅農業大學學報, 2023, 58(1): 46-54. DOI:10.13432/j.cnki.jgsau.2023.01.006.
[18]趙雪輝,陳雙建,成繼東,等.3個桃品種抗寒性分析研究[J].果樹資源學報, 2020,1(6): 14-19. DOI:10.16010/j.cnki.14-1127/s.2020.06.004.
[19]王金鑫,王延松,明文,等.不同果型翅果油樹的抗寒性[J]. 東北林業大學學報, 2023, 51(6): 25-32. DOI:10.13759/j.cnki.dlxb.2023.06.014.
[20]王召元,田啟航,常瑞豐,等.桃不同品種對低溫脅迫的生理響應及評價[J].中國農業大學學報, 2022, 27(2): 66-77. DOI:10.11841/j.issn.1007-4333.2022.02.07.
[21]劉敏,靳娟,阿布都卡尤木·阿依麥提,等.新疆3個鮮食棗品種的抗寒性評價[J].新疆農業科學, 2023, 60(4): 916-924. DOI:10.6048/j.issn.1001-4330.2023.04.016.
[22]劉在國,趙通,馬超,等.7個平歐雜交榛子品種的生理解剖特性及其抗寒性研究[J].干旱地區農業研究, 2023, 41(5): 138-149. DOI:10.7606/j.issn.1000-7601.2023.05.15.
[23]溫日宇,劉建霞,李順,等.低溫脅迫對不同藜麥幼苗生理生化特性的影響[J].種子, 2019, 38(5): 53-56. DOI:10.16590/j.cnki.1001-4705.2019.05.053.
[24]曹燕燕,葛昌斌,齊雙麗,等.不同冬小麥品種(系)拔節期低溫脅迫生理反應及抗寒性評價[J].江蘇農業科學, 2022, 50(17): 59-66. DOI:10.15889/j.issn.1002-1302.2022.17.010.
[25]徐功勛,周佳,呂德國,等.4個蘋果品種的抗寒性評價[J].果樹學報, 2023, 40(4): 669-679. DOI:10.13925/j.cnki.gsxb.20220359.
[26]李桂榮,連艷會,程珊珊,等.低溫脅迫對山葡萄等6個葡萄品種抗寒性的影響[J].江蘇農業科學, 2019, 47(8): 130-134. DOI:10.15889/j.issn.1002-1302.2019.08.029.
[27]戚建莉,吳文俊,姜成英,等.不同引進希臘品種油橄欖的抗寒性評價[J].經濟林研究, 2023, 41(2): 49-57. DOI:10.14067/j.cnki.1003-8981.2023.02.006.
[28]石林曌,倪天虹,裴云霞,等.湖北省主栽油茶品種苗期抗寒性評價[J].經濟林研究, 2019, 37(4): 10-19. DOI:10.14067/j.cnki.1003-8981.2019.04.002.
[29]王海鷗,高展,艾克拜爾·毛拉,等.自然越冬條件下17個葡萄砧木品種 幾個生理指標變化分析[J].經濟林研究, 2019, 37(4): 120-128. DOI:10.14067/j.cnki.1003-8981.2019.04.016.
[30]雷夢瑤,高小峰,白清敏,等.不同品種石榴枝條的抗寒性評價[J].河南農業科學, 2023, 52(6): 120-130. DOI:10.15933/j.cnki.1004-3268.2023.06.013.
[31]李桂榮,連艷會,程珊珊,等.低溫脅迫下不同無核葡萄品種抗寒性的分析[J].西南農業學報, 2018, 31(11): 2399-2406. DOI:10.16213/j.cnki.scjas.2018.11.029.
[32]孟詩原,呂桂云,張明忠,等.5種衛矛屬植物對低溫脅迫的生理響應及抗寒性評價[J].西北植物學報, 2020, 40(4): 624-634.
[33]陽妮,蔣景龍,李麗,等.3個柑橘品種幼苗對低溫脅迫的生理響應及其抗寒性評價[J].中國果樹, 2020(5): 32-39, 143. DOI:10.16626/j.cnki.issn1000-8047.2020.05.006.
(責任編輯:趙藏賞)
