砧用瓠瓜對枯萎病的抗性配合力及遺傳分析
2024-08-24 00:00:00鐘川廖建杰方華蓉陽燕娟李昌霞王鵬
中國瓜菜 2024年7期


摘" " 要:探討砧用瓠瓜對枯萎病的抗性遺傳規律,明確砧用瓠瓜種質的育種利用價值,為瓠瓜抗枯萎病育種的親本選擇和雜種后代的抗病表型預測提供參考依據。利用對瓠瓜枯萎病表現不同抗性水平的4份砧用瓠瓜材料作為親本,按Griffing完全雙列雜交方法配制雜交組合,采用苗期傷根灌注法接種瓠瓜枯萎病病原菌,統計病情指數,分析抗病配合力;采用Hayman遺傳分析方法探討抗性遺傳規律。結果表明,砧用瓠瓜枯萎病的抗性遺傳符合“加性-顯性-上位性”模型,以加性效應為主,顯性程度為部分顯性,顯性方向為減效,抗病親本中顯性基因占比較大,抗病性狹義遺傳力為77.68%;砧用瓠瓜材料H041的一般配合力為負向效應,且負向效應最大,極顯著提高雜交后代的抗病性,以其為親本配制的雜交組合具有較高的特殊配合力。因此,在枯萎病抗性育種中,應盡可能選用抗病性強的親本進行雜交育種;H041可作為瓠瓜抗枯萎病育種的骨干親本,在較早世代進行選擇更有利于配制出雜種優勢強的雜交種。
關鍵詞:瓠瓜;枯萎病;配合力;加性-顯性-上位性
中圖分類號:S642.9 " " " " " " " " " "文獻標志碼:A 文章編號:1673-2871(2024)07-029-07
Analysis on combining ability and genetics of resistance to Fusarium wilt of rooted bottle gourd
ZHONG Chuan1, LIAO Jianjie1, 2, FANG Huarong1, YANG Yanjuan1, LI Changxia1, WANG Peng1
(1. Guangxi University, Nanning 530004, Guangxi, China; 2. Guangxi Minzu Normal University, Chongzuo 532200, Guangxi, China)
Abstract: The genetic pattern of resistance to calabash Fusarium wilt of rooted bottle gourd was explored, to provide reference for parental selection of calabash Fusarium wilt resistance and phenotypes prediction in hybrid progeny. Four materials with different resistance to calabash Fusarium wilt were used as parents to formulate hybrid combinations by Griffing complete diallel hybridization method. The pathogens of Fusarium wilt was inoculated the using root perfusion at seedling stage, and the disease index was calculated to analyze the resistance and combining ability. Hayman genetic analysis method was employed to explore the inheritance of resistance. The result showed that the inheritance of resistance to calabash Fusarium wilt was consistent with the additive-dominance-epistasis model, with additive effects as the main effect, partial dominance as the degree of dominance, and reduced dominance as the direction of dominance. The resistant parents contain more dominant genes than recessive genes, and the narrow heritability of resistance was 77.68%. The general combining ability(GCA)of H041 was negative, and the negative effect was the largest among the four materials, which could significantly improve the disease resistance of the hybrid offspring. Formulating hybrid combinations with H041 as the parent have a high specific coordination ability(SCA). In summary, in the process of breeding for wilt resistance, the parents with strong disease resistance should be selected for hybrid breeding. H041 can be used as the backbone parent for the breeding for wilt resistance in bottle gourd, and selection in earlier generations is more conducive to formulating hybrids with strong heterosis.
Key words: Bottle gourd; Fusarium wilt; Combining ability; Additive-dominance-epistasis
尖孢鐮刀菌(Fusarium. oxyspoorum)為瓜類枯萎病病原菌,根據分類依據將其分為尖孢鐮刀菌葫蘆專化型(F. oxyspoorum f. sp. lagenariae)、西瓜專化型(F. oxyspoorum f. sp. niveum)、黃瓜專化型(F. oxyspoorum f. sp. cucumerinum)等8個分化型,且各分化型對瓜類蔬菜的致病性存在差異[1]。瓠瓜枯萎病具有較強的侵染力,由尖孢鐮刀菌葫蘆專化型引起,主要從瓠瓜根部傷口入侵維管束,侵染植株后,由于毒素和導管堵塞等多因素共同作用,植株的輸導機能受損,葉片從下向上逐漸變黃萎蔫,直至整株枯萎死亡[2]。當前,枯萎病是制約瓜類生產的重要土傳病害之一。在重茬和連作區域,瓠瓜枯萎病發生尤為嚴重,會造成嚴重減產甚至絕收,給生產帶來巨大的經濟損失。采用嫁接栽培對瓜類枯萎病進行綠色防控是較為常用的手段,因此抗枯萎病的砧用瓠瓜選育是有效解決瓜類枯萎病危害問題的現實需要。瓠瓜根系發達,高抗黃瓜專化型、西瓜專化型等多種尖孢鐮刀菌,甚至達到免疫水平,廣泛用作黃瓜[3]、西瓜[4-5]、苦瓜[6]等瓜類作物的嫁接砧木。但是,不同瓠瓜對尖孢鐮刀菌葫蘆專化型的抗性水平差異較大,生產上應篩選抗葫蘆專化型枯萎病的瓠瓜作為瓜類作物的嫁接砧木。因此,探究瓠瓜抗枯萎病遺傳規律,分析雜交組合的配合力水平,對瓠瓜抗枯萎病雜交育種的親本選配具有重要意義。
在實際生產中,雜交育種的關鍵是精準選擇優良親本,雜交后代表型取決于自交系親本,配合力極其重要。配合力是預測雜交組合優劣的依據,測定親本一般配合力,利于有目的地選擇適宜配合力的親本進行雜交育種,可提高定向育種效率,更快更好地實現育種目標,避免盲目配制浪費試驗時間和成本。根據配合力篩選雜交親本,廣泛運用于甜瓜[7]、苦瓜[8]、西瓜[9]、黃瓜[10]和籽用西葫蘆[11]等瓜類育種中,但主要集中在以產量和品質為目標性狀的育種中,抗病育種相關的研究報道仍然較少。目前關于瓜類抗枯萎病的遺傳規律仍不明確,因抗原材料、分析方法不同而呈現遺傳多樣化的特點,限制了瓜類作物抗枯萎病遺傳育種研究進程和在實際生產中的應用。有關黃瓜抗枯萎病黃瓜專化型的遺傳規律研究,包括單基因顯性[12]、單基因隱性、寡基因和數量性狀遺傳[13]、多基因調控[14]、“兩對加性-顯性-上位性主基因+加性-顯性多基因(MX2-ADI-AD)”混合遺傳模型[15]等。西瓜枯萎病抗性遺傳規律符合“加性-顯性”模型,由多個隱性基因共同調控[16]。甜瓜抗枯萎病可能由單基因或多基因控制,Chikh-Rouhou 等[17]研究證明甜瓜枯萎病抗性由隱性多基因調控;陳渝文等[18]發現甜瓜枯萎病抗性是由2對主基因控制的數量性狀,遺傳規律符合“兩對加性-顯性-上位性主基因+加性-顯性-上位性多基因”模型;而趙玉龍[19]則認為甜瓜枯萎病抗性為單基因顯性遺傳。在苦瓜抗枯萎病遺傳特性研究中,趙秀娟等[20]認為苦瓜抗枯萎病受單一顯性核基因控制;而劉子記等[21]的研究則發現苦瓜抗枯萎病由兩對主基因共同調控,符合“加性-顯性-上位性主基因+加性-顯性多基因”遺傳模型,呈現數量性狀遺傳的特征;琚茜茜等[22]發現苦瓜枯萎病抗性遺傳符合-上位“MX2-ADI-AD”模型,由主基因和多基因共同控制,兩對主基因的遺傳顯性度不同,分別為超顯性和不完全顯性。筆者所在團隊的研究表明,瓠瓜抗枯萎病為多基因共同調控的數量性狀[23],遺傳規律也符合“MX2-ADI-AD”模型,由2對加性-顯性-上位性主基因+加性-顯性多基因控制[24]。
抗病砧用瓠瓜已廣泛用于瓜類嫁接栽培,但主要針對非葫蘆專化型的其他瓜類枯萎病,有關瓠瓜抗葫蘆專化型枯萎病的抗性配合力研究較少,很大程度制約了瓠瓜抗枯萎病基因定位研究和育種工作的開展。筆者選擇對瓠瓜枯萎病表現不同抗病水平的砧用瓠瓜做親本,進行完全雙列雜交,鑒定親本及其雜種一代(F1)的抗病水平,分析抗性配合力及遺傳規律,探討砧用瓠瓜對枯萎病的抗性遺傳規律,明確砧用瓠瓜種質的育種利用價值,為瓠瓜抗枯萎病育種的親本選擇、雜種后代的抗病表型預測提供理論依據,也為抗病基因定位研究奠定基礎。
1 材料與方法
1.1 材料
試驗于2017-2018年在廣西大學農學院試驗基地進行。供試砧用瓠瓜4份種質為廣西大學農學院蔬菜團隊經高代自交得到的純合自交系,編號分別為H02(果實近圓形,感瓠瓜枯萎病)、H07(果實梨形,中抗瓠瓜枯萎病)、H041(果實梨形、抗瓠瓜枯萎病)、 H12(果實梨形,抗瓠瓜枯萎病)。采用Griffing完全雙列雜交設計,獲得正反交12個F1代種子。
供試的病原菌為尖孢鐮刀菌葫蘆專化型,從廣西瓜類蔬菜產區南寧市五塘鎮的瓠瓜枯萎病發病區域采集得到,通過分離純化和保存作為接種源。
1.2 方法
病原菌接種采用苗期傷根灌注法,參考農業行業標準《植物新品種特異性、一致性和穩定性測試指南 瓠瓜》[25]鑒定抗病表型,具體操作步驟參考廖建杰[23]的方法。菌液濃度 1×106 CFU·mL-1,每株灌注10 mL菌液。各親本和F1分別接種42株,3次重復。根據病情指數(DI)判定抗病水平,分為抗病(R,DI ≤25)、中抗(MR,25lt;DI ≤55)、感病(S,DI gt;55)。
DI=∑(各病級發病植株數×相對病級數值)/(供試株數×3)×100。
1.3 數據處理
采用DPS7.05軟件進行數據處理,各親本和F1的病情指數經反正弦轉換后,按照Griffing完全雙列雜交方法分析配合力和Hayman遺傳分析方法的模型Ⅰ進行抗病遺傳分析[26]。
2 結果與分析
2.1 病情指數分析
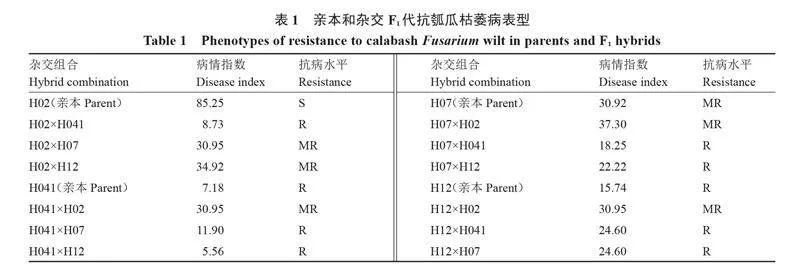
4個砧用瓠瓜親本及12個雜交組合的瓠瓜枯萎病抗性鑒定結果如表1 所示。7個F1表現高抗(R),病情指數在5.56~24.60之間,其中雜交組合H041×H12的抗病性最強,病情指數最低(5.56),其次為雜交組合H02×H041,病情指數為8.73;5個F1表現中抗(MR),病情指數在30.95~37.30之間。對病情指數進行方差分析,結果(表2)表明,處理間p =0.000 1lt;0.01,差異極顯著。說明親本間的抗病遺傳基礎存在極顯著差異,親本及雜交組合抗枯萎病表型差異顯著,可進行配合力和遺傳分析。
2.2 抗病性配合力分析
2.2.1 總體配合力分析 使用Griffing法進行抗病性配合力分析。結果表明,一般配合力(GCA)和特殊配合力(SCA)均存在極顯著差異(p=0.000 1lt;0.01),且F值比值大于1(GCA/SCA=1 666.135/237.708=7.01),說明瓠瓜抗枯萎病表型受加性效應和非加性效應共同影響,且加性效應大于非加性效應,即以加性效應為主(表3)。反交效應(reciprocal effect)與一般配合力和特殊配合力相同,也達到了極顯著差異水平(p=0.000 1lt;0.01),說明瓠瓜抗枯萎病除了受加性效應和非加性效應的影響外,還受細胞質基因的影響,且表現出核質基因互作效應。
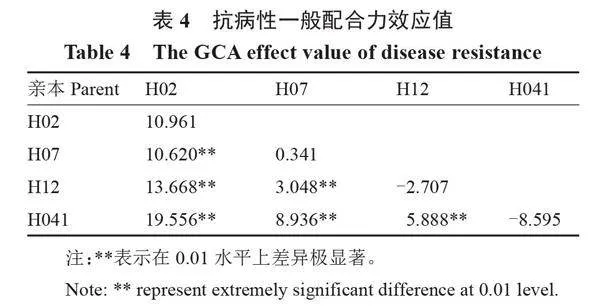
2.2.2" " "一般配合力效應分析" " 4個砧用瓠瓜親本對瓠瓜枯萎病抗性的一般配合力效應值如表4所示,在4份(H02、H07、H12和H041)親本材料中,枯萎病抗性的一般配合力效應值在-8.595~19.556之間,其中感病親本H02和中抗親本H07的一般配合力均為正值,表現為正向效應,說明以感病材料H02和中抗材料H07為親本進行雜交不利于高抗病性雜交種的獲得;抗病親本H041、H12的一般配合力均為負值,即表現負向效應,說明用其作為親本配制雜交組合易獲得高抗病性的雜交種。此外,各親本之間雜交的效應值均呈極顯著差異,推測這4個砧用瓠瓜親本之間的內在基因型不同。
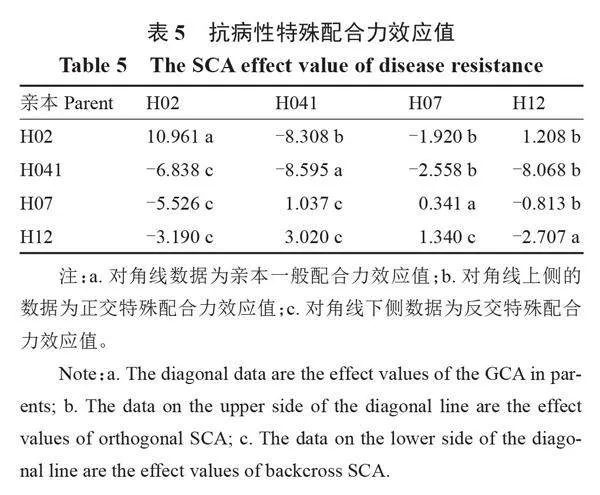
2.2.3" " 特殊配合力效應分析" " 對4個砧用瓠瓜親本進行枯萎病抗性特殊配合力效應分析,結果如表5所示,以H02、H041、H07和H12為親本,雜交得到的12個F1代中,相同親本雜交所得F1的特殊配合力效應值差異較大,說明特殊配合力在F1代中起不同作用,表現較為復雜。其中,在6個正交組合中,僅以感病親本H02為母本的1個組合(H02×H12)的特殊配合力效應值為正值(1.208),表現正向效應,其余5個組合均表現負向效應;在6個反交組合中,以H02為父本的3個組合(H041×H02、H07×H02、H12×H02)的特殊配合力效應值均為負值,表現為負效應,其余3個組合均表現正向效應。以抗病親本H041為父本或母本的雜交組合H02×H041、H041×H12的特殊配合力效應值分別為-8.308、-8.068,表現負向效應,抗病雜種優勢較強。以上結果表明,母本細胞質基因參與調控抗瓠瓜枯萎病表型,在抗枯萎病雜交育種中,應選擇抗病性較強的材料作為母本,同時考慮親本的一般配合力與特殊配合力效應,因此H041可作為抗瓠瓜枯萎病雜交育種的骨干親本。
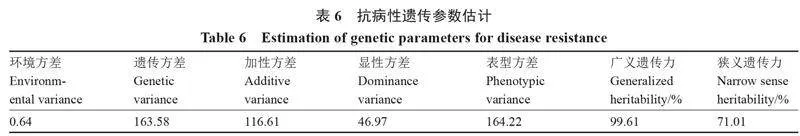
2.2.4" " 遺傳參數估計" " 為評估砧用瓠瓜的抗病性遺傳表現,進行遺傳參數估計(表6),加性方差(116.61)大于顯性方差(46.97),表明砧用瓠瓜抗瓠瓜枯萎病受加性和顯性效應共同作用,且加性效應占主導地位。而砧用瓠瓜抗瓠瓜枯萎病的廣義遺傳力達到99.61%,狹義遺傳力為71.01%,表明本研究中供試的砧用瓠瓜親本抗瓠瓜枯萎病的表型對環境不敏感。
2.3 抗病性遺傳分析
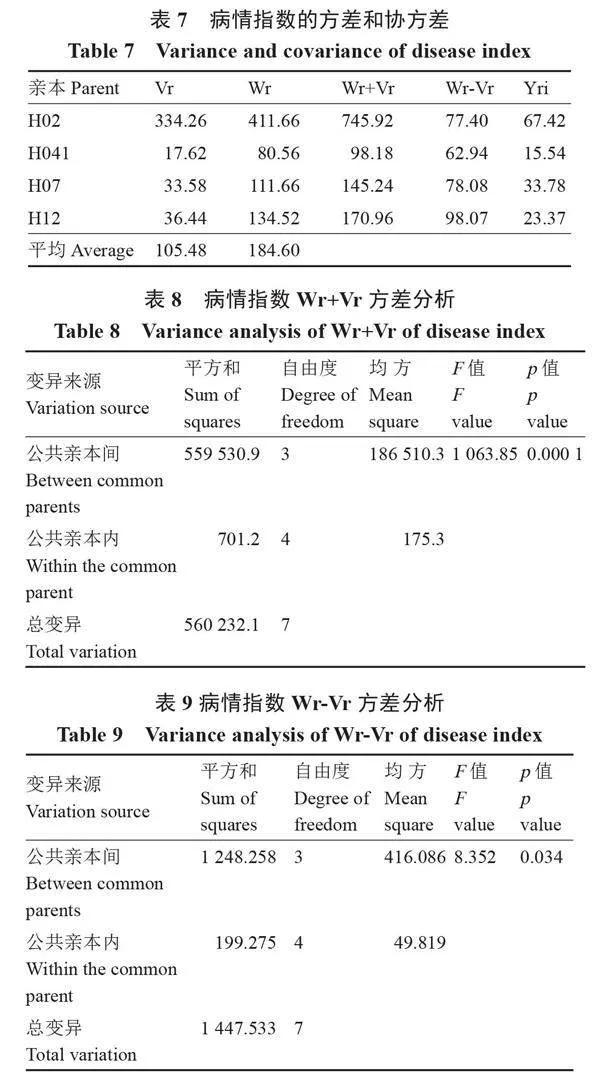
2.3.1" " 基因互作及顯隱性分析" " 通常認為,非等位基因的互作即上位性作用引起行列協方差與行列方差之差(Wr-Vr)的多樣性;等位基因的互作即顯性高差引起行列協方差與行列方差之和(Wr+Vr)的多樣性。對病情指數的方差和協方差(表7)進行Wr+Vr和Wr-Vr方差分析(表8、表9),結果表明,Wr+Vr呈極顯著差異,Wr-Vr呈顯著差異,說明砧用瓠瓜親本間存在顯性效應和上位性作用,且顯性效應大于上位性作用,抗病遺傳機制復雜。
進行Wr+Vr與病情指數平均值(Yri)相關性分析,結果表明,r=0.959 (plt;0.05),二者呈顯著正相關,說明病情指數的遺傳表現為高值受隱性基因控制,低值受顯性基因控制,顯性基因起減效作用,即Wr+Vr值低的親本可能存在更多顯性調控基因。由此得出,親本H041含有更多調控抗枯萎病的顯性基因,可作為抗瓠瓜枯萎病雜交育種的優勢骨干材料。
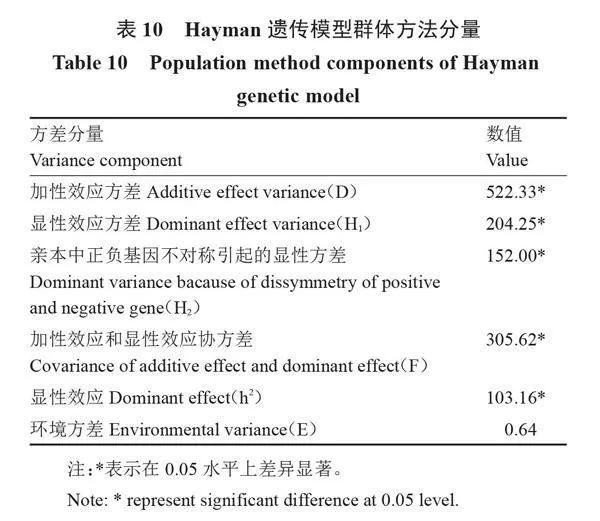
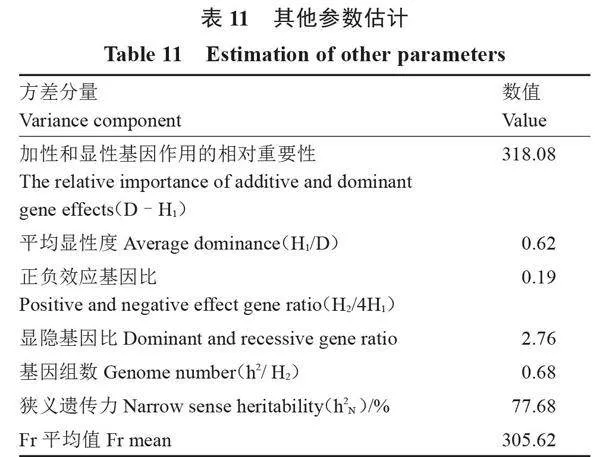
2.3.2" " 方差分量估算" " 對瓠瓜抗枯萎病遺傳參數進行估算,結果如表10和表11所示,病情指數的加性方差(D)與顯性方差(H1、H2)均達到顯著水平,表明遺傳機制中顯性效應和加性效應起關鍵作用。加性和顯性基因作用的相對重要性(D-H1)=318.08gt;0,進一步證明遺傳機制存在加性效應和顯性效應,且加性效應大于顯性效應,以加性效應為主。平均顯性度(H1/D=0.625)小于1,表明親本抗病性不存在超顯性遺傳,表現出部分顯性。正負效應基因比(H2/4H1=0.19)小于最大值0.25,表明顯性位點上的基因分布不對稱,即親本增效和減效基因分布不同。親本顯隱基因比為2.76,進一步說明親本中顯性基因和隱性基因數量不同,且顯性基因數量多于隱性基因。F1顯性效應(h2)值為103.16,達到顯著水平,且砧用瓠瓜抗枯萎病顯性基因組數小于1,說明至少有1對純合顯性基因控制砧用瓠瓜枯萎病抗性。加性和顯性效應協方差Fr的平均值F=305.62gt;0,達到顯著水平,說明抗病親本中顯性等位基因多于隱性等位基因,與顯隱性基因比相符。抗病性狹義遺傳力為77.68%,說明雜交后代的抗性水平受親本影響,因此后續的育種工作應在較早世代進行選擇,更有利于配制出雜種優勢強的雜交種。
3 討論與結論
通過構建若干雜交家系,利用統計模型估算一般配合力及其他遺傳參數,進一步全面了解該性狀的遺傳特性,有利于快速選擇適宜的親本,從而進行雜交得到優良雜交種[27]。為了適應生產的需要 ,近幾年瓠瓜利用雜種優勢選育雜種一代抗病新品種的研究在逐步開展,但相關報道仍然極少。筆者所在課題組前期針對砧用瓠瓜雜交種開展了西瓜枯萎病[5]和瓠瓜枯萎病[23]的抗病雜種優勢研究,選用優勢親本配制出的F1代均表現較強抗病雜種優勢。龐兆良等[28]培育出瓠瓜砧木的雜交種雪峰強砧1號 ,表現出與西瓜嫁接親和性強,高抗枯萎病,適宜在我國南方西瓜重茬連作區進行嫁接栽培。然而,我國對于瓠瓜育種的研究起步相對較晚,且大部分集中在生理特性方面 ,而目前市場上瓠瓜品種的抗病性是育種的關鍵問題。因此,加強瓠瓜兼抗或高抗主要病害的育種工作,并緊密結合市場受歡迎的品質、產量等性狀進行研究,是目前研究的工作重點。
抗病砧用瓠瓜已廣泛用于各種瓜類嫁接栽培,不僅高抗瓠瓜枯萎病,也對西瓜[29]、黃瓜[30]枯萎病具有較強的抗性。此外,由于瓜類枯萎病病原菌具有多種分化型,抗性材料的遺傳背景及抗性鑒定方法也有所差異,因此不同瓜類抗枯萎病的遺傳規律各不相同,具有復雜性。研究表明,瓜類枯萎病抗性遺傳由多基因控制,西瓜抗枯萎病遺傳符合“加性-顯性”模型[16],具有主基因和微效基因共同控制的特點[31];冬瓜抗枯萎病遺傳符合“加性-顯性”模型[32];甜瓜抗枯萎病遺傳符合“兩對加性-顯性-上位性主基因+加性-顯性多基因”混合遺傳模型[18]。此外,也有相關報道證明瓜類枯萎病抗性由單基因控制,在苦瓜中,單一顯性核基因控制枯萎病抗性[20];黃瓜枯萎病抗性受單個顯性基因控制[12, 33];在西瓜中,也存在單基因調控枯萎病抗性的研究內容[34-35]。由于瓜類抗枯萎病遺傳規律具有復雜性,制約了快速精準開展抗枯萎病育種工作的進行,探究瓠瓜抗枯萎病的遺傳特性,對科學制定育種策略具有重要的指導意義。本研究結果表明,砧用瓠瓜抗枯萎病遺傳符合“加性-顯性-上位性”遺傳模型,以加性效應為主,并且存在非等位基因互作效應,研究結果與黃瓜抗枯萎病[30]和西瓜抗枯萎病[36]的研究結果存在共同點,抗病性由顯性基因控制,不存在超顯性遺傳,且表現為部分顯性和存在細胞質基因互作;與冬瓜抗枯萎病基因顯隱性及遺傳特點略有差異[32],其原因可能是研究材料和枯萎病分化型不同。
瓜類作物的抗病性通常是由多個基因共同調控的復雜數量性狀,因此進行數量性狀的遺傳研究對抗病育種工作具有重要意義。筆者所在課題組前期進行瓠瓜枯萎病抗性遺傳研究時,首先采用卡方檢驗分析得出瓠瓜抗枯萎病符合數量性狀遺傳特點,由多基因調控[23],然后通過“主基因+多基因混合遺傳模型”分析得出抗病遺傳符合“MX2-ADI-AD”模型,由 2 對主基因共同調控[24],這與黃瓜[15]、苦瓜[21-22]篩選出的最佳遺傳模型類似,證明了瓠瓜枯萎病抗性受 2 對主基因共同調控,主基因存在加性效應和顯性效應,且加性效應大于顯性效應。
綜上所述,筆者采用 Hayman 完全雙列雜交分析法深入研究瓠瓜枯萎病抗性遺傳規律,結果表明,瓠瓜抗枯萎病遺傳符合“加性-顯性-上位性”模型;對親本及雜交組合進行一般配合力及其他遺傳參數分析,發現顯性方向指向減效,說明抗病親本中顯性基因數量多于隱性基因,參與調控抗病且表現出顯性效應的純合基因至少有1對。此外,抗病性狹義遺傳力為77.68 %,表明加性效應遺傳的重要性,砧用瓠瓜對枯萎病的抗性能穩定傳遞給子代,并且在雜種的早期世代進行選擇更有利于篩選出抗性強的雜交組合。研究結果為瓠瓜抗枯萎病雜交的親本選擇提供了參考,為發掘和驗證抗病基因功能奠定了研究基礎。
參考文獻
[1]" " " 佘小漫,何自福,羅方芳,等.瓜類枯萎病及其綜合防治技術[J].廣東農業科學,2011(2):84-87.
[2]" " " 靳穎玲,郭利敏.瓜類枯萎病研究進展[J].蔬菜,2023(1):26-29.
[3]" " " 費雨蘭,王晶,沈佳,等.不同砧木嫁接對黃瓜長勢及果實品質的影響[J].江蘇農業科學,2013,41(12):147-149.
[4]" " " 邢乃林,張蕾琛,應泉盛,等.嫁接對葫蘆砧木及西瓜接穗甲基化水平的影響[J].江西農業學報,2016,28(9):27-30.
[5]" " " 陳文明,鐘川,廖建杰,等.雙砧木嫁接對西瓜生長及果實品質的影響[J].中國瓜菜.2017,30(7):13-16.
[6]" " " 廖道龍,陳貽誦,胡艷平,等.海南夏秋季苦瓜嫁接砧木篩選試驗[J].蔬菜,2018(5):18-23.
[7]" " " 沈佳,壽偉松,張躍建.甜瓜果實主要數量性狀的配合力分析[J].中國瓜菜,2017,30(12):4-8.
[8]" " " 吳立東,劉亞婷,鐘金仙,等.苦瓜親本配合力、遺傳距離與雜種優勢的相關性分析[J].上海農業學報,2020,36(3):30-35.
[9]" " " 攸學松,朱莉,曾劍波,等.西瓜砧木自交系產量與品質性狀配合力分析[J].園藝與種苗,2021,41(8):23-26.
[10]" " 王慧,臧柯昕,陳勁楓,等.歐洲溫室型黃瓜品質性狀雜種優勢及配合力分析[J].南京農業大學學報,2022,45(3):483-492.
[11]" " 楊永升,王萍,于曉婧,等.籽用西葫蘆主要性狀配合力、遺傳力及雜種優勢分析[J].內蒙古農業大學學報(自然科學版),2022,43(3):19-23.
[12]" " MATSUMOTO Y,MIYAGI M.A single dominant gene confers resistance to Fusarium oxysporum f.sp. melonis race 1 in West Indian Gherkin (Cucumis anguria L.) accessions[J].South African Journal of Botany,2012,83:117-120.
[13]" " VAKALOUNAKIS D J,LAMPROU K.The Foc gene governs resistance to race 3 of Fusarium oxysporum f.sp. cucumerinum in the cucumber cv.SMR-18[J].European Journal of Plant Pathology,2018,152(3):653-656.
[14]" " 周紅梅,毛愛軍,張麗蓉,等.黃瓜枯萎病接種方法及抗性遺傳的研究[J].華北農學報,2010,25(4):186-190.
[15]" " DONG J P,XU J,XU X W,et al.Inheritance and quantitative trait locus mapping of Fusarium wilt resistance in cucumber[J].Frontiers in Plant Science,2019,10:1425.
[16]" " 羊杏平,姚懷蓮,劉廣,等.西瓜品種枯萎病抗性的遺傳研究[J].江蘇農業學報,2008,24(6):882-887.
[17]" " CHIKH-ROUHOU H,GONZALEZ-TORRES R,OUMOULOUD A,et al.Inheritance of race 1.2 Fusarium wilt resistance in four melon cultivars[J].Euphytica,2011,182(2):177-186.
[18]" " 陳渝文,趙榮茺,沈慕潔,等.甜瓜抗枯萎病的遺傳分析[J].分子植物育種,2022,20(12):4044-4050.
[19]" " 趙玉龍.甜瓜橙色果肉、全雌系及主要抗病性狀MAS的建立與評價[D].哈爾濱:東北農業大學,2017.
[20]" " 趙秀娟,唐鑫,胡開林.苦瓜枯萎病抗性鑒定與抗性遺傳規律研究[J].園藝學報,2013,40(4):685-692.
[21]" " 劉子記,朱婕,牛玉,等.苦瓜枯萎病抗性材料Thai4-6的遺傳模型分析[J].熱帶作物學報,2018,39(8):1501-1506.
[22]" " 琚茜茜,黃如葵,黃熊娟,等.苦瓜種質枯萎病抗性鑒定與抗性遺傳特征分析[J/OL].分子植物育種,[2024-03-20].https://link.cnki.net/urlid/46.1068.S.20240319.1127.002.
[23]" " 廖建杰.砧用瓠瓜對瓠瓜枯萎病的抗性遺傳分析及嫁接應用研究[D].南寧:廣西大學,2019.
[24]" " 趙榮茺,沈慕潔,張歸,等.瓠瓜枯萎病抗性的遺傳規律分析[J/OL].分子植物育種,[2024-03-10].https://kns.cnki.net/kcms/detail/46.1068.S20230310.1349.004.html.
[25]" " 全國植物新品種測試標準化技術委員會.植物新品種特異性、一致性和穩定性測試指南 瓠瓜:NY/T 2504-2013[S].北京:中國標準出版社,2013.
[26]" " 蘭巨生.作物遺傳參數統計法[M].石家莊:河北人民出版社,1982.
[27]" " 李加納.數量遺傳學概論[M].重慶:西南師范大學出版社,2007.
[28]" " 龐兆良,莫小平,羅華,等.瓠瓜砧木新品種‘雪峰強砧1號’的選育[J].中國瓜菜,2020,33(5):66-68.
[29]" " 陳文明,鐘川,廖建杰,等.砧用瓠瓜種質資源對西瓜枯萎病的抗性及嫁接適用性研究[J].中國蔬菜,2017(6):25-31.
[30]" " 樂素菊,陳珉,吳定華,等.黃瓜枯萎病抗性遺傳規律研究[J].安徽農業大學學報,2014,41(2):270-272.
[31]" " 鄒小花,張海英,李勝,等.野生西瓜種質PI296341-FR抗枯萎病菌生理小種2的遺傳規律[J].園藝學報,2011,38(9):1699-1706.
[32]" " 謝大森,何曉明,彭慶務,等.冬瓜枯萎病的抗性遺傳規律[J].熱帶作物學報,2009,30(7):1005-1008.
[33]" " 毛愛軍,張峰,張麗蓉,等.黃瓜品系WIS2757對黃瓜枯萎病生理小種4和黑星病的抗性遺傳與連鎖分析[J].中國農業科學,2008,41(10):3382-3388.
[34]" " 姚懷蓮.西瓜枯萎病抗性遺傳及生理生化基礎研究[D].江蘇揚州:揚州大學,2007.
[35]" " 劉廣,黃曉云,徐錦華,等.西瓜枯萎病抗性分子標記篩選與應用[J].江蘇農業科學,2022,50(18):279-283.
[36]" " PAL S,RAO E S,SRIRAM S.Genetic analysis of resistance to Fusarium oxysporum f.sp.niveum race 2 in cultivated watermelon [Citrullus lanatus (Thunb.) Matsum amp; Nakai][J].Australasian Plant Pathology,2020,49(3):319-326.
收稿日期:2024-04-19;修回日期:2024-05-28
基金項目:國家自然科學基金項目(31660568);廣西自然科學基金項目(2023GXNSFAA026508);廣西科技重大專項(桂科 AA17204039-2,AA17204026-1)
作者簡介:鐘" 川,女,助理研究員,研究方向為蔬菜遺傳育種。E-mail:825964063@qq.com
通信作者:王 " 鵬,男,副研究員,研究方向為蔬菜遺傳育種。E-mail:wangpeng@gxu.edu.cn om
