巴西橡膠樹GRAS基因家族鑒定及表達分析
2024-10-09 00:00:00李娟袁淵吳挺開鄧治程漢
熱帶作物學報 2024年9期

關鍵詞:巴西橡膠樹;GRAS 基因家族;生物信息學;基因表達;DELLA
中圖分類號:S794.1 文獻標志碼:A
轉錄因子(transcription factor, TF)是功能基因組學的核心,它是一種能與特異DNA 序列結合的蛋白,可以單獨或與其他蛋白形成復合體,提高或阻斷特定基因RNA 聚合酶的招募,調控基因的表達[1]。轉錄因子在調控高等植物的生長發育和非生物脅迫響應中起重要作用[2]。已知的轉錄因子包括GRAS、WRKY、AP2/EREBP、bZIP、MYB、MADS 和bHLH 等。
GRAS家族是一類植物特有的轉錄因子,其命名來源于該家族最早鑒定的3 個功能特征基因GAI(gibberellic-acid insensitive)、RGA(repressorof GA1-3 mutant)、SCR(scarecrow)[3-5]。GRAS蛋白的長度為400~770 個氨基酸不等,具有1 個高度保守的C-端GRAS 結構域。GRAS 結構域約390 個氨基酸,由5 個不同基序按照特定順序排列組成:LHR I(leucine heptad repeat I)、VHIID、LHR II(leucine heptad repeat II)、PFYRE 和SAW[6]。其中,VHIID 基序與2 個亮氨酸七肽重復區( LHR I 和LHR II ) 結合形成LHRI-VHIID-LHR II 復合體,該結構可能在蛋白-DNA或蛋白-蛋白互作中發揮關鍵功能[7]。PFYRE 和SAW 基序功能仍未闡明,但這2 個基序發生缺失或錯義突變導致擬南芥(Arabidopsis thaliana)表型異常[4, 8]。GRAS 蛋白的N 端保守性較差,但一些GRAS 家族成員在N 端區域含有保守的序列,如DELLA 亞家族包含2 個保守的基序DELLA 和TVHYNP[9]。最初根據擬南芥和水稻的研究將GRAS 家族分為DELLA、HAM(hairy meristem)、PAT1(phytochrome a signal transduction 1)、LAS(lateral suppressor)、SHR(short root)、SCR、SCL3(SCR- like 3)、LISCL(lilium longiflorumSCR-like)等8 個亞家族。而其他植物物種,如番茄(Solanum lycopersicum)[10]、茶樹(Camelliasinensis)[11]和葡萄(Vitis vinifera)[12]中有13 個亞家族,葫蘆(Lagenaria siceraria)[13]中有16 個亞家族,CENCI 等[14]將被子植物GRAS 家族分為17 個亞家族。目前,GRAS 基因家族已在多個物種中被報道,其中擬南芥(Arabidopsis thaliana)[15]有33 個成員、水稻(Oryza sativa)[15]有60 個、玉米(Zea mays)[16]有86 個、大白菜(Brassica rapassp. pekinensis)[17]有48 個、毛果楊(Populustrichocarpa ) [18] 有106 個、中國薔薇( Rosachinensis)[19]有56 個成員,大戟科的木薯(Manihotesculenta)[20]有77 個、麻風樹(Jatropha curcas)[21]和蓖麻(Castor bean)[22]均有48 個成員。
GRAS基因家族在植物生長發育、激素信號傳導、逆境響應等多個生物學過程中發揮著重要的調控作用。如GRAS 家族成員DELLA 蛋白作為赤霉素(gibberellin, GA)信號轉導途徑的負調控因子,通過與其他蛋白互作從而介導GA 調控植物生長[23]。此外,DELLA 還參與調控茉莉酸(jasmonic acid, JA)信號轉導[24]及次生細胞壁形成[25-26];PAT1 作為光敏色素A 信號途徑的正調控因子參與光信號途徑調控[27];HAM 參與莖尖分生組織的生長發育[28];SCR 和SHR 通過形成復合體進而共同調控擬南芥根和芽的徑向生長[29];SCL3 參與調控陸地棉根的伸長[30];在大豆中過表達GmGRAS37 基因可以增強植株的耐旱和耐鹽能力[31]。近年來,隨著對GRAS 基因的深入研究,越來越多的證據表明,GRAS 基因家族在植物的適應性進化和環境適應性方面起著關鍵的作用[32-34]。
巴西橡膠樹(Hevea brasiliensis),簡稱橡膠樹,原產于巴西亞馬遜河流域馬拉岳西部地區,是大戟科橡膠樹屬一種典型的熱帶雨林樹種,也是我國及世界熱區的一種重要經濟作物。橡膠樹所產生的膠乳是天然橡膠的主要原料。天然橡膠是重要的戰略物資和工業原料,盡管世界上有2000 多種產膠植物,如銀膠菊[35]、橡膠草[36]和杜仲[37]等,但目前所使用的天然橡膠約98%仍來源于橡膠樹[38]。隨著橡膠樹全基因組測序工作的完成,為基因家族的全基因組分析提供了便利,也為橡膠樹功能基因組學研究提供了可靠的基因組數據信息[39-41]。基于GRAS 基因家族在植物生長發育和逆境脅迫響應中的重要作用,本研究利用生物信息學方法,以橡膠樹基因組為基礎,鑒定橡膠樹GRAS 基因家族成員,并對其系統進化、基因結構、染色體位置和啟動子順式作用元件等進行分析,利用轉錄組數據及qPCR 技術分析GRAS 基因表達模式,從而為橡膠樹GRAS 基因功能的解析提供理論依據。
1 材料與方法
1.1 材料
本研究所采用的試驗材料均來自于中國熱帶農業科學院試驗場(海南省儋州市)種植的橡膠樹(Hevea brasiliensis)熱研73397 品種。采集橡膠樹不同組織(雌花、雄花、木質部、葉片、樹皮和膠乳),不同節間木質部(選取橡膠樹頂端分生組織從上往下數第一、第三和第五節間木質部),以及150 mg/L 外源GA3 處理21周后(每周噴灑處理1次)的橡膠樹幼苗的木質部(以H2O處理作為對照)樣品,立即置于液氮中,帶回實驗室后于–80 ℃中保存備用。每5 株樹為1 個生物學重復,每個樣品3個生物學重復。
1.2 方法
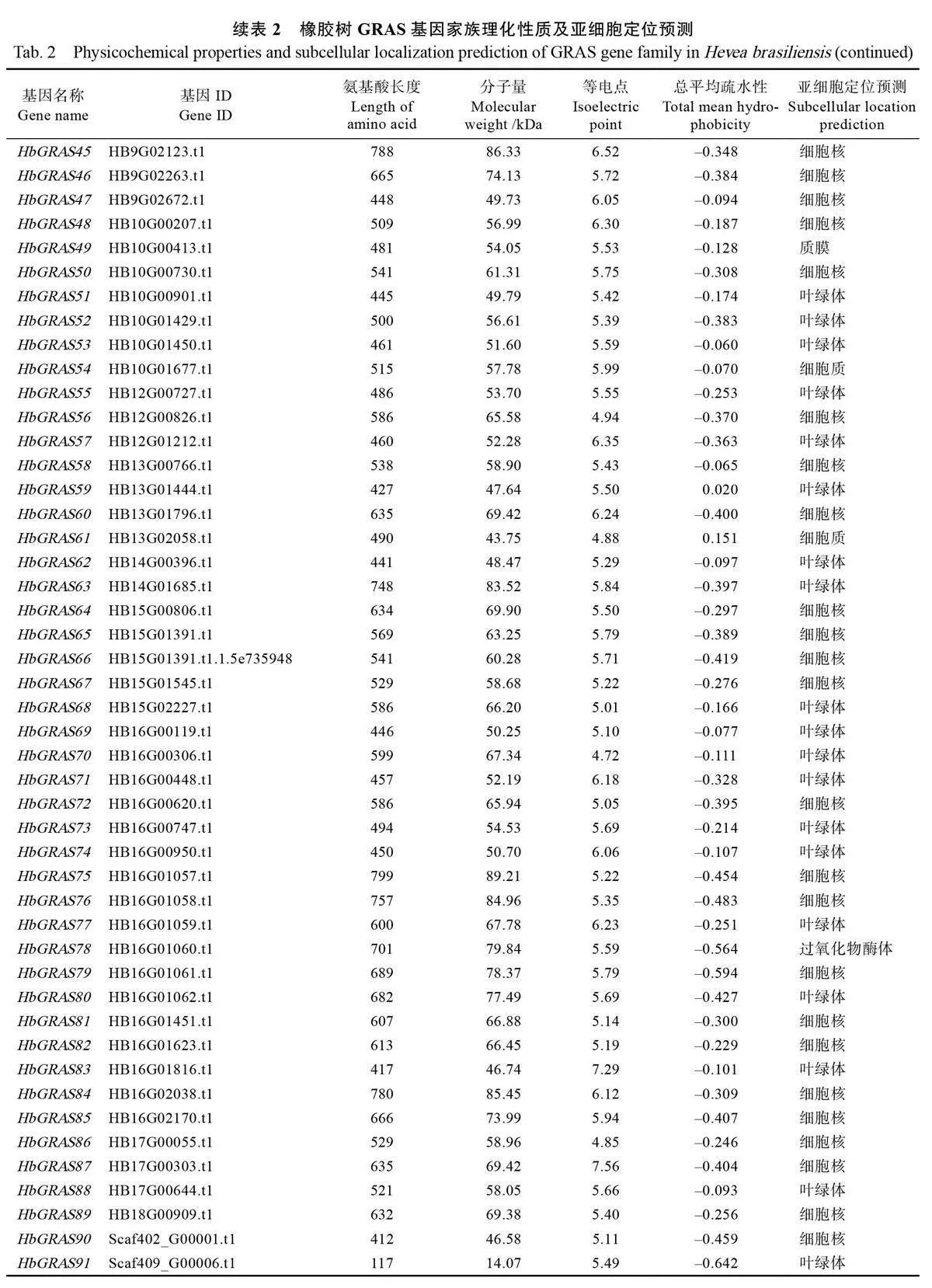
1.2.1 橡膠樹GRAS家族成員的鑒定及理化性質分析 從NCBI數據庫(https://www.ncbi.nlm.nih.gov/)中下載橡膠樹基因組序列、蛋白質序列和注釋信息。利用Pfam 數據庫(http://pfam-legacy.xfam.org/)下載的GRAS 結構域(PF03514)和隱馬爾可夫模型(HMM)文件,通過hmmer 3.0軟件篩選橡膠樹基因組數據庫中含有GRAS 結構域的全部蛋白序列,選取e 值小于1e–5 的候選蛋白序列。使用ClustalX 2.1 軟件將候選蛋白序列進行一一比對去除冗余,并通過SMART(https://smart. embl.de/)和NCBI-CDD 數據庫(https://www.ncbi. nlm.nih.gov/Structure/cdd/cdd.shtml)鑒定其保守結構域,去除不含GRAS 結構域的序列,得到橡膠樹GRAS 家族成員。分別利用ExPASyProtParam( https://web.expasy.org/protparam/)和Plant-mPLoc ( http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/)在線軟件進行氨基酸長度、分子量大小、等電點、疏水性等理化性質和亞細胞定位預測。
1.2.2 GRAS 的系統進化分析 通過PlantTFdb(http://planttfdb.gao-lab.org/index.php)數據庫和NCBI 數據庫下載擬南芥、毛果楊和水稻GRAS家族蛋白序列,并與橡膠樹GRAS 蛋白序列進行多序列比對。利用MEGA 11 軟件,采用鄰接法(neighbor-joining method, NJ)構建系統進化樹,bootstrap 設置為1000 以測試進化樹的可靠性。利用Evolview(https://www.evolgenius.info/evolviewv2/#login)在線軟件和AI 軟件進行結果美化。
1.2.3 橡膠樹GRAS基因結構分析 根據橡膠樹基因組注釋信息,利用在線網站MEME(https://meme-suite.org/meme/)與NCBI Batch CD-search( https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi)在線軟件進行motif 預測和保守結構域分析,其中motif 數量設置為10,其他參數均為默認值。通過使用TBtools 軟件對橡膠樹GRAS家族的外顯子-內含子結構進行分析,并將系統進化樹、蛋白保守基序、保守結構域和基因結構進行可視化展示。
1.2.4 橡膠樹GRAS 基因染色體定位分析 利用TBtools軟件在橡膠樹基因組注釋信息的基礎上,將橡膠樹GRAS 基因定位到染色體上,并作共線性分析,同時利用該軟件計算其Ka/Ks 值。
1.2.5 橡膠樹GRAS 基因啟動子順式作用元件分析 選取橡膠樹GRAS 轉錄起始位點上游2000 bp的序列作為啟動子分析區域,利用PlantCARE( https://bioinformatics.psb.ugent.be/webtools/plantcare/html/)在線軟件進行啟動子順式作用元件預測,并通過TBtools 進行可視化展示。
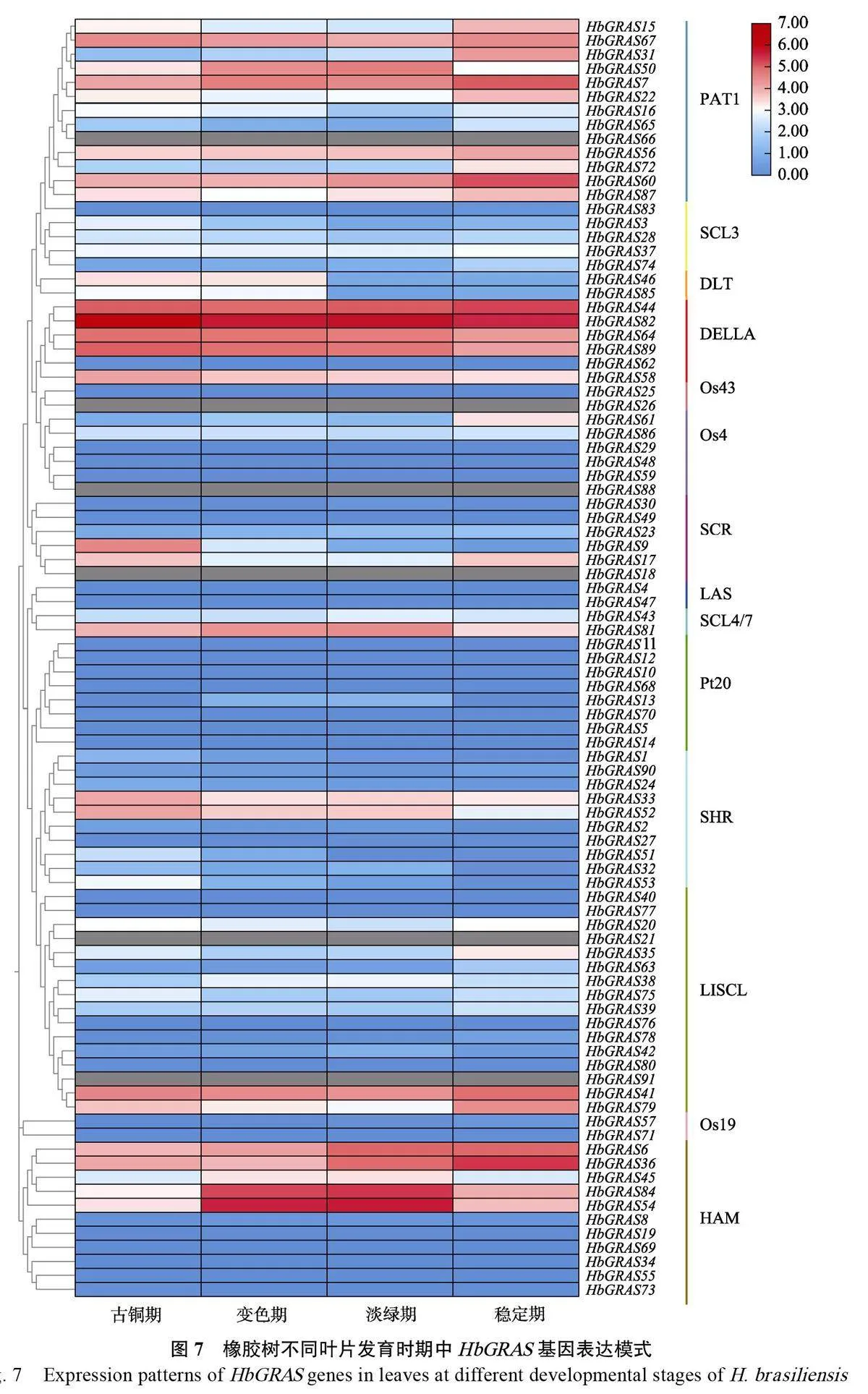
1.2.6 基于RNA-Seq 數據的基因表達分析 利用課題組前期獲得的RNA-Seq 數據,以及HeveaDB(http://hevea.catas.cn/home/index)數據庫中公布的轉錄數據,獲得不同組織(膠乳、樹皮、葉片、雌花、雄花和木質部)及葉片不同發育時期(古銅期、變色期、淡綠期和穩定期)的橡膠樹GRAS 家族成員的轉錄本FPKM(fragments perkilobase of exon per million reads mapped)值,并使用TBtools 軟件對橡膠樹GRAS 基因表達水平進行聚類及熱圖繪制。
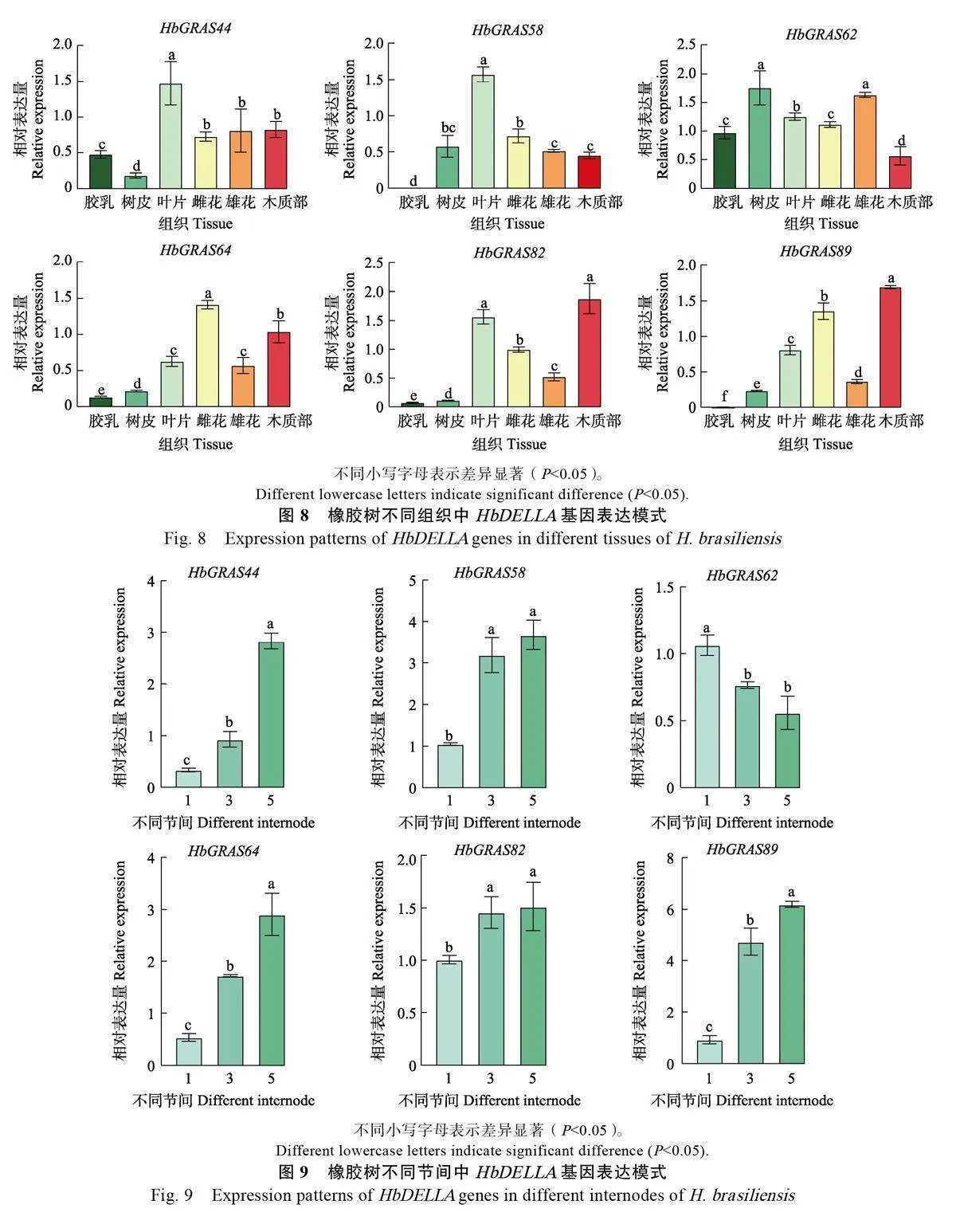
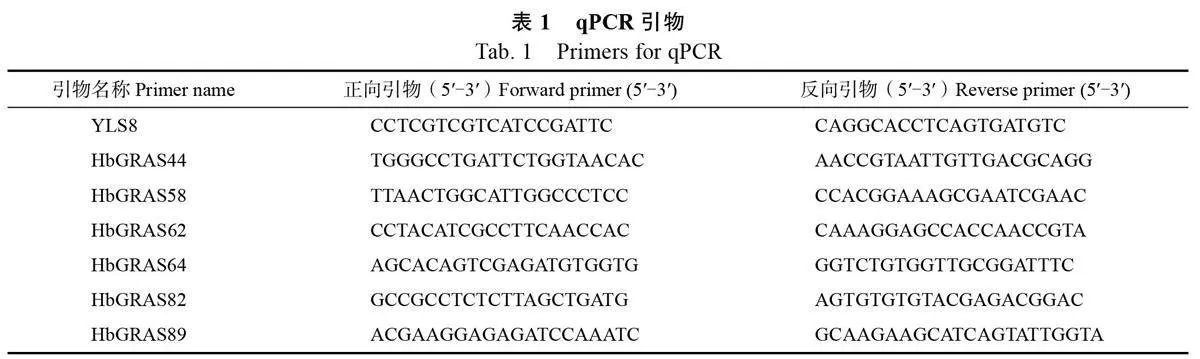
1.2.7 qPCR 分析 對橡膠樹DELLA 亞家族6個成員組織的轉錄組表達數據進行qPCR 驗證,并分析其在不同節間木質部及GA3處理下的表達模式。利用天根生化科技(北京) 有限公司的RNAprep Pure 多糖多酚植物總RNA 提取試劑盒(TIANGEN, Beijing, China)提取樣品總RNA。采用NanoDrop 2000分光光度計和凝膠電泳檢測RNA 樣品的濃度和完整性。將1 μg RNA 樣品用PrimeScript? RT reagent Kit with gDNA Eraser 試劑盒(Takara, Dalian, China)反轉錄為cDNA。采用Primer 3軟件設計HbGRAS 基因的qPCR 引物(表1),以YLS8 為內參基因。將上述反轉錄得到的cDNA 作為模板,利用2×Q3 SYBR qPCRMaster Mix(Universal)(TOLOBIO, China)進行qPCR,反應體系與擴增程序按照說明書進行。采用2–ΔΔCT 法計算基因的相對表達水平。本實驗包括3個生物學重復和3個技術重復。
2 結果與分析
2.1 橡膠樹GRAS基因家族成員鑒定與理化性質分析
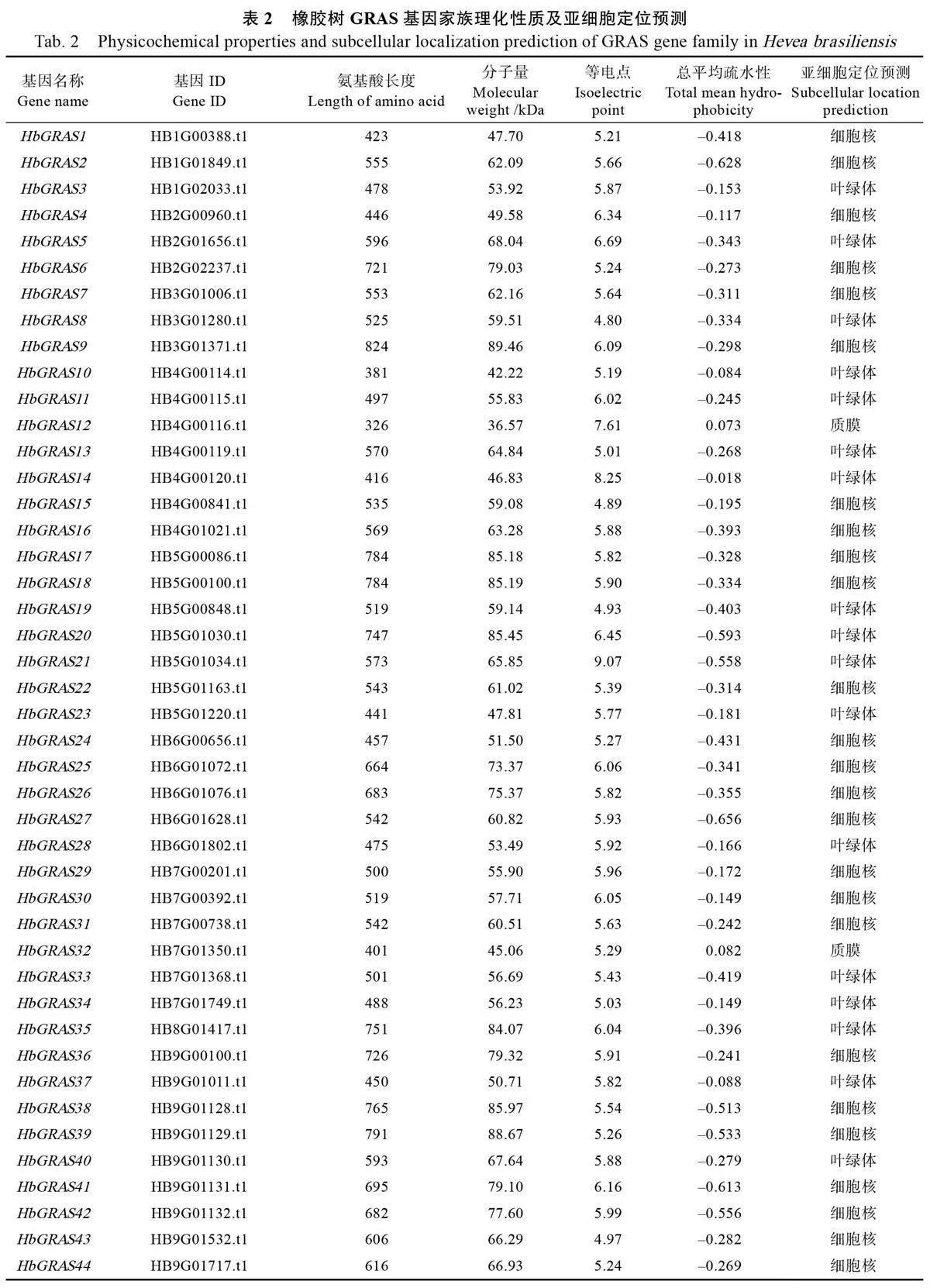
根據NCBI 數據庫中下載的橡膠樹蛋白質序列及隱馬爾可夫模型,結合SMART 和NCBI-CDD數據庫篩選和驗證,最終得到91 個非冗余的橡膠樹GRAS 基因家族成員,根據其在染色上的位置將其命名為HbGRAS1~HbGRAS91(表2)。理化性質分析結果顯示,橡膠樹GRAS 基因編碼的蛋白長度在117~824 個氨基酸之間,其中HbGRAS91的氨基酸序列最短,HbGRAS9 最長。蛋白分子量在14.07~89.46 kDa 之間,等電點在4.72(HbGRAS70)~9.07(HbGRAS21)之間,平均值為6.90。蛋白總平均疏水性在–0.656~0.151 之間,其中,除HbGRAS12 、HbGRAS32 、HbGRAS59 和HbGRAS61 的總平均疏水性大于0 之外,其余皆小于0,說明橡膠樹GRAS 家族成員大部分為親水蛋白。蛋白亞細胞定位預測結果顯示,有49 個HbGRAS 蛋白定位于細胞核,36 個定位于葉綠體,3 個定位于質膜,2 個定位于細胞質,1 個定位于過氧化物酶體。
2.2 橡膠樹GRAS 基因家族系統進化分析
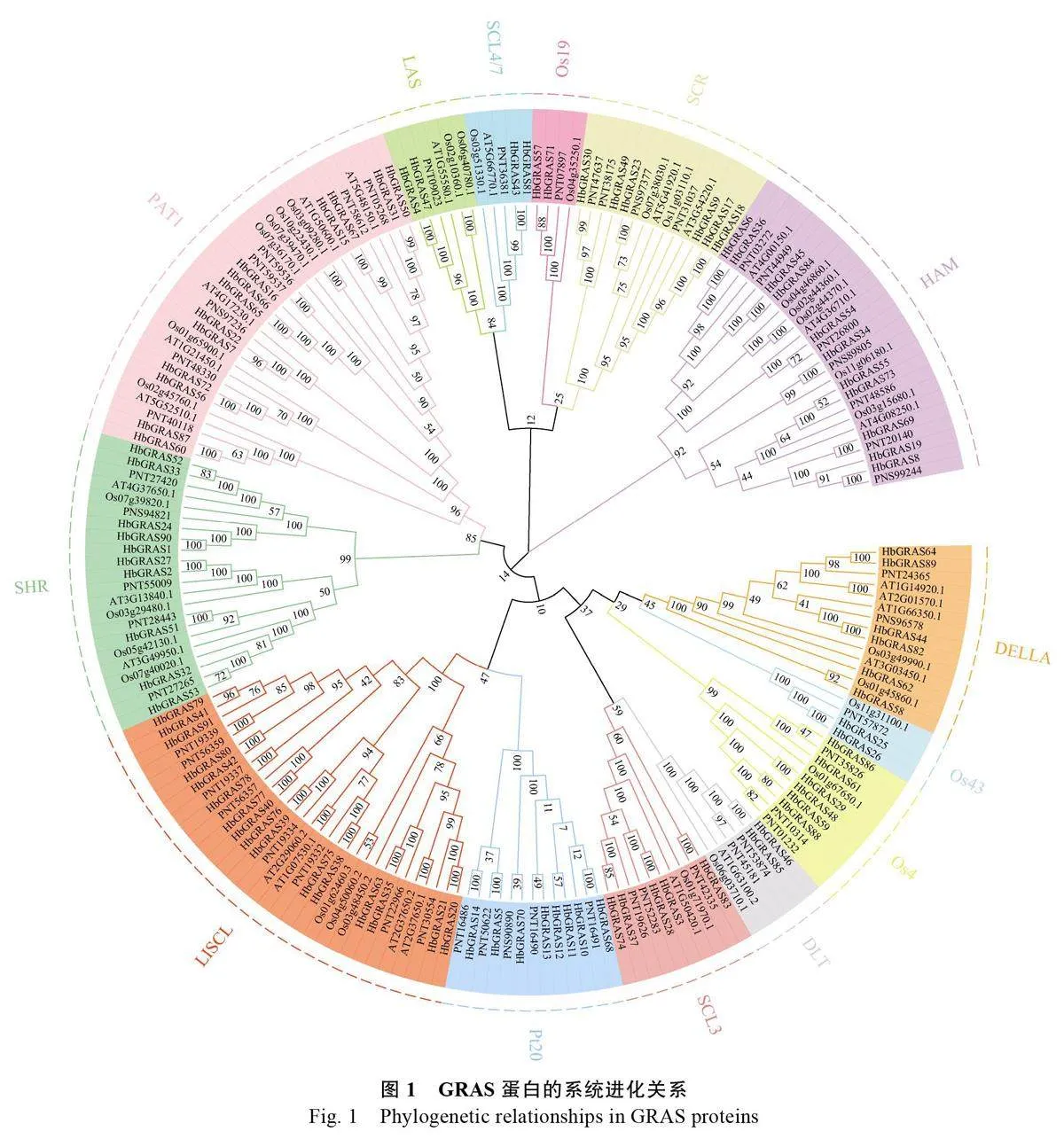
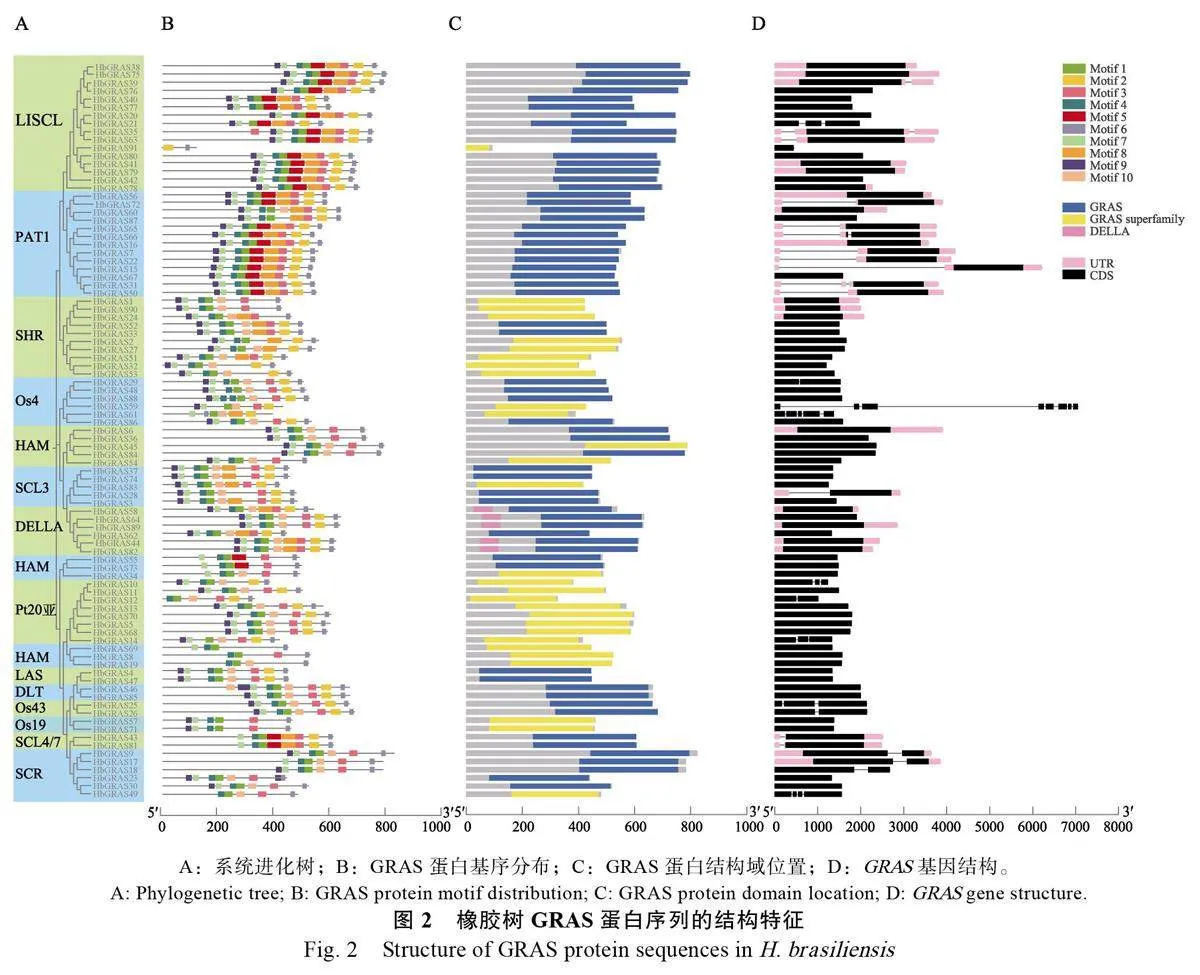
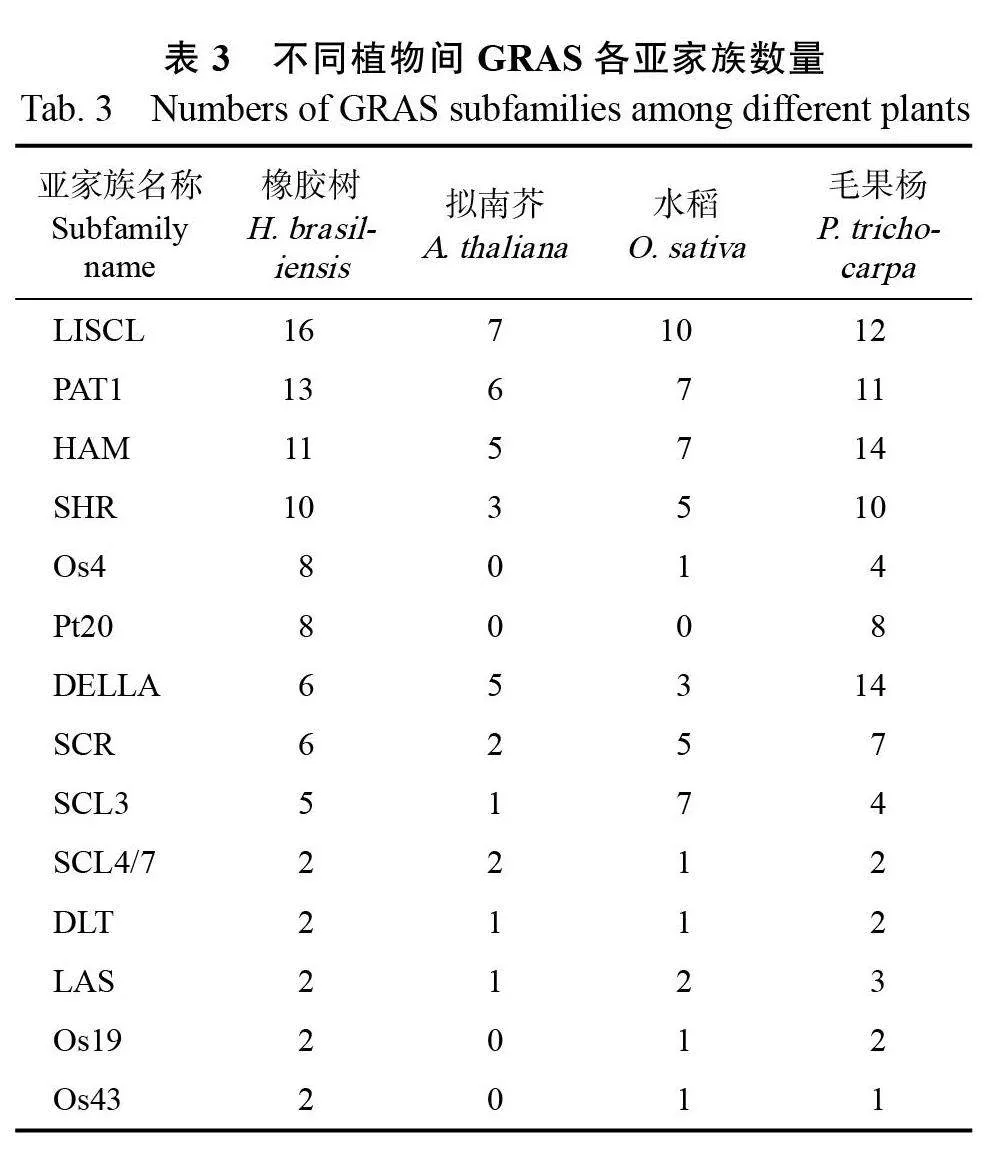
為解析橡膠樹GRAS 蛋白的特征,分別從PlantTFdb 和NCBI 數據庫中下載擬南芥、毛果楊和水稻GRAS 家族蛋白序列,與本研究鑒定得到的91 個橡膠樹GRAS 家族成員的蛋白序列進行比對及系統進化分析。結果表明,GRAS 蛋白被劃分為14 個亞家族,分別是Os19、HAM、SCL4/7、LAS、SCR、DLT、SCL3、Os4、Os43、DELLA、Pt20、LISCL、SHR 和PAT1(圖1)。其中LISCL亞家族的成員數目最多有16 個,PAT1 亞家族有13 個,HAM 亞家族有11 個,SHR 亞家族有10個,Pt20 亞家族有8 個,Os4、SCR 和 DELLA亞家族各有6 個,SCL3 亞家族有5 個,Os19、Os43、SCL4/7、LAS 和DLT 各亞家族僅有2 個成員。比較不同植物之間GRAS 各亞家族的數量發現,橡膠樹、擬南芥和水稻中LISCL 亞家族的數量最多,分別為16、7 和10 個。毛果楊中DELLA和HAM 亞家族成員數量最多,均為14 個。Pt20亞家族僅存在于橡膠樹和毛果楊中,而擬南芥無Os4、Os43 和Os19 亞家族成員(表3,圖2A)。
2.3 橡膠樹GRAS家族保守結構域和基因結構分析
保守基序分析發現,大多數motifs 位于序列C 端,motif 6存在于全部HbGRAS 蛋白中,且始終在C 端。motif 1 大多與motif 4、motif 5 或者motif 10相鄰。同一亞族的HbGRAS 成員通常具有相似的基序組成。例如, LISCL 亞族除HbGRAS91只含有motif 2 和motif 6 外,其余均包括motif 9、motif 7、motif 1、motif 8、motif 5、motif 3、motif 2、motif 6,DELLA 亞族均含有motif 9、motif 7、motif 4、motif 1、motif 10、motif3、motif 2、motif 6,HAM 亞家族成員僅含有4個保守的motifs(圖2B)。
保守結構域分析結果顯示,HbGRAS成員均含有GRAS 或GRAS superfamily 結構域,除HbGRAS91外其他均位于C端,而DELLA結構域僅存在于DELLA 亞家族中。同一亞家族的成員其基因結構相似,如PAT1 亞家族僅有GRAS結構域,Pt20 亞家族僅有GRAS superfamily 結構域。DELLA 亞家族中除HbGRAS62J1JlAwCy+K8obl18ie46cA==外,其余均含有DELLA 保守結構域(圖2C)。
基因結構多樣性是基因家族進化的重要組成部分[42]。橡膠樹GRAS 基因具有0~7 個數量不等的內含子,其中63 個HbGRAS 無內含子,18 個HbGRAS 僅有1 個內含子,HbGRAS59 含有7 個內含子,數量最多。DELLA、LAS、Os19、HAM和DLT 亞家族無內含子,Os4/7、SCL3 和SHR亞家族含有0~1 個內含子,Os4 亞家族含有0~7個內含子(圖2D)。總體來說,系統進化中同一亞家族的橡膠樹GRAS 基因具有相似的外顯子-內含子結構。
2.4橡膠樹GRAS基因染色體定位和基因復制分析
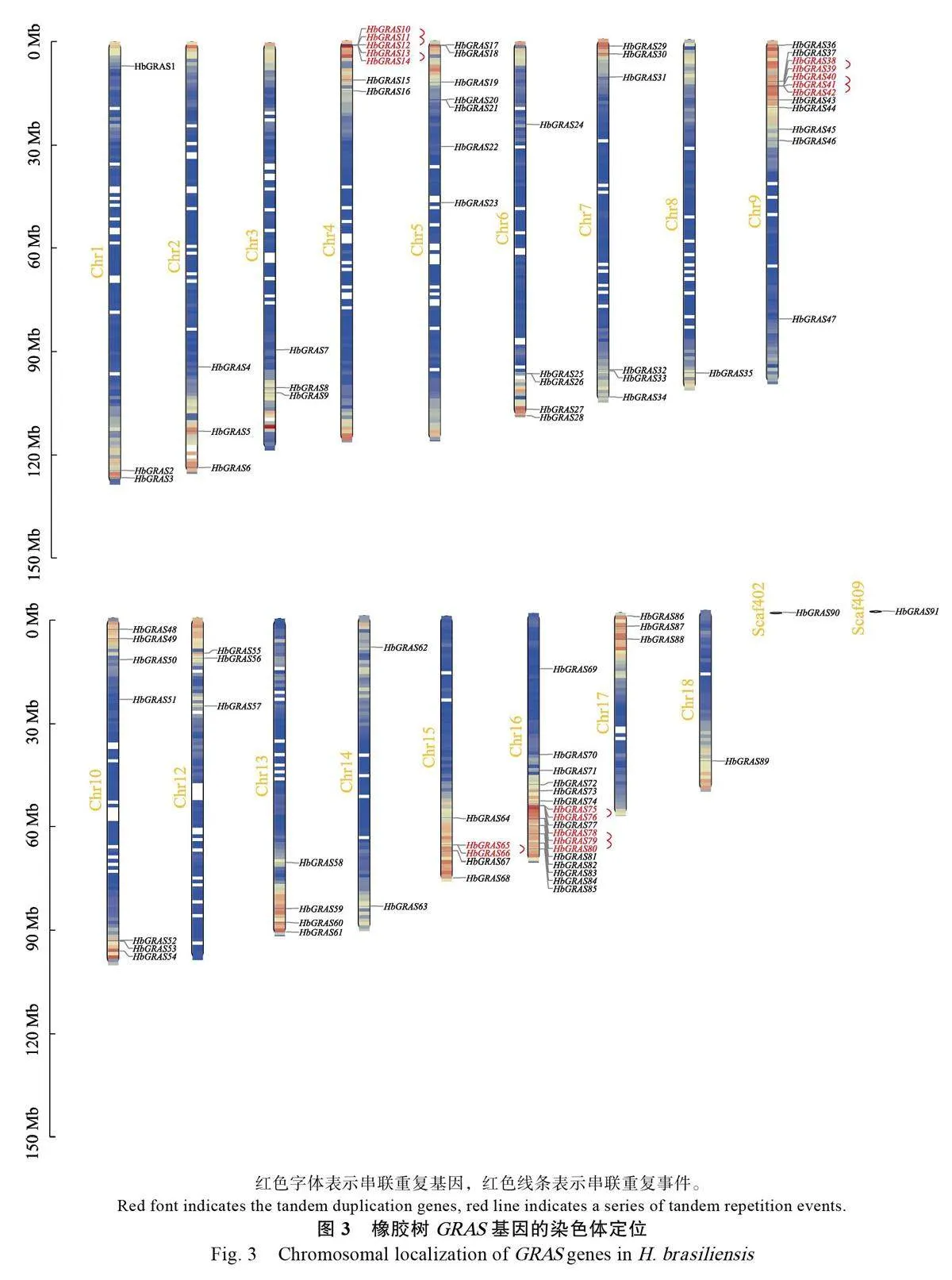
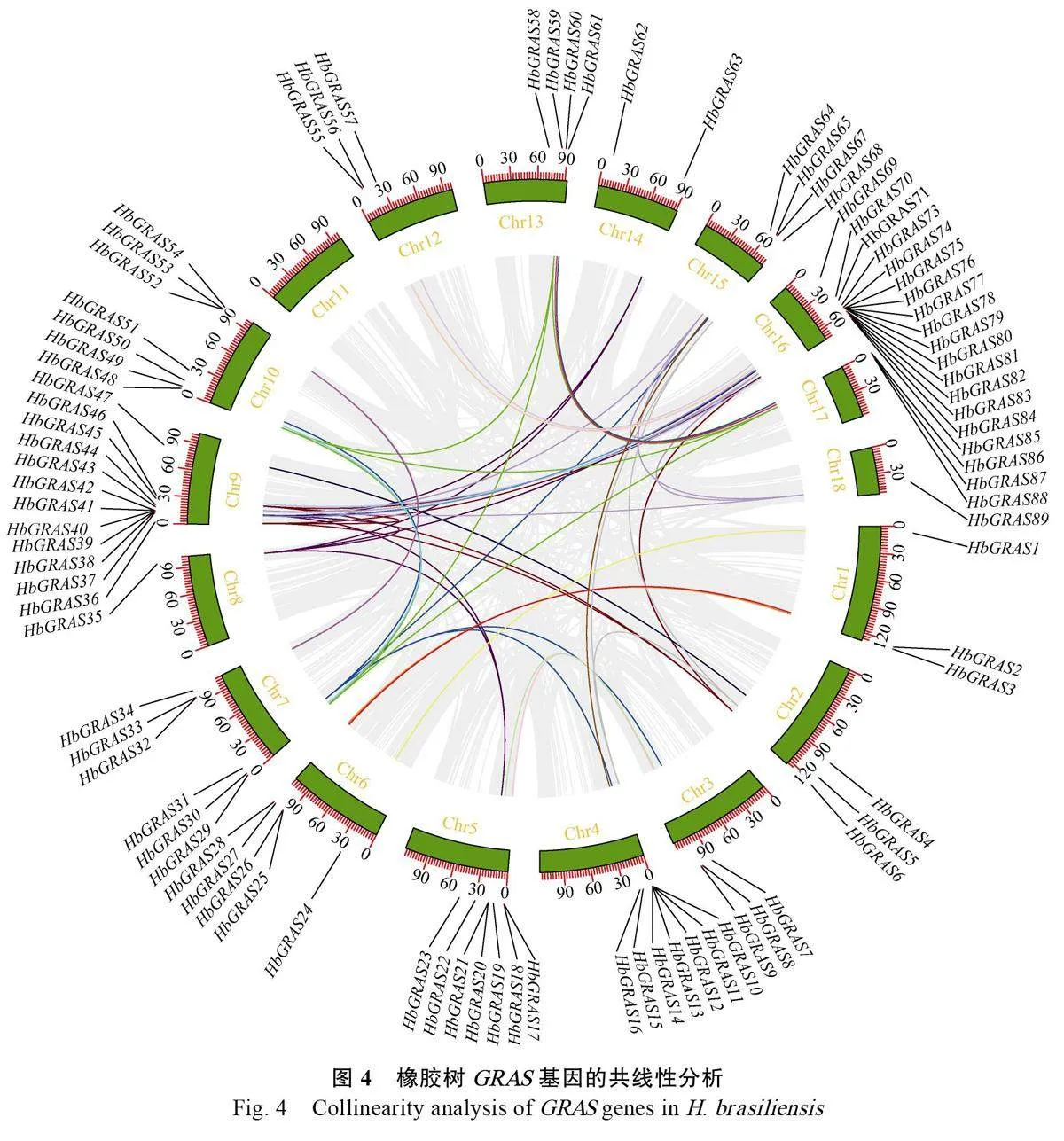
橡膠樹GRAS基因的染色體定位如圖3所示,91個HbGRAS基因分布于除11號染色體外的其他17條染色體和2條Scaffold上,并且每個染色體上分布的數目不同。16號染色體上分布最多,有17個基因,約占總數的18.68%,其次是9號染色體分布有12個基因,而8和18號染色體,以及Scaffold402和Scaffold409分布最少,各自僅含1個HbGRAS基因。
串聯重復事件是指200kb內包含2 個或更多基因的染色體區[43],在基因組進化、調控和穩定性等方面具有重要的作用[44]。在橡膠樹4、9、15和16號等4條染色體上共發現17 個HbGRAS基因的10個串聯重復事件(圖3)。共線性分析顯示,橡膠樹GRAS 基因片段重復區域中發現了87個基因(圖4),說明片段重復事件可能是橡膠樹GRAS 基因家族成員進化的主要原因。HbGRAS家族的同源基因對的Ka/Ks 值在0.091~0.388 之間,說明這些基因經過片段復制后發生了純化選擇。
2.5橡膠樹GRAS基因啟動子順式作用元件分析
啟動子區順式作用元件分析結果顯示,橡膠樹GRAS 家族主要包含植物激素應答順式作用元件,如GA、茉莉酸甲酯(methyl jasmonate, MeJA)、脫落酸(abscisic acid, ABA)、生長素(auxin)和水楊酸(salicylic acid, SA)響應元件,脅迫應答順式作用元件,如傷害、干旱和低溫響應元件,以及參與分生組織、胚乳、種子等時空表達及光響應的順式作用元件等。其中,光響應元件在全部91 個HbGRAS 基因中都存在且數量最多,有1111個。其次,是茉莉酸甲酯響應元件有178 個,厭氧誘導元件164 個,脫落酸響應元件129 個,GA 響應元件85 個,其中數量最少的元件是光敏色素互作因子、SEF1因子結合位點和參與柵欄葉肉細胞分化的順式作用元件,各有2個(圖5)。上述結果表明,HbGRAS 可能參與多種植物激素和環境脅迫應答,在植物生長發育和逆境脅迫響應中起著重要作用。
2.6 橡膠樹GRAS基因家族成員表達分析
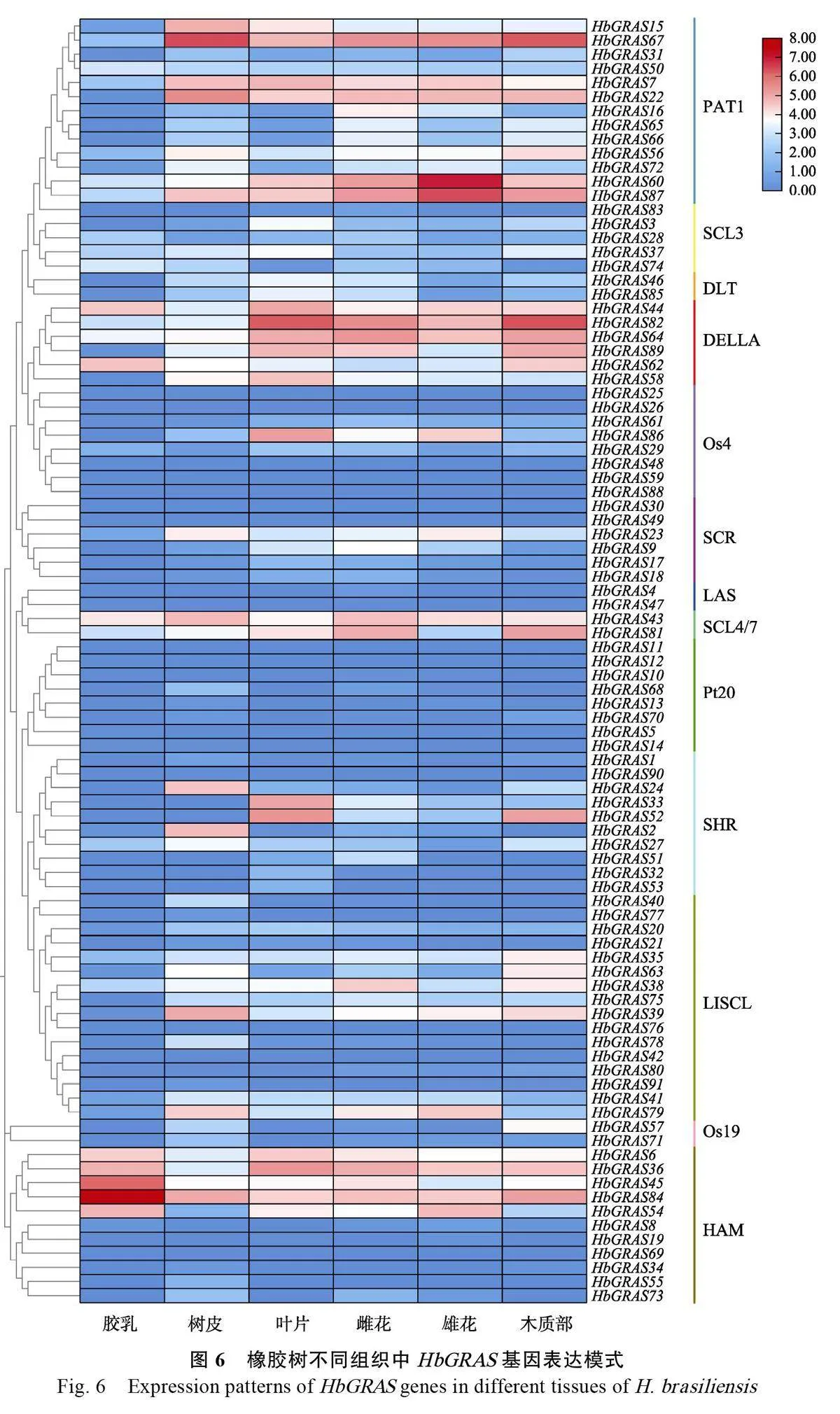
HbGRAS基因在橡膠樹膠乳、樹皮、葉片、雌花、雄花和木質部等不同組織的表達模式如圖6所示,其中HbGRAS11、HbGRAS12、HbGRAS26、HbGRAS30、HbGRAS42、HbGRAS48、HbGRAS49、HbGRAS59 和HbGRAS69等9 個基因在所有檢測組織中均不表達。HbGRAS45 和HbGRAS84 在膠乳中高表達,HbGRAS67 在樹皮和木質部中表達量較高,HbGRAS82在葉片和木質部中的表達豐度較高。雄花中表達豐度最高的是HbGRAS60,其次是HbGRAS87。不同亞族中HbGRAS 的組織表達模式也不盡相同,比如LAS 和Pt20 亞家族成員在所有組織中的表達水平都低于其他亞家族,而PAT1、DELLA 和SCL4/7 亞家族的相對表達量較其他各亞族略高。
為進一步了解HbGRAS 在橡膠樹葉片發育中的作用,從Hevea DB數據庫下載HbGRAS基因在橡膠樹葉片不同發育時期的轉錄組數據,表達模式分析結果顯示,DELLA 亞家族中HbGRAS44 隨著葉片的發育進程表達量逐漸上調,而HbGRAS58、HbGRAS64、HbGRAS82 和HbGRAS89 則逐漸下調表達;PAT1 亞家族中HbGRAS7和HbGRAS60,HAM亞家族中HbGRAS6 和HbGRAS36 均隨著葉片的成熟表達量逐漸上調。HAM 亞族中HbGRAS84和HbGRAS54 在變色期和淡綠期的表達量相對較高(圖7),說明HbGRAS 基因表達與葉片發育有關,但可能存在功能差異。
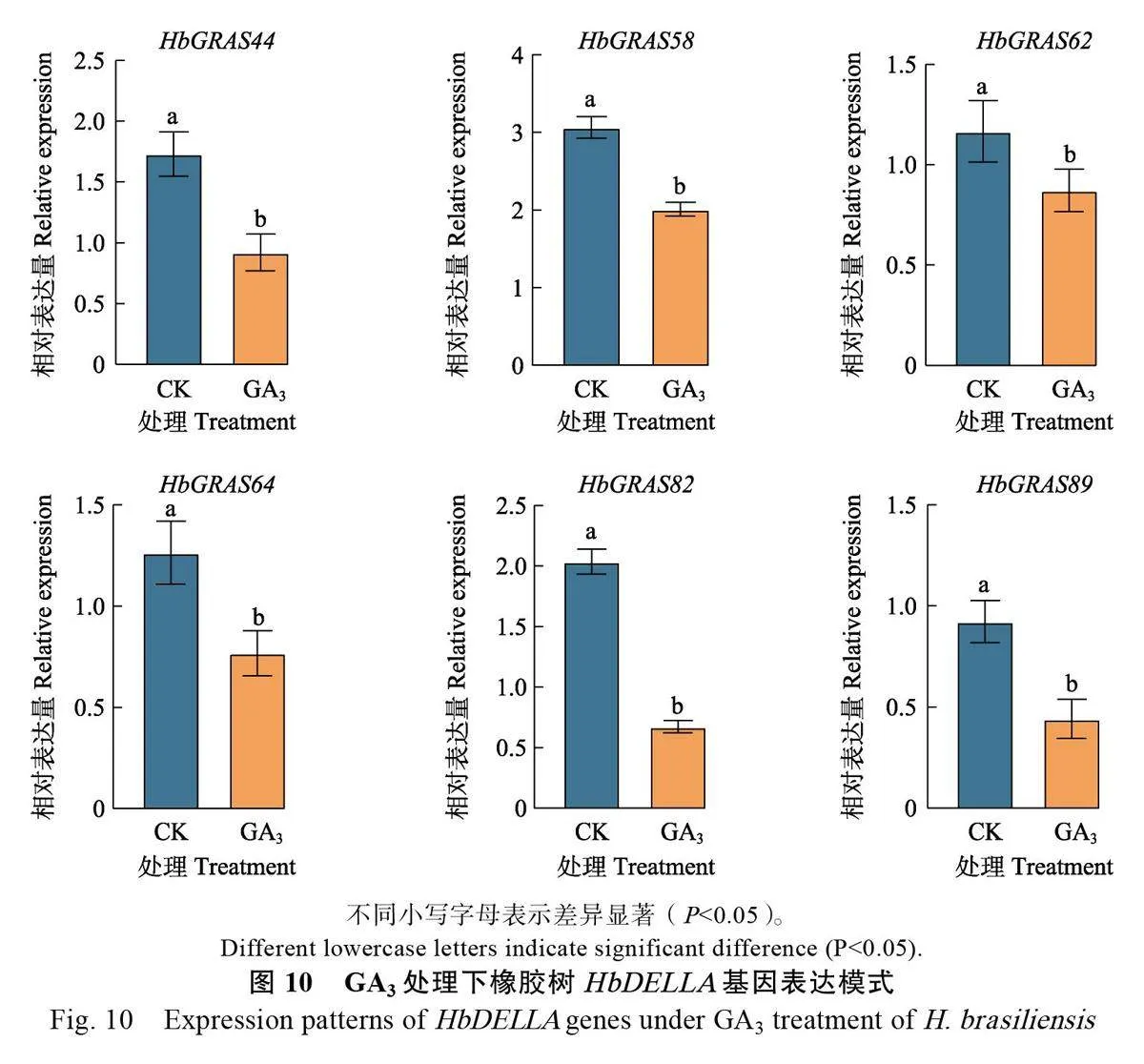
作為GRAS 家族的重要成員,DELLA 是GA信號傳導途徑的負調控因子,在植物生長發育過程中發揮重要作用[45]。因此,進一步利用qPCR技術分析橡膠樹DELLA 亞家族成員的基因表達模式。結果顯示HbGRAS62在樹皮中表達量最高,木質部中表達量最低,而 RNA-Seq結果顯示表達量最高和最低的組織分別為膠乳和雌花。其余5個DELLA基因表達量最高和最低的組織均與RNA-seq 結果一致,其中HbGRAS44分別為葉片和樹皮、HbGRAS58 分別為葉片和膠乳、HbGRAS64分別為雌花和膠乳、HbGRAS82和HbGRAS89 均分別為木質部和膠乳(圖8)。造成HbGRAS62 qPCR和RNA-Seq表達模式不一致的原因可能與2 種技術本身的差異(即轉錄組測序是對該基因的所有轉錄本定量,而qPCR則可能不能代表所有的轉錄本)有關。在不同節間木質部中,除HbGRAS62外,其余5 個橡膠樹DELLA基因均隨木質化程度加深上調表達(圖9),說明橡膠樹DELLA 基因可能參與橡膠樹莖的次生細胞壁形成。外源GA3 處理抑制DELLA 亞家族成員表達(圖10),說明DELLA基因表達受GA 調控。
3討論
GRAS作為植物中特有的一類轉錄因子,在植物的生長發育以及非生物脅迫應答中具有重要的調控作用,但目前在橡膠樹中尚無該轉錄因子家族分析相關的研究報道。本研究從橡膠樹全基因組中篩選并鑒定到HbGRAS 基因家族成員91個,所有HbGRAS 蛋白在結構上存在顯著差異,表明其高度復雜。其成員數量與擬南芥(33個)、水稻(60個)、玉米(86個)、葡萄(43個)、辣椒(50個)[46]、楊樹(106個)、蘋果(127 個)[47]等物種相比差異較大,較同科的木薯(77個)、蓖麻(48個)和麻風樹(48 個)中GRAS 基因數量明顯增加,且蛋白長度在117~824 個氨基酸之間變化,這種較大的差異可能與基因組大小或基因復制事件有關[20]。系統進化分析顯示,HbGRAS蛋白可分為14 個亞家族,而在水稻的每個亞群中均鑒定到HbGRAS 蛋白,這表明GRAS 家族的分化可能早于單、雙子葉植物的分化。橡膠樹GRAS亞族數量與木薯一致[20],其中LISCL 成員最多(16個,17.58%),與擬南芥和水稻等植物相似[15],表明這些GRAS 基因家族在長期進化過程中可能具有較強的分化能力。基于系統進化關系和多序列比對結果,發現大多數HbGRAS 蛋白的N 端包含1 個高度無序的區域,這導致了GRAS 蛋白的多樣化,并影響了其功能分化。DELLA 亞家族蛋白N 端相對保守。橡膠樹DELLA 亞家族成員中除HbGRAS62 無DELLA 保守結構域外,其余5個均含有DELLA 結構域,這可能是HbGRAS62在進化過程中發生了DELLA 基因的丟失事件,或者該基因不包含典型的“DELLA”和“DXLLX”五肽結構而導致[48]。
分析91個HbGRAS 基因的內含子和外顯子結構發現,無內含子的HbGRAS 基因比例(69.23%)與同科的木薯(53.25%)[20]、蓖麻(78.3%)[22]和麻風樹(95.83%)[21]類似,均較高。內含子可以增加基因長度及重組頻率,有利于物種進化[49]。雖然內含子較少的基因在物種進化或重組中沒有優勢,但它們往往對壓力反應迅速[50]。因此,推測許多橡膠樹GRAS 基因成員可能會對環境變化做出快速響應。
基因擴增是基因組進化的重要驅動力,可以導致新的功能基因的產生和新物種的分化,從而使植物在進化過程中更好地適應環境[17]。染色體定位結果顯示,HbGRAS 基因位于除Chr11外的幾乎所有染色體上,這可能是進化過程中發生片段丟失或染色體移位導致。同時,發現有17 個HbGRAS 基因(18.68%)發生串聯重復事件,所有具有串聯重復序列的HbGRAS 基因均來自同一亞家族,且主要集中在LISCL 亞家族中(58.82%)。這意味著,基因復制保留在全基因組復制后存在一定程度的偏倚,不同亞家族的保留和丟失也不同。另外,有研究發現,如果蛋白質與基因編碼的其他產物存在相互作用,則在復制事件發生后,這類基因會發生偏置[51]。共線性分析顯示,片段重復區域中有87 個HbGRAS 基因(95.6%),說明片段重復在橡膠樹GRAS 基因進化中可能發揮主要作用。
GA促進植物次生細胞壁形成[52],DELLA通過與相關轉錄因子互作調控水稻次生細胞壁中纖維素合成[53]。外施GA3 抑制橡膠樹木質部DELLA表達,DELLA亞家族成員HbGRAS82 和HbGRAS89在木質部中高表達,且隨橡膠樹木質化程度加深上調表達,故推測它們可能參與橡膠樹次生細胞壁的形成。擬南芥DELLA 蛋白競爭性地阻止阻遏蛋白JAZ 與轉錄因子MYC2 結合,從而提高MYC2 對靶基因的調控能力[54];WILD等[55]證實擬南芥RGL3能分別與MYC2 和JAZ 蛋白互作,進而調控JA 介導的響應;MYC2是調控橡膠生物合成途徑之一——JA 信號途徑的關鍵因子。吳紹華等[56]發現割膠和MeJA調控橡膠樹HbGAIPB基因的表達。本研究結果顯示橡膠樹DELLA亞族成員在膠乳中均有表達,且除HbGRAS44外其他成員都含有MeJA 響應元件。據此,推測橡膠樹DELLA基因可能通過調控JA 信號途徑參與橡膠生物合成。以上結果為進一步研究橡膠樹GRAS基因的生物學功能、調控橡膠樹生長發育與逆境響應的分子機制奠定了一定的理論基礎。
