利用酵母雙雜交篩選菠蘿AcSWEET11的互作蛋白
2024-10-09 00:00:00林文秋劉勝輝張秀梅張紅娜李運(yùn)合吳青松
熱帶作物學(xué)報(bào) 2024年9期


關(guān)鍵詞:菠蘿;AcSWEET11;酵母雙雜交;互作蛋白
中圖分類號:S668.3 文獻(xiàn)標(biāo)志碼:A
菠蘿[Ananas comosus(L.)Merr]是世界重要的熱帶果樹之一,也是我國重要的熱帶經(jīng)濟(jì)作物。菠蘿是聚花果植物,花芽分化一旦開始,花序和果實(shí)的發(fā)育就會持續(xù)下去,直到果實(shí)成熟[1],成花與否決定了菠蘿產(chǎn)量與品質(zhì)的形成。目前,生產(chǎn)中菠蘿的開花途徑有2 個,一是經(jīng)過冬季低溫后自然成花[2],二是栽培12~16個月后經(jīng)人工誘導(dǎo)成花。自然開花會導(dǎo)致果實(shí)成熟期不一致,收獲期延長,甚至出現(xiàn)部分植株3~4a才能自然開花的現(xiàn)象,限制了菠蘿產(chǎn)業(yè)的發(fā)展[3]。為了解決這一問題,生產(chǎn)上主要利用乙烯利、電石等乙烯衍生物對菠蘿進(jìn)行人工誘導(dǎo)成花(催花)達(dá)到產(chǎn)期調(diào)節(jié)的效果[4] ,但催花效果受到品種、環(huán)境條件和菠蘿的生長發(fā)育時期的影響。如果催花不當(dāng),會造成減產(chǎn)、果實(shí)品質(zhì)下降,甚至是絕收的情況,嚴(yán)重影響了經(jīng)濟(jì)效益,阻礙了產(chǎn)業(yè)的升級[5] 。因此,解析菠蘿的成花機(jī)制,對促進(jìn)菠蘿產(chǎn)期的調(diào)節(jié)具有重要意義。
SWEET 廣泛存在于植物體,在植物的許多生理過程中發(fā)揮著重要的作用。自2010年CHEN等[6]從擬南芥中第一次鑒定出糖轉(zhuǎn)運(yùn)蛋白基因SWEET 以來,通過對已經(jīng)發(fā)表的基因組序列進(jìn)行全基因組檢索分析,在多種植物中鑒定到了大量的SWEETs[7-9]。SWEET 屬于MtN3/saliva 家族,膜內(nèi)區(qū)域高度保守,具有雙向轉(zhuǎn)運(yùn)糖的功能,包含7 個跨膜a-螺旋結(jié)構(gòu),形成三螺旋束,促進(jìn)糖的跨膜運(yùn)輸[10]。不同植物的SWEET 系統(tǒng)進(jìn)化分析結(jié)果表明,在植物中SWEET 基因家族可以分為4 個亞類,且每個亞類的主要轉(zhuǎn)運(yùn)底物不同:Ⅰ 亞類主要轉(zhuǎn)運(yùn)脫氧葡萄糖, 擬南芥的AtSWEET1 是第一個以糖轉(zhuǎn)運(yùn)為特點(diǎn)的糖轉(zhuǎn)運(yùn)蛋白,具有葡萄糖的轉(zhuǎn)運(yùn)活性,參與花粉對單糖的吸收[11];Ⅱ亞類主要轉(zhuǎn)運(yùn)單糖,葡萄VvSWEET4主要負(fù)責(zé)葡萄糖的轉(zhuǎn)運(yùn),水稻OsSWEET5 參與半乳糖轉(zhuǎn)運(yùn)[11-12];Ⅲ亞類主要轉(zhuǎn)運(yùn)蔗糖,擬南芥AtSWEET11 和AtSWEET12 負(fù)責(zé)將細(xì)胞內(nèi)的蔗糖流出到細(xì)胞壁間隙進(jìn)而裝載到韌皮部用于蔗糖的長距離運(yùn)輸[13];而Ⅳ亞類主要介導(dǎo)了果糖的單向運(yùn)輸,AtSWEET17 定位到液泡膜上,外部供應(yīng)果糖時,其在根延伸區(qū)高度表達(dá),促進(jìn)根從外部吸收果糖,介導(dǎo)液泡對果糖的攝取并儲存[14-15]。SWEETs 調(diào)控植物花器官的發(fā)育,在植物開花過程中發(fā)揮著重要的調(diào)控作用。在水稻中OsSWEET11在圓錐花序和花藥中高表達(dá),遺傳轉(zhuǎn)化研究結(jié)果表明OsSWEET11 調(diào)控小孢子在花粉母細(xì)胞階段和未成熟花粉的發(fā)育。OsSWEET11 通過降低花粉發(fā)育過程中淀粉的含量導(dǎo)致育性下降[7]。OsSWEET14 基因敲除后,突變植株的種子變小,生長延緩,植株的生殖發(fā)育延遲[16]。一些研究結(jié)果表明SWEETs 不僅能夠影響植物的育性,在植物的花器官形成過程中也起著重要的作用。通過分析牡丹花花器官形成過程中葡萄糖含量變化和PsSWEET 表達(dá)量變化發(fā)現(xiàn),PsSWEET8 極可能通過調(diào)控牡丹花瓣葡萄糖的轉(zhuǎn)運(yùn)從而調(diào)控開花過程[17]。最新的研究表明,在長日照條件下超表達(dá)AtSWEET10 后擬南芥提前開花,進(jìn)一步的研究結(jié)果表明AtSWEET10 位于FT 信號途徑的下游[18],這是首次有直接證據(jù)證明SWEET參與了植物成花轉(zhuǎn)變的過程。但是AtSWEET10 是如何調(diào)控開花的機(jī)制尚不清楚。因此,推測AtSWEET10 可能在FT 的誘導(dǎo)下進(jìn)行轉(zhuǎn)錄然后運(yùn)輸糖或者FT 蛋白來促進(jìn)開花[18]。
已有研究表明,菠蘿(無刺卡因)成花過程中可溶性糖和蔗糖含量顯著提高[19],且本課題組前期研究結(jié)果證實(shí),AcSWEET11 能夠促進(jìn)果實(shí)糖積累[20],并能促進(jìn)擬南芥提早開花(待發(fā)表),但其在成花中的作用機(jī)制尚不清楚。本研究利用酵母雙雜交技術(shù), 篩選菠蘿花芽分化過程的cDNA 文庫中與AcSWEET11 互作的候選蛋白,為進(jìn)一步研究AcSWEET11 參與成花調(diào)控的機(jī)制奠定基礎(chǔ)。
1 材料與方法
1.1 材料
酵母粉(yeast extract)和胰蛋白胨(tryptone)購自O(shè)XOID 公司;缺陷培養(yǎng)基(minimal syntheticaldefined medium, SD)DDO(SD/-Leu/-Trp)、TDO( SD/-Leu/-Trp/-His )、TDO/X( SD/-Leu/-Trp/-His/X-a-gal)、QDO(SD/-Leu/-Trp/-His/-Ade)和YPDA 培養(yǎng)基購自北京索萊寶科技有限公司;X-a-gal 購自YEASEN 公司;酵母菌株感受態(tài)NMY51 和pBT3、pNubG-Fe65、pTSU2-APP、pPR3等載體購自武漢金凱瑞生物工程有限公司;菠蘿花芽的cDNA 文庫由本研究中心保存。
1.2 方法
1.2.1 誘餌載體的毒性和自激活檢測 毒性和自激活檢測方法參照王會勤[21]的方法。以pNubGFe65和pTSU2-APP 共轉(zhuǎn)化NMY51 酵母菌為陽性對照,pPR3-N 和pTSU2-APP 共轉(zhuǎn)化酵母菌為陰性對照。將pBT3-STE-AcSWEET11 和獵物空載pPR3-N 通過LiAc 的方法共轉(zhuǎn)化至酵母NMY51中,在DDO、TDO、TDO/AT 和QDO 培養(yǎng)基上30 ℃培養(yǎng)3~4 d,觀察菌落的生長情況,檢測pBT3-STE-AcSWEET11 誘餌蛋白的毒性和自激活活性。通過對pBT3-STE-AcSWEET11 和Post-Nubal 共轉(zhuǎn)化NMY51 酵母菌,在DDO、TDO/3?AT和QDO 培養(yǎng)基上培養(yǎng),觀察菌落的生長情況,檢測pBT3-STE-AcSWEET11 在酵母NMY51 中的表達(dá)功能。
1.2.2 互作蛋白的篩選 從SD/-Leu 平板挑取生長狀態(tài)良好、含有pBT3-STE-AcSWEET11 的單克隆接種至50 mL 的SD/-Leu 液體培養(yǎng)基,30 ℃培養(yǎng)至OD600 為0.2 后,轉(zhuǎn)接至YPDA 液體培養(yǎng)基中,震蕩培養(yǎng),使OD600 為0.6~0.8(12~16 h);室溫離心,收集菌體,用無菌水重懸,離心后棄上清;加入LiAc 重懸菌體,混勻,置冰上,制備含有誘餌載體的NMY51 感受態(tài)。將菠蘿cDNA文庫質(zhì)粒轉(zhuǎn)化含有誘餌載體的NMY51 感受態(tài)后,均勻涂布于TDO/X/3?AT 5 mmol/L 平板上,30 ℃培養(yǎng)3~4 d。挑選TDO/X/3?AT 板上變藍(lán)的單菌落,轉(zhuǎn)接于新的QDO/X 平板上,30 ℃培養(yǎng)3 d,統(tǒng)計(jì)2 種培養(yǎng)基上變藍(lán)的單菌落,即為候選陽性克隆。
1.2.3 候選互作蛋白的測序鑒定 提取陽性克隆質(zhì)粒,利用通用引物(Up: CTTTCCTTATACATTAGGACC,Dn: GGGACCTAGACTTCAGGTTG)進(jìn)行PCR 擴(kuò)增。選取條帶單一,大于500 bp 的有效克隆片段,送至生工生物工程(上海)股份有限公司進(jìn)行測序驗(yàn)證。測序結(jié)果在NCBI(https://www.ncbi.nlm.nih.gov)數(shù)據(jù)庫上進(jìn)行BLAST 比對分析。
1.2.4 生物信息學(xué)分析 利用在線網(wǎng)站UniProt(https://www.uniprot.org)注釋候選蛋白的功能。將獲得的候選蛋白進(jìn)行Gene Ontology(GO)和Kyoto Encyclopedia of Genes and Genomes(KEGG)通路分析。
1.2.5 候選蛋白基因的表達(dá)分析 對乙烯利誘導(dǎo)菠蘿成花過程(0、8、16、32 d)進(jìn)行轉(zhuǎn)錄組測序,獲得花芽分化過程中基因表達(dá)量的數(shù)據(jù),利用HemI 軟件計(jì)算各時期基因表達(dá)量的FPKM(fragments per kilobase of exon model per millionmapped fragments)值,并繪制候選蛋白基因在乙烯利誘導(dǎo)菠蘿成花過程中的表達(dá)熱圖。
2 結(jié)果與分析
2.1 誘餌載體的毒性和自激活檢測
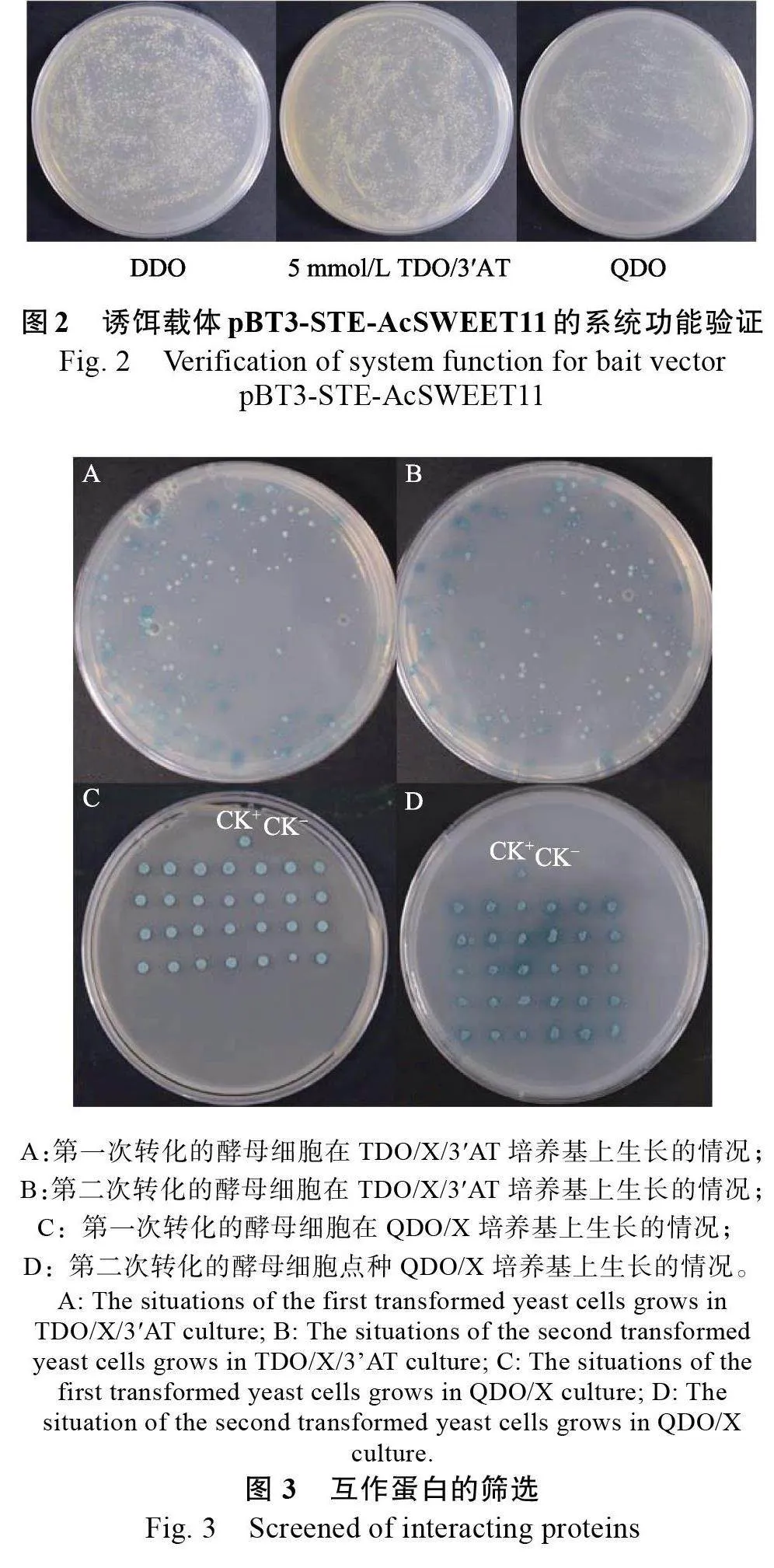
以pNubG-Fe65 和pTSU2-APP 共轉(zhuǎn)化NMY51 酵母菌為陽性對照,pPR3-N 和pTSU2-APP 共轉(zhuǎn)化酵母菌為陰性對照。將pBT3-STEAcSWEET11誘餌重組質(zhì)粒和獵物空載pPR3-N 共轉(zhuǎn)化NMY51 酵母感受態(tài)。結(jié)果表明,pBT3-STEAcSWEET11+pPR3-N 在DDO 培養(yǎng)基上的酵母菌生長狀態(tài)良好,說明pBT3-STE-AcSWEET11 和pPR3-N 已成功轉(zhuǎn)入NMY51 酵母菌中,且對NMY51 酵母細(xì)胞無毒性。pNubG-Fe65+pTSU2-APP 在DDO、TDO/3?AT、QDO 培養(yǎng)基上生長良好,pPR3-N+pTSU2-APP 在DDO 培養(yǎng)基上正常生長,在TDO/3?AT、QDO 無菌落生長,誘餌重組質(zhì)粒pBT3-STE-AcSWEET11+pPR3-N的酵母菌在TDO 培養(yǎng)基能夠生長, 表明pBT3-STE- Ac-SWEET11+pPR3-N 有自激活現(xiàn)象,激活HIS3 的表達(dá)(圖1A~圖1H)。為了抑制酵母菌中HIS3的表達(dá),在TDO 培養(yǎng)基中添加不同濃度的3?AT。結(jié)果顯示,在添加3?AT 后酵母菌沒有生長,HIS3的表達(dá)受到抑制(圖1I~圖1K)。進(jìn)一步分析其在QDO 培養(yǎng)基上的生長情況,結(jié)果顯示,轉(zhuǎn)化pBT3-STE-AcSWEET11+pPR3-N 的酵母菌不能生長(圖1L)。以上結(jié)果表明,pBT3-STE-AcSWEET11+pPR3-N 在TDO/3?AT 培養(yǎng)基和QDO培養(yǎng)基上自激活受到抑制,后續(xù)可在TDO/3?AT 和QDO 培養(yǎng)基上進(jìn)行互作蛋白的篩選。
將Post-Nubal 和pBT3-STE-AcSWEET11 共轉(zhuǎn)化NMY51 酵母菌,分別在DDO、TDO/3?AT 和QDO 培養(yǎng)基上培養(yǎng)。結(jié)果表明,在DDO 培養(yǎng)基上有菌落生長,說明共轉(zhuǎn)化成功;在TDO/3?AT和QDO 培養(yǎng)基上菌落生長良好,表明pBT3-STEAcSWEET11在NMY51 系統(tǒng)能正確表達(dá),可進(jìn)行下一步文庫篩選(圖2)。
2.2 候選互作蛋白的篩選及鑒定
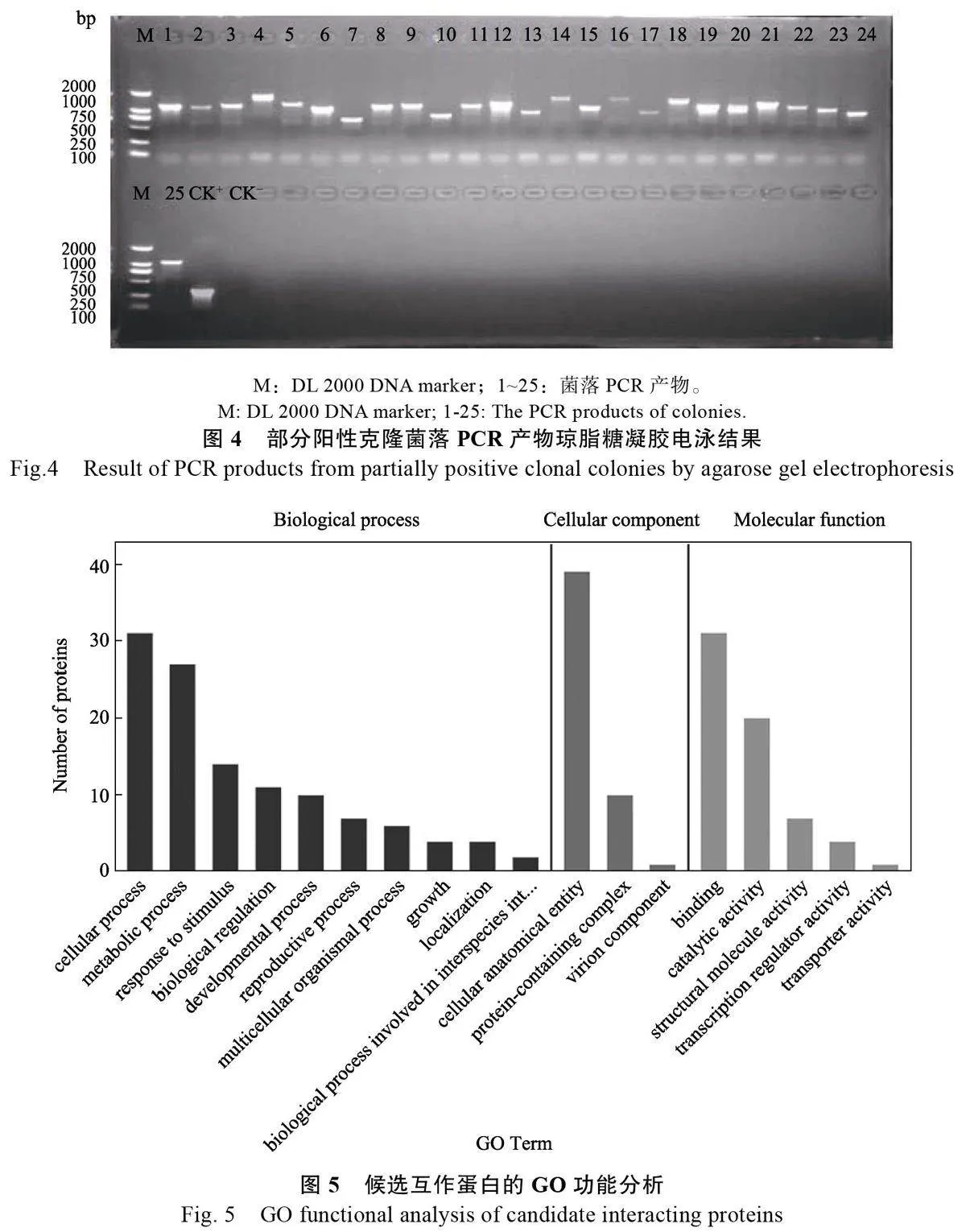
利用菠蘿花芽分化的cDNA 膜文庫篩選與AcSWEET11 互作的蛋白。以pTSU2-APP和pNubGFe65共轉(zhuǎn)化菌液為陽性對照(CK+),pTSU2-APP和pPR3-N 共轉(zhuǎn)化菌液為陰性對照(CK–),經(jīng)過2 次篩選,共獲得81 個在TDO/X/3?AT 培養(yǎng)基中生長良好的藍(lán)色菌落(圖3A、圖3B)。進(jìn)一步加壓培養(yǎng),將58 個大小均一的單克隆點(diǎn)種在含有QDO/X 培養(yǎng)基上進(jìn)行再次篩選。結(jié)果顯示,58個藍(lán)色菌斑均能正常生長(圖3C、圖3D),初步篩選出58 個陽性克隆。將這58 個陽性克隆進(jìn)行菌落PCR 反應(yīng)(pPR3-N 為陽性對照,CK+;H2O為陰性對照,CK–),電泳檢測結(jié)果表明,插入片段條帶單一,大小約為500~2000 bp(圖4)。分別提取陽性克隆質(zhì)粒并進(jìn)行測序分析,合并重復(fù)序列,測序結(jié)果在NCBI 進(jìn)行比對,篩選到48 個可能與AcSWEET11 相互作用的蛋白(表1)。主要包括E3 泛素連接酶RING1-like、海藻糖磷酸合成酶、細(xì)胞色素P450、LUX 轉(zhuǎn)錄因子等。
2.3候選蛋白的生物信息學(xué)分析
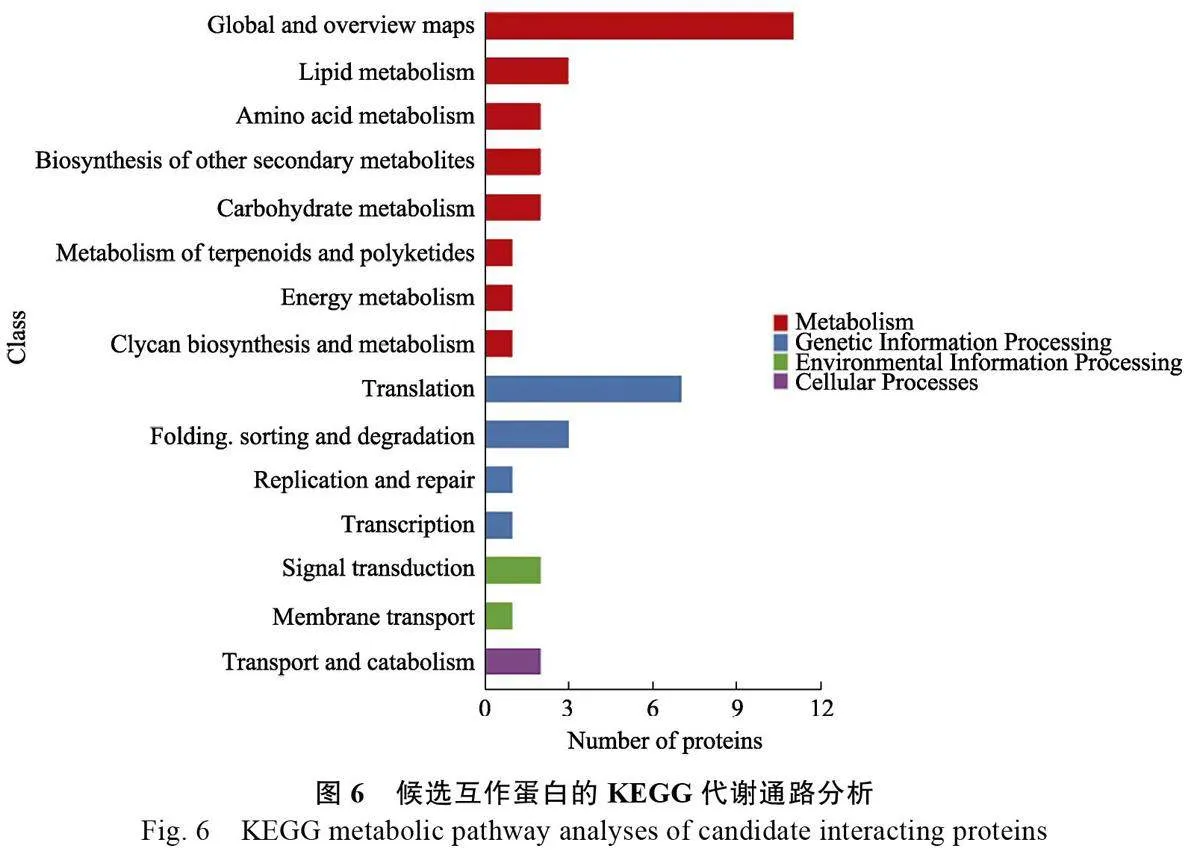
對獲得的48個候選互作蛋白進(jìn)行GO 和KEGG 分類分析。GO分析結(jié)果表明,48個蛋白主要富集在細(xì)胞進(jìn)程(cellular process)、代謝過程(metabolic process)、刺激反應(yīng)(responsetostimulus)、細(xì)胞解剖實(shí)體(cellular anatomicalentity)、結(jié)合(binding)和催化活性(catalyticactivity)等生物過程(圖5)。KEGG 分析結(jié)果顯示,48 個蛋白主要的KEGG 途徑包括脂類代謝(lipid metabolism)、氨基酸代謝(amino acidmetabolism)、其他次生代謝物的生物合成(biosynthesisof other secondary metabolites)和碳水化合物代謝(carbohydrate metabolism)等新陳代謝途徑,翻譯(translation)和折疊、分類和降解(folding,sorting and degradation)等遺傳信息途徑,以及信號轉(zhuǎn)導(dǎo)(signal transduction)與運(yùn)輸和分解代謝(transport and catabolism)等通路(圖6)。
2.4 候選蛋白的基因表達(dá)分析
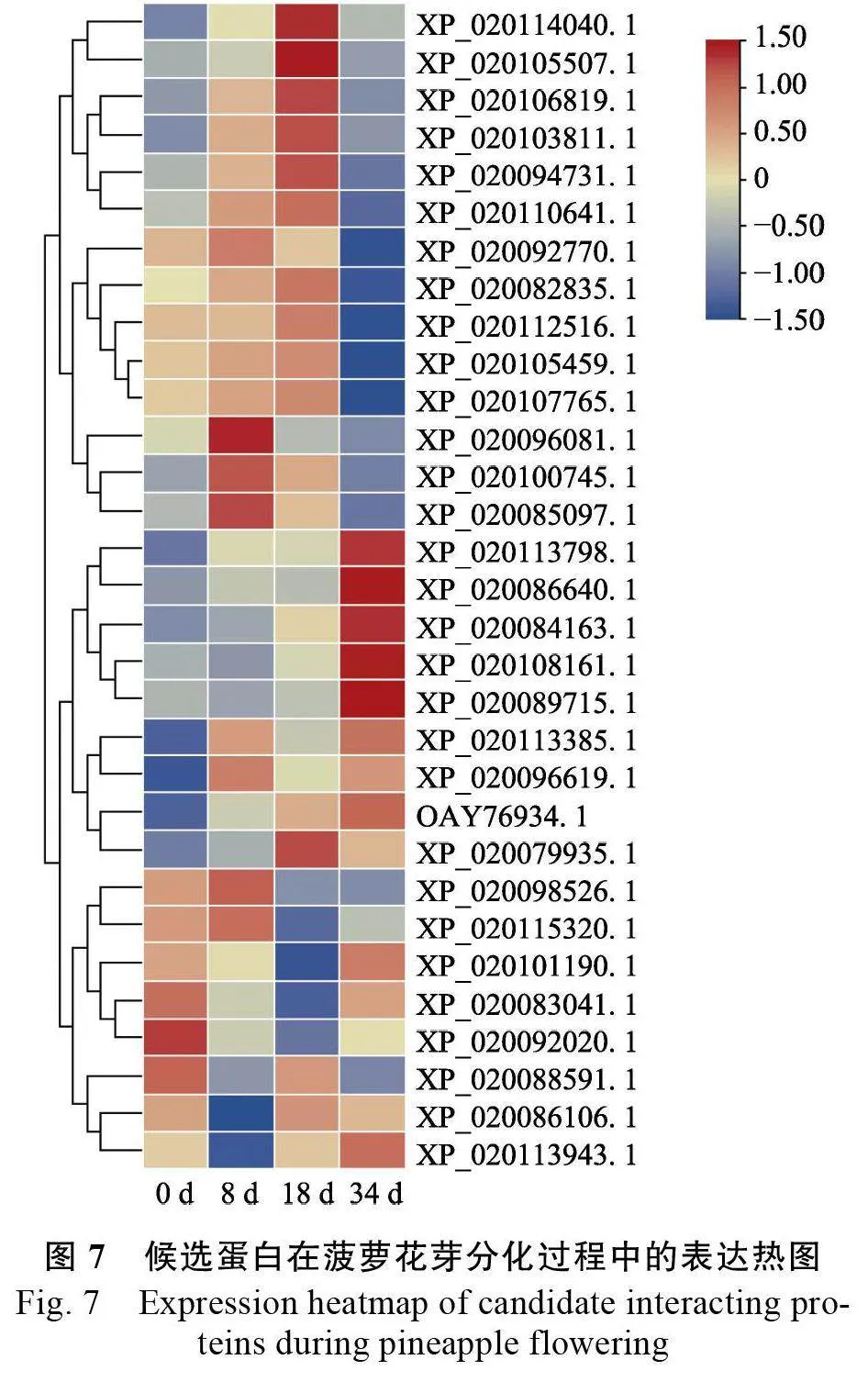
為了確定候選互作蛋白基因的表達(dá)情況,對前期獲得的菠蘿花芽分化過程中的轉(zhuǎn)錄組數(shù)據(jù)進(jìn)行分析,結(jié)果表明,共有30 個候選蛋白基因檢測到表達(dá)量,18個候選蛋白基因在乙烯利誘導(dǎo)菠蘿成花中未檢測到。進(jìn)一步分析結(jié)果表明,Trehalosephosphatesynthase 7(XP_020105459.1)、ProteinTIFY 3-like(XP_020082835.1)、40S ribosomal proteinS27(XP_020092770.1)、Heterogeneous nuclearribonucleoprotein 1-like(XP_020112516.1)等4 個基因與AcSWEET11(XP_020107765.1)表達(dá)一致,在菠蘿成花過程中下調(diào)表達(dá);Dihydrolipoyl dehydrogenase2(XP_020113798.1)、Putative lipidtransferprotein DIR1(XP_020086640.1)、Clathrinassembly protein At4g32285(XP_020108161. 1)等3個基因在菠蘿成花過程中上調(diào)表達(dá)(圖7)。表明Trehalose-phosphate synthase 7 等7 個候選互作蛋白在菠蘿成花過程中具有重要作用。
3 討論
酵母雙雜交是一種篩選互作蛋白的常用方法,廣泛應(yīng)用于未知互作蛋白的篩選和已知蛋白間相互作用的驗(yàn)證。但該系統(tǒng)存在假陽性率和假陰性率高等局限性[22]。自激活和毒性是導(dǎo)致假陽性率和假陰性率高的主要因素。當(dāng)誘餌蛋白和獵物蛋白單獨(dú)或結(jié)合后,出現(xiàn)毒性,抑制酵母細(xì)胞的正常生長,從而出現(xiàn)假陰性;而當(dāng)誘餌蛋白存在自激活活性,獵物蛋白與誘餌蛋白結(jié)合后可激活報(bào)告基因的表達(dá),從而出現(xiàn)假陽性[23]。為了抑制誘餌蛋白的自激活活性,不同缺陷培養(yǎng)基以及在缺陷培養(yǎng)基中添加不同濃度3?AT 被廣泛應(yīng)用于酵母雙雜交的過程中。謝瑞瑩等[23]通過在TDO培養(yǎng)基中添加10 mmol/L 的3?AT,抑制了pGBKT7-NtMYB4a+pGADT7 的自激活活性;在葡萄中,將pGBKT7-VvJAZ9+pGADT7 在QDO/X/A 的培養(yǎng)基上培養(yǎng),其自激活活性受到抑制[24]。本研究結(jié)果顯示,pBT3-STE-AcSWEET11+pPR3-N 存在自激活現(xiàn)象,通過在TDO培養(yǎng)基中添加不同濃度的3?AT 后或在QDO培養(yǎng)基上培養(yǎng),酵母菌不能生長,表明3?AT 和QDO培養(yǎng)基能夠抑制pBT3-STE-AcSWEET11+pPR3-N 的自激活活性。
菠蘿花芽分化是一個復(fù)雜的生物學(xué)過程,受到自身因素和外界環(huán)境條件的影響,涉及糖、氨基酸、植物激素等生理生化的變化[19, 25-26]。本課題組的前期研究結(jié)果表明,AcSWEET11可促進(jìn)可溶性糖的積累[20],但其在成花中的作用機(jī)制尚不清楚。本研究利用酵母雙雜交技術(shù),篩選了48 個與AcSWEET11互作的蛋白,這些蛋白主要富集在氨基酸、碳水化合物以及信號轉(zhuǎn)導(dǎo)和運(yùn)輸、分解等代謝通路上,推測AcSWEET11可能通過氨基酸、碳水化合物和信號轉(zhuǎn)導(dǎo)等途徑參與菠蘿的成花過程。
擬南芥AtSWEET10 能夠促進(jìn)擬南芥提早開花,且處于FT 的下游[18]。在本研究中,沒有篩選到FT 等成花相關(guān)的基因,推測AcSWEET11 可能不是通過直接與FT 等成花相關(guān)基因相互作用參與菠蘿的成花過程。Trehalose-phosphate synthase(TPS)基因在擬南芥成花啟動中具有重要作用,其主要通過調(diào)控SPL 等基因的表達(dá)參與擬南芥的成花過程,是糖作為信號物質(zhì)參與成花的關(guān)鍵途徑[27]。本研究結(jié)果表明,AcSWEET11 和AcTPS7能夠產(chǎn)生互作,且其在菠蘿成花過程中的表達(dá)模式一致,推測AcSWEET11 可能通過TPS 信號途徑參與菠蘿成花。乙烯是誘導(dǎo)菠蘿成花的關(guān)鍵激素,外源乙烯通過促進(jìn)內(nèi)源乙烯的合成誘導(dǎo)菠蘿成花[4]。已有報(bào)道表明E3 泛素連接酶(E3 ubiquitin-protein ligase RING1)能直接與ACS 合成酶(1-aminocyclopropane-1-carboxylate synthase)相互作用來調(diào)節(jié)乙烯的合成[28]。本研究結(jié)果顯示,AcSWEET11 和E3 泛素連接酶存在互作,推測AcSWEET11 可能通過與E3 泛素連接酶互作來調(diào)控乙烯的合成參與菠蘿的成花過程。此外,本研究還篩選到了1個LUX 轉(zhuǎn)錄因子。在水稻中,OsLUX 通過調(diào)控OsELF3-1 和OsELF4s 蛋白抑制Hd1/Ghd7 的表達(dá)參與成花[29]。而在菠蘿中,AcSWEET11是否與水稻相似,能夠通過與LUX相互作用,調(diào)控ELF 等蛋白的表達(dá)來參與菠蘿的成花過程,需要進(jìn)一步研究。
4結(jié)論
本研究通過對pBT3-STE-AcSWEET11+pPR3-N 的毒性和自激活活性的檢測,建立AcSWEET11的酵母雙雜交系統(tǒng)。利用該系統(tǒng)篩選了48個與AcSWEET11互作的候選蛋白。對48 個蛋白進(jìn)行GO和KEGG分析,其主要分布在細(xì)胞進(jìn)程、代謝過程、刺激反應(yīng)和催化活性等生物過程,參與氨基酸代謝、碳水化合物代謝和信號轉(zhuǎn)導(dǎo)和運(yùn)輸?shù)刃玛惔x途徑,推測AcSWEET11 可能通過氨基酸代謝、碳水化合物代謝或者信號轉(zhuǎn)導(dǎo)等途徑參與菠蘿的成花過程。
