基于單拷貝核基因的差不嘎蒿譜系地理與遺傳結構研究
2024-11-05 00:00:00劉夢瑤王鐵娟秀芳唐思如
草地學報 2024年10期



收稿日期:2024-01-26;修回日期:2024-03-24
基金項目:內蒙古自治區自然科學基金項目(2019MS03041);內蒙古師范大學基本科研業務費專項資金(2022 JBTD010)資助
作者簡介:
劉夢瑤(1996-),女,漢族,內蒙古鄂爾多斯人,碩士研究生,主要從事植物生態學研究,Email:1143343811@qq.com;*通信作者Author for correspondence,E-mail:wtj105@163.com
摘要:為揭示我國東北部重要固沙植物差不嘎蒿(Artemisia halodendron)的群體遺傳結構與種群歷史動態,本研究利用2個直系同源單拷貝核基因對來自呼倫貝爾沙地和科爾沁沙地的8個種群154個個體進行分析。結果表明,2個核基因c7847和c9002分別檢測到38和47個單倍型,單倍型多樣性(H)分別為0.9474和0.9158,各種群均在0.8以上,反映出差不嘎蒿有著高的遺傳多樣性。2個基因的分子變異分析(AMOVA) 顯示,差不嘎蒿96.524%和96.985%的遺傳變異來自種群內,種群間和2個沙地間分化很小;種群遺傳分化系數N均大于G,但差異不顯著,表明差不嘎蒿沒有明顯的譜系地理結構。中性檢驗顯示,2個核基因的Tajima’s D值均不顯著,而Fu’s Fs值顯著(PCg4iBgUTcOfvnupoVw6wfQ==<0.05),結合歧點分布分析,該種在中性模式下經歷有限的種群擴張,其中分布區北部的呼倫貝爾沙地的新巴爾虎左旗(DQ)和海拉爾(HLR)種群最為明顯。
關鍵詞:差不嘎蒿;單拷貝核基因;遺傳結構;譜系地理
中圖分類號:Q948.2 文獻標識碼:A 文章編號:1007-0435(2024)10-3034-09
Phylogeography and Genetic Structure of Artemisia halodendron Based on Single Copy Nuclear Genes
LIU Meng-yao, WANG Tie-juan*, XIU Fang, TANG Si-ru
(College of Life Science and Technology, Inner Mongolia Normal University, Key Laboratory of Biodiversity Conservation and Sustainable Utilization in Mongolian Plateau for College and University of Inner Mongolia Autonomous Region, Hohhot, Inner Mongolia 010022, China)
Abstract:To reveal the population genetic structure and historical dynamics of Artemisia halodendron,an important sand-fixing plant in northeast China,phylogeographic analysis was conducted on 154 Artemisia halodendron individuals from eight populations in the Hulunbuir Sandy Land and Horqin Sandy Land using two co-dominant nuclear genes in this study. The results showed that 38 and 47 haplotypes were detected by the two nuclear genes c7847 and c9002,respectively. The haplotype diversity(H)was 0.9474 and 0.9158,respectively,and all populations were above 0.8,which indicated had a high genetic diversity of A. halodendron. Analysis of molecular variance (AMOVA) showed that 96.524% and 96.985% of genetic variation existed within the populations of A. halodendron based on c9002 and c7847,respectively,with limited differentiation between populations and between the two sandy lands. The population genetic differentiation coefficients N were all higher than G,but the difference was not significant (P>0.05),indicating no obvious phylogeographic structure in A. halodendron. The neutral test showed that Tajima’s D values of the two nuclear genes were not significantly different (P>0.05),while Fu’s Fs values had significant differences (P<0.05),and combined with the mismatch distribution analysis,A. halodendron experienced limited population expansion under the neutral mode. Specifically,the New Barag Left Banner (DQ) and Hailar (HLR) populations coming from the Hulunbuir Sandy Land in the northern part of the distribution area,were most pronounced.
Key words:Artemisia halodendron;Single-copy nuclear genes;Genetic structure;Phylogeography
譜系地理學(phylogeography)作為生物地理學的一個重要分支,于1987年由Avise[1]提出。該學科利用分子標記結合種群遺傳學等理論,以及古氣候、孢粉和地質證據等從時間和空間上將物種的遺傳變異形式與地理歷史事件相結合,從而探討物種的進化歷史[2]。在我國,譜系地理學的研究以青藏高原及其毗鄰地區為熱點區域[3],取得了大量的研究成果。對干旱區的研究也有著一定的報道,尤其是第四紀氣候的振蕩、干旱加劇促進了沙漠化的發展,驅動了一些沙生植物的種群分化和替代物種形成[4-7]。
蒿屬(Artemisia L.)的化石孢粉最早見于亞洲北部第三紀漸新世的地層中,在第四紀進行了大規模的遷移與分化,目前廣泛分布于北半球溫帶至亞熱帶,在各類植被的種類組成中均占據著較大的比例[8]。其中龍蒿組(Sect. Dracunculus)植物主要分布于北溫帶的干旱與半干旱地區[9],在草原和荒漠中常見,尤其在我國西北的各大沙漠和北部草原區的沙地中,龍蒿組植物常成為建群種,它們的形成與發展與第四紀冰期的氣候振蕩和沙漠化過程密不可分。
差不嘎蒿(Artemisia halodendron)又名鹽蒿,是蒿屬龍蒿組的半灌木,在我國主要分布于東北部呼倫貝爾沙地和科爾沁沙地[10],在蒙古的東南部和俄羅斯的達烏里地區也有分布,為東蒙古-科爾沁分布種[11]。該種在半流動沙丘、半固定沙地可成為建群種,也可生長于流動沙丘的背風坡、固定沙地和干旱坡地[12]y2uveT2cC4EvM8ED+4k5WXHmHMo1N0hPkRkcRPBwSeI=。差不嘎蒿高50~80 cm,主根粗長,根狀莖發達,具多數營養枝,多由基部分枝,呈密叢,下邊的營養枝常匍匐地面生長,萌生不定根,固著沙土[13]。其種子遇水吸濕后迅速分泌膠質物,粘結沙粒,并保水供種子萌發,成為沙地的先鋒植物,也是我國東北部地區優良的固沙植物[13]。關于差不嘎蒿遺傳多樣性與遺傳分化研究有一定的報道,包括基于隨機擴增DNA多態性(Random amplified polymorphic DNA,RAPD)[14]、簡單重復序列區間(Inter-simple sequence repeats,ISSR)[13,15]的片段分析和核基因內部轉錄間隔區(Internal transcribed spacer,ITS[16])、葉綠體非編碼區(chloroplast non-coding region,psbA-trnH[17])的序列分析,不過ITS與葉綠體基因對差不嘎蒿種群變異的檢測能力不強。作為雙親遺傳的單拷貝核基因核苷酸置換速率更快,在譜系地理學研究中具有重要的價值,尤其對于蒿屬這樣進化的類群,具有更高的靈敏性。本研究基于轉錄組開發得到的直系同源單拷貝核基因,篩選出在差不嘎蒿及其近緣種間存在變異的2個基因片段進行譜系地理學研究,闡明差不嘎蒿的遺傳多樣性、遺傳分化水平以及種群擴張歷史,為龍蒿組植物的遷移進化提供資料,同時也為科爾沁沙地和呼倫貝爾沙地的形成與變遷對物種地理分布格局影響研究提供參考。
1 材料與方法
1.1 試驗材料
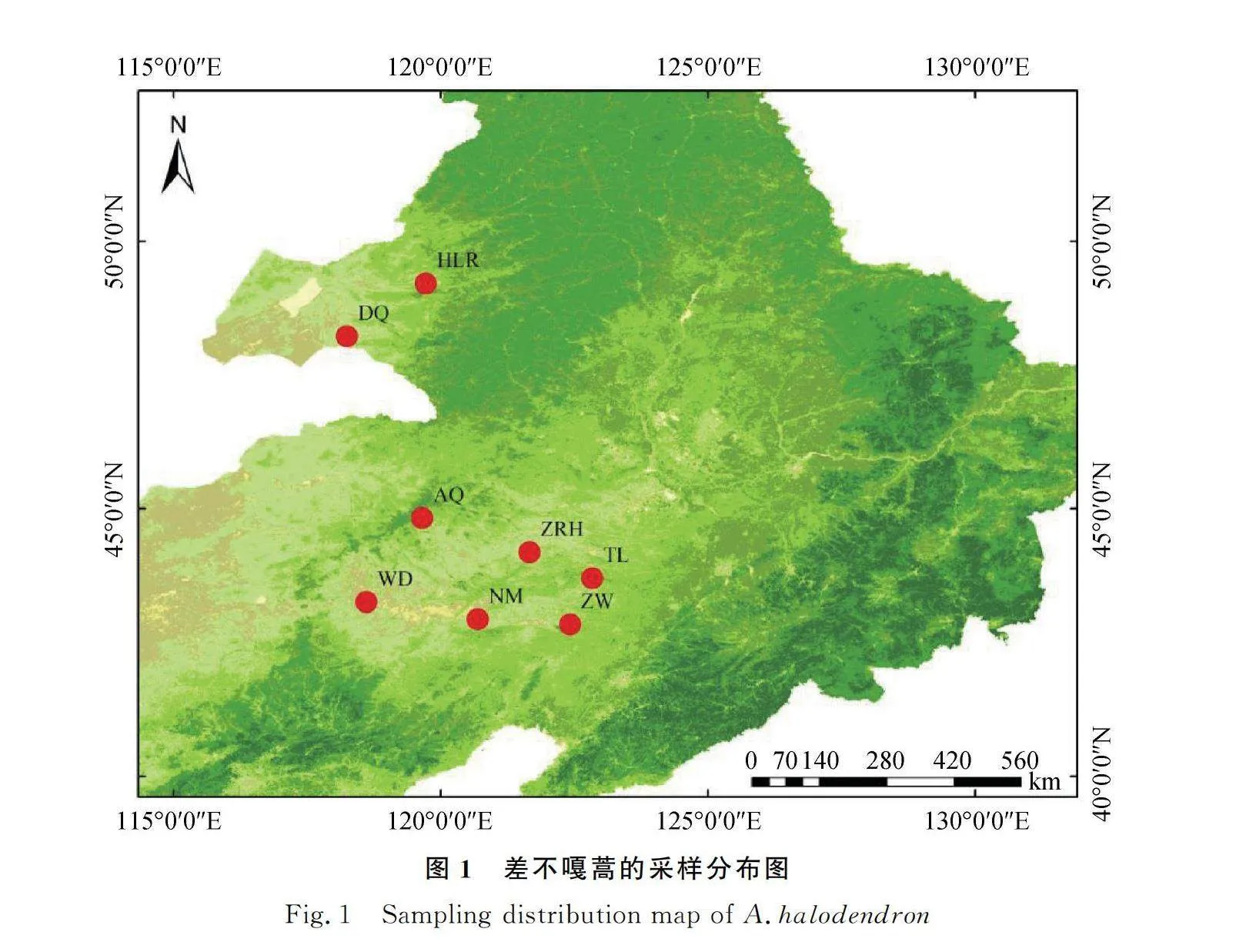
差不嘎蒿分布于我國的呼倫貝爾沙地和科爾沁沙地。呼倫貝爾沙地位于蒙古高原的東北緣,處于大興安嶺西麓森林草原向典型草原的過渡帶,地理坐標為117°10′~121°12′ E,47°20′~49°50′ N,海拔600~800 m,氣候具有半濕潤、半干旱的過渡特點[18],較為寒冷。本研究在呼倫貝爾沙地調查2個種群,調查地點年平均氣溫為-1~-0.7℃,年降水量為281.5~287.4 mm,土壤為風沙土。科爾沁沙地主要分布在西遼河平原,地理坐標為117°49′~123°42′ E,41°41′~46°05′ N,海拔0~2024 m,該地區景觀類型多樣化,流動沙丘、半固定沙丘、固定沙丘等景觀單元交錯分布[19],形成了坨甸相間的地形組合,主體位于半干旱區,東南部臨近半濕潤區。本研究在科爾沁沙地調查6個種群,采樣地點年平均氣溫為3.8~7.5℃,年降水量為356.4~509.2 mm,土壤為風沙土。
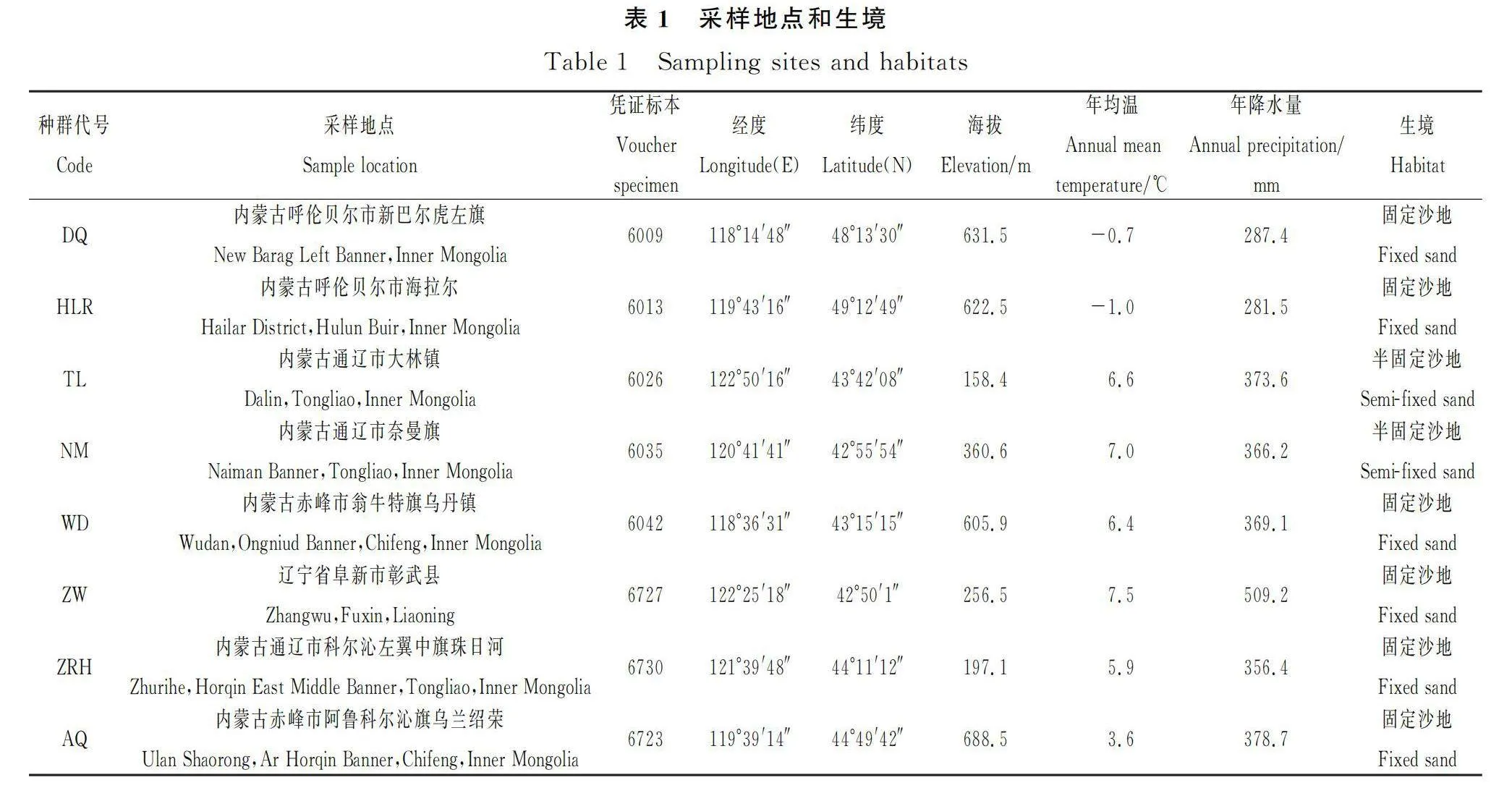
所調查的差不嘎蒿8個自然種群涵蓋該種分布區的中心以及四周,具體地點與生境見表1和圖1。在各種群內隨機選取生長良好的成株24~26株,個體間隔10 m以上距離,采集植株營養枝頂端的嫩葉,每株采集10 g左右,將采集好的樣品放入編號后裝有變色硅膠的塑封袋中,期間更換硅膠使樣品快速干燥。將所有樣品帶回實驗室于-20℃的冰箱保存備用。
1.2 試驗方法
1.2.1 總DNA提取 取每個個體干樣0.3 g,利用北京天根生化科技公司DNA secure的DNA提取試劑盒提取DNA。采用0.7%瓊脂糖凝膠進行電泳檢測,結合超微量紫外分光光度計測量濃度。將DNA母液統一稀釋為40 ng·μL-1,并于-20℃冰箱保存。
1.2.2 引物的篩選 依據前期對內蒙古蒿屬10種龍蒿組植物進行轉錄組測序得到的直系同源單拷貝核基因,選擇在差不嘎蒿、褐沙蒿(A. intramongolica)、黑沙蒿(A.ordosica)(三個種為近緣種)中存在2~5個變異位點的基因序列,共205個。通過NCBI網站中的Primer-Blast進行引物設計,由北京擎科生物公司合成引物。經PCR擴增和測序,選擇擴增效果好、個體間存在較豐富變異的基因c7847,c9002進行研究。c7847引物序列為F:5′-ATGTTCATGCTGACCCCAAT-3′和R:5′-CCGGATCCTCCATCGATAAC-3′,c9002為F:5′-GCCAATGAACGTAGTTTGGATCA-3′ 和R:5′-ACTCA-TTTTGCTGGACACACG-3′。
1.2.3 PCR擴增及測序 將8個差不嘎蒿種群全部個體DNA進行PCR擴增,PCR擴增體系為25 μL:其中12.5 μL Master Mix,2 μL(80 ng)DNA模板,2 μL引物(10 μmol·L-1),8.5 μL dd HO。PCR擴增程序:70℃預變性1 min,94℃變性1 min,55℃退火20 s,72℃延伸50 s,2個循環;接著94℃變性20 s,55℃退火20 s,72℃延伸50 s,38個循環,72℃延伸5 min,4℃保存。擴增后的產物送北京擎科生物公司測序。由于是核基因,測序結果存在雙峰位點,根據雙峰位點組成類型,選擇部分個體進行克隆測序,以期后續對軟件拆分的單倍型序列進行矯正。
1.3 數據處理
通過Chromas軟件(http://technelysium.com.au/)查看每個個體的測序峰圖,將雙峰位點按照IUPAC編碼標準錄入(K=G/T;M=A/C;R=A/G;S=C/G;W=A/T;Y=C/T),利用DNASTAR Lasergene7.1軟件[20]對全部序列進行比對,進而通過MEGA7.0軟件[21]進行手工矯正,前后切平,并保存為Fasta格式。通過DNAsp5.10軟件[22]中的UNPHASE/Genotype功能將核基因序列進行拆分,生成單倍型后,通過克隆測序結果對單倍型進行驗證核對,進而計算單倍型多樣性(H)、核酸多樣性(π)、G,N值以及基因流N。利用Arlequin3.5軟件[23]進行種群分子變異方差分析(AMOVA),計算差不嘎蒿種群的分化程度(1000次置換檢驗)。由PERMUT軟件[24]計算種群遺傳分化系數G和N的值(1000次置換檢驗)。利用Arlequin3.5軟件計算中性檢驗的Tajima’s D,Fu’Fs,SSD,Raggedness index值,通過DNAsp5.10繪制錯配圖。利用軟件POPART[25]中的Median Joining Network模型構建單倍型網絡關系圖。
2 結果與分析
2.1 單倍型多樣性與核苷酸多樣性
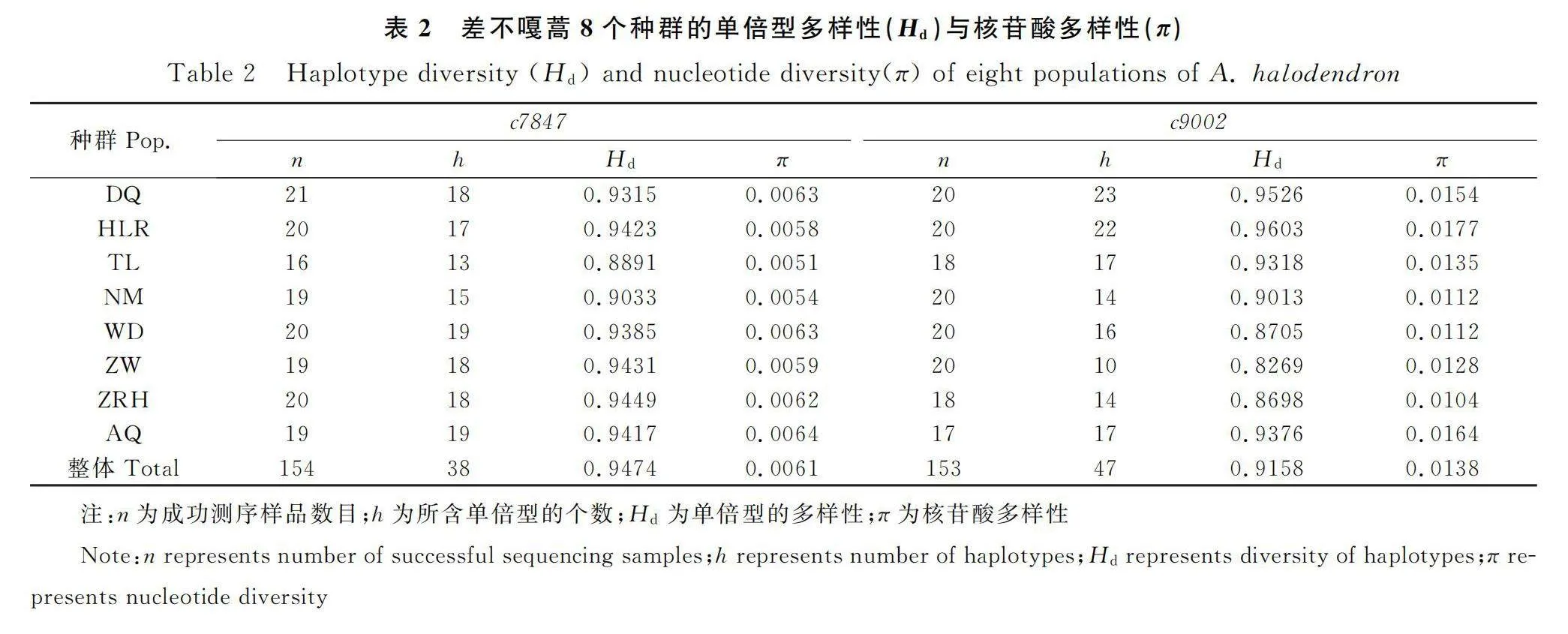
8個差不嘎蒿種群154個個體成功擴增核基因c7847片段,序列長度為475 bp,存在14個變異位點,產生38個單倍型(GeneBank注冊序列號:OR690025~ OR690062);153個個體成功擴增核基因c9002片段,序列長度較c7847短,為378 bp,但變異位點更豐富,有19個,得到47個單倍型(GeneBank注冊序列號:OR690063~ OR690109)。在種的水平上,c7847和c9002的H均很高,分別為0.9474和0.9158,π分別為0.0061和0.0138;在種群水平上,兩個基因的H在0.8891~0.9449和0.8269~0.9603之間,π在0.0051~0.0064和0.0104~0.0177之間。H均大于0.5,π均大于0.005,多樣性程度高[26],且c9002的核苷酸多樣性(π)明顯高(表2)。就各指標綜合來看,呼倫貝爾沙地的2個種群DQ,HLR以及科爾沁沙地的AQ種群多樣性相對較高。
2.2 種群遺傳結構與譜系地理結構分析
基于c7847和c9002的AMOVA分析結果如表3所示,差不嘎蒿種群96.524%和96.985%的遺傳變異來自于種群內,種群間的遺傳變異很小,分別為1.784%和1.836%,只有1.692%和1.179%的遺傳變異存在于2個沙地間(地區間)。F在0~0.05之間說明種群間分化很弱[27],2個基因的F分別為0.035和0.030,表明差不嘎蒿種群間的分化程度處于很弱水平。由基因流的結果可以看出(表4),基于c7847和c9002的基因流N分別為5.320和5.520,均遠遠大于1,為高水平,表明差不嘎蒿種群之間有非常廣泛的基因流動,減少了種群間的遺傳分化。
N與G的大小關系可以顯示種群是否存在譜系地理結構,N>G表明存在譜系地理結構,從差不嘎蒿的結果來看,c7847的G=0.021,N=0.026,雖然N>G,但是差異不顯著;c9002與c7847結果一致,N(0.023)>G(0.012),差異同樣不顯著。結果表明差不嘎蒿種群間沒有明顯的譜系地理結構。
2.3 單倍型網絡關系分析
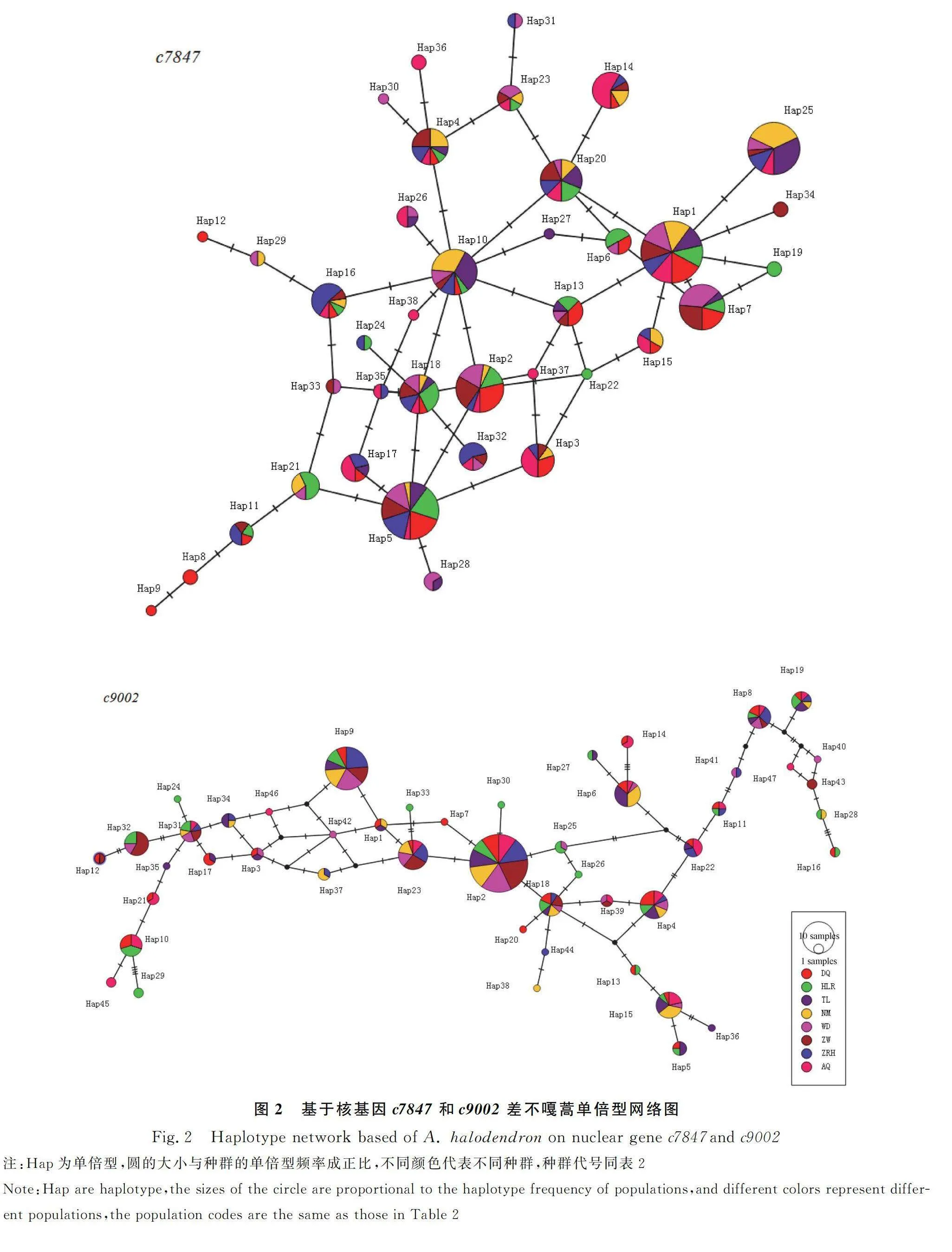
倍型網絡圖能直觀反映單倍型間的關系,圖2中的圓代表單倍型,圓的大小與該單倍型的出現頻率成正比,不同顏色代表不同種群。由c7847得到的網絡圖可知,38個單倍型呈明顯網狀結構,Hap1和Hap5在各種群普遍存在,且數量最多;種群間共享單倍型較多,出現在5~8個種群的單倍型占31.6%,種群特有單倍型有11個,占28.9%。由c9002的結果可知,47個單倍型交錯連接,Hap2處于整個網絡圖的中心,在各種群普遍存在且數量最多,推測為原始單倍型;種群間共享單倍型也較多,有10個單倍型在5~8個種群中出現,占21.3%,種群特有單倍型較多18個,占38.3%。2個基因的種群特有單倍型均為低頻率的,一般只在該種群1~2個個體中出現。
2.4 種群的中性檢驗及種群擴張分析
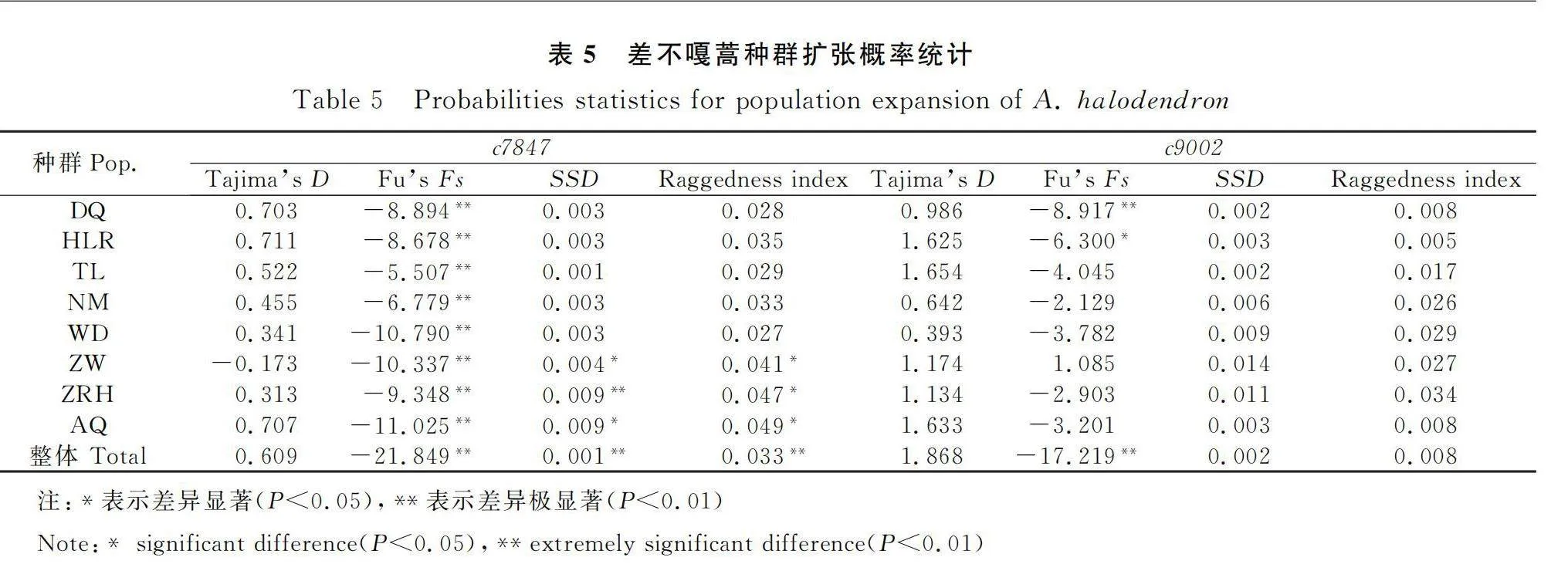
對差不嘎蒿總體和種群進行中性檢驗(表5),2個基因的Tajima’s D值均不顯著(P>0.05),說明符合中性進化假設。c7847和c9002的Fu’s Fs總的值(分別為-21.849和-17.219)均顯著(P<0.01),從錯配分析圖來看(圖3),c7847為單峰、c9002近似單峰,支持Fu’s Fs的結果,表明差不嘎蒿存在一定的種群擴張。以Fu’s Fs結合歧點分布分析,位于分布區北部呼倫貝爾沙地的新巴爾虎左旗(DQ)和海拉爾(HLR)種群在2個基因的各指標顯示存在種群擴張(Fu’s Fs值顯著,SSD和Raggedness index值不顯著),科爾沁沙地的通遼(TL)、奈曼(NM)、烏丹(WD)種群多數指標顯示存在擴張,綜合各指標來看,差不嘎蒿種群在中性模式下經歷了有限的擴張。
3 討論
3.1 差不嘎蒿的遺傳多樣性與遺傳結構
以往對于差不嘎蒿遺傳多樣性的研究,基于片段分析的RAPD[14]和ISSR[13]結果顯示,差不嘎蒿的多態位點百分率在90%以上,基于序列分析的葉綠體psbA-trnH結果顯示,其單倍型多樣性為0.831[17],均表明差不嘎蒿具有高的多樣性水平。本研究采用核基因序列分析,與葉綠體基因相比,2個核基因片段檢測的單倍型多樣性(H)均大于0.9,高于葉綠體基因的結果,2個核基因的核苷酸多樣性(π)分別為0.0061和0.0138,均大于0.005,同樣為高水平,不同于葉綠體的低水平(0.0018)[17]。主要因為核基因的變異明顯高于葉綠體基因,并且本研究選擇差不嘎蒿的不同地理種群,空間的跨度大,能夠檢測出更多的變異性。差不嘎蒿具有高的遺傳多樣性水平,反映出該種作為沙生植物群落的優勢種,對生存環境具有良好的適應性,同時作為蒿屬這一進化類群的成員,高的遺傳多樣性顯示出該種具有強的進化潛能。
從種群遺傳結構來看,基于核基因c7847與c9002的AMOVA分析結果表明,差不嘎蒿的遺傳變異主要來源于種群內,種群間的遺傳分化系數很小,分別占1.784%和1.836%。這與RAPD(12.85%)[14]和ISSR(9.90%[13],10.17%[15])等的研究一致,只是核基因揭示的種群間分化水平更低,與序列分析得到的變異位點不如片段分析豐富有關。差不嘎蒿為風媒傳粉,花粉粒小,花期受東南季風的影響,能夠隨風遠距離傳播,2個基因檢測得到的基因流分別為5.32和5.52,均遠大于1,也說明差不嘎蒿種群間基因交流非常廣泛,減少了種群間的遺傳分化。本研究主要分析不同地理區域間差不嘎蒿自然種群的遺傳變異,因而取樣均選擇該種的典型生境,相似的環境不存在特別的選擇壓力(如人類活動影響、特殊環境等),因而沒有因其他因素造成的遺傳多樣性變化而引起的分化。
3.2 差不嘎蒿的種群歷史動態分析
根據古生物學資料,蒿屬植物被認為起源于第三紀漸新世[28],其起源中心在亞洲北部[9],上新世是該屬植物發育的主要時期,第四紀為該屬植物多樣化和在世界范圍內擴張的重要時期[29]。在第四紀冰期,蒿屬植物在我國境內從東北至西南或西部遷移,差不嘎蒿所在的龍蒿組沿大興安嶺西部至西藏西北部以北的干旱或半干旱地區遷移[8]。差不嘎蒿在我國間斷分布于呼倫貝爾沙地與科爾沁沙地,北部的呼倫貝爾沙地與俄羅斯的達烏里地區和蒙古的東蒙古地區連成一片構成差不嘎蒿的一個分布區域,南部以科爾沁沙地為中心構成一個分布區域,之間為大興安嶺所隔[11]。第四紀歷次冰期和間冰期的旋回,對2個沙地產生巨大的影響,呼倫貝爾沙地自晚更新世以來經歷了8次大的氣候變化[30],科爾沁沙地則形成于第四紀中更新世[31],隨著氣候的變遷也經過了多次的擴張與收縮,2個沙地均具有不同沙層交互成層的沙丘剖面,說明經歷了沙漠化循環發展與逆轉的過程。沙地的形成、發展、活化與固定,必然對植被產生影響,2個沙地不同土層的孢粉組合揭示第四紀以來植被的變化。差不嘎蒿形成的具體時間還沒有古生物學資料的證據,但必然伴隨著第四紀以來氣候的干旱化和沙地的形成而形成,也隨著地質時期氣候的振蕩發生著變化。
在我國,對于沙生植物的譜系地理學研究集中在西北干旱區,如Shi等[5]對荒漠植物霸王(Zygophyllum xanthoxylon)的研究表明西部和東部地區存在多個更新世避難所,并推測霸王最可能起源于東部,經河西走廊向西遷移,種群間的低分化可歸因于物種的擴散和繁殖模式所導致的廣泛的基因流動。白刺(Nitraria tangutorum)的研究也有類似的結果[6],河西走廊可能是白刺冰期時的避難所,并且是該種從東部向西部沙漠地區遷移的“擴散走廊”。Li等[32]對紅砂(Reaumuria soongarica)的研究表明,該種明顯的種群分化和區域范圍擴張可能與青藏高原最近的隆升和末次冰期中國西部沙漠的發展相對應,提出沙漠生境中植物的進化史受到過去地質和氣候變化的嚴重影響。目前,對于我國東北部沙地植物的譜系地理學研究很少,不同于西北地區在第四紀的干旱化和沙漠擴張,我國東北部沙地還受到更新世東亞季風變化的影響,在間冰期和全新世氣候溫暖和濕潤時期,許多流動沙丘被植被覆蓋并固定[30-31],使得沙地不斷經歷沙漠化與綠洲化的交替,這種變化導致沙生植物種群間的遺傳均勻性隨著沙地擴張增加[33]。2個核基因分析顯示差不嘎蒿經歷了一定的種群擴張,其中分布于呼倫貝爾沙地的新巴爾虎左旗(DQ)和海拉爾(HLR)種群擴張最為明顯。呼倫貝爾沙地位于大興安嶺西側,位置靠北,受冰期影響更為明顯,因而種群具有更大的波動。一些西北干旱區植物譜系地理結構會受到山脈隔離作用的影響[4],但霸王的研究卻有所不同,雖跨越了沙漠、山脈等,但顯著屏障的影響并不明顯,表現為缺少譜系地理結構。本研究顯示同樣的特點,差不嘎蒿種群間沒有明顯的譜系地理結構,且種群分化不明顯,除與風媒傳粉有關外,可能也與第四紀氣候變化過程中,經歷了多次擴張與收縮有關,在擴張過程中促成了核基因呈均質化的地理分布模式。
Xu等[33]對科爾沁沙地的東北木蓼(Atraphaxis manshurica)進行研究,結果顯示東北木蓼的總遺傳多樣性較低,但種群間差異較大,避難所通常與高水平的遺傳多樣性相關,其中種群BLY和KQ的遺傳多樣性高,這2個種群所處的地區位于西拉木倫河附近,氣候比其他地區更濕潤,因而認為2個地區是東北木蓼在冰川期持續生存的避難所。差不嘎蒿的研究結果顯示,各種群的遺傳多樣性相差不很大,其中WD種群也位于西拉木倫河附近,但多樣性水平并不高,說明不同物種因繁殖、傳播方式和適應性不同存在著一定的差異。不過差不嘎蒿AQ種群的多樣性水平相對較高,該種群位于科爾沁沙地北緣的阿魯科爾沁旗,地處大興安嶺南麓向西遼河平原過渡的交匯地帶,水分條件更好,且冬季大興安嶺對盛行西北風具有一定的阻擋,自然條件較好,因而推測可能是差不嘎蒿避難所之一,尚有待于其他方法的進一步研究與證實。
4 結論
本研究首次利用直系同源單拷貝核基因對差不嘎蒿8個自然種群進行譜系地理學研究,結果顯示差不嘎蒿具有高的遺傳多樣性,種群間遺傳分化很小,存在廣泛的基因交流。差不嘎蒿的單倍型網絡圖交錯連接,種群共享單倍型較多,沒有明顯的譜系地理結構。中性檢驗與歧點分布分析顯示,該種在中性模式下經歷了有限的種群擴張,其中分布區北部呼倫貝爾沙地的新巴爾虎左旗(DQ)和海拉爾(HLR)種群最為明顯,科爾沁沙地的通遼(TL)、奈曼(NM)、烏丹(WD)種群也存在一定的擴張。
參考文獻
[1]AVISE J C,ARNOLD J,BALL R M,et al. Intraspecific phylogeography:the mitochondrial DNA bridge between population genetics and systematics [J]. Annual Review of Ecology and Systematics,1987,18(1):489-522
[2]AVISE J C. Phylogeography:the history and formation of species[M]. Cambridge:Harvard University Press,2000:3-9
[3]QIU Y X,FU C X,COMES H P. Plant molecular phylogeography in China and adjacent regions:Tracing the genetic imprints of Quaternary climate and environmental change in the world’s most diverse temperate flora [J]. Molecular Phylogenetics and Evolution,2011,59 (1):225-244
[4]MENG H H,GAO X Y,HUANG J F,et al. Plant phylogeography in arid Northwest China:Retrospectives and perspectives[J]. Journal of Systematics and Evolution,2015,53(1):33-46
[5]SHI X J,ZHANG M L. Phylogeographical structure inferred from cpDNA sequence variation of Zygophyllum xanthoxylon across north-west China[J]. Journal of Plant Research,2015,128(2):269-282
[6]YIN H X,WANG L R,SHI Y,et al. The East Asian Winter Monsoon acts as a major selective factor in the intraspecific differentiation of drought-tolerant Nitraria tangutorum in Northwest China [J]. Plants,2020,9(9):1100
[7]唐思如,王鐵娟,秀芳,等. 基于單拷貝核基因的褐沙蒿遺傳結構和譜系地理學分析[J]. 植物研究,2022,42(6):956-963
[8]林有潤. 論世界蒿屬植物區系[J]. 植物研究,1995,15(1):1-37
[9]林有潤. 論蒿屬的演化系統兼論蒿屬與鄰近屬的親緣關系[J]. 植物研究,1982,2(2):1-60
[10]劉夢瑤,李偉瓊,王鐵娟. 差不嘎蒿莖葉解剖特征及其與氣候因子的關系[J]. 草地學報,2021,29(5):1118-1124
[11]王鐵娟,楊持,呂桂芬,等. 中國北部六種沙蒿的地理替代規律及其主導生態因子[J]. 生態學報,2005,25(5):1012-1018
[12]李進. 差不嘎蒿的分布及其在天然植被演替中的地位[J]. 中國沙漠,1991,11(2):55-60
[13]王鐵娟,李偉瓊,張淑艷,等. 差不嘎蒿自然種群的遺傳多樣性與遺傳分化[J]. 林業科學,2010,46(12):171-175
[14]王鐵娟,韓國棟. 中國蒿屬沙生地理替代種親緣關系的RAPD分析[J]. 植物研究,2007,27(4):449-454
[15]HUANG W D,ZHAO X Y,ZHAO X,et al. Relationship between the genetic diversity of Artemisia halodendron and climatic factors[J]. Acta Oecologica,2014,55:97-103
[16]黃文達,趙學勇,左小安,等. 不同生境梯度下差不嘎蒿(Artemisia halodendron)種群ITS基因的序列分析[J]. 中國沙漠,2013,33(2):554-559
[17]HUANG W D,ZHAO X Y,ZHAO X,et al. Genetic diversity in Artemisia halodendron (Asteraceae) based on chloroplast DNA psbA-trnH region from different hydrothermal conditions in Horqin sandy land,northern China[J]. Plant Systematics and Evolution,2013,299(1):107-113
[18]張天琪,楊光,劉峰,等. 呼倫貝爾沙地2000-2020年土地利用變化及生態服務價值[J]. 水土保持通報,2021,41(4):331-338,349
[19]陳銀萍,李曉輝,羅永清,等. 科爾沁不同沙地類型植被動態特征及其與凋落物的關系研究[J]. 草地學報,2021,29(1):115-120
[20]MISENER S,KRAWETZ S A. Methods in molecular biology[M].Totowa:Humana Press,2000:71-91
[21]KUMAR S,STECHER G,TAMURA K. MEGA7:Molecular evolutionary genetics analysis version 7.0 for bigger datasets[J]. Molecular Biology and Evolution,2016,33(7):1870-1874
[22]LIBRADO P,ROZAS J. DnaSP v5:A software for comprehensive analysis of DNA polymorphism data[J]. Bioinformatics,2009,25(11):1451-1452
[23]EXCOFFIER L,LISCHER H. Arlequin suite ver 3.5:a new series of programs to perform population genetics analyses under Linux and Windows[J]. Molecular Ecology Resources,2010,10(3):564-567
[24]PONS O,PETIT R J. Measuring and testing genetic differentiation with ordered versus unordered alleles[J]. Genetics,1996,144(3):1237-1245
[25]LEIGH J W,BRYANT D. POPART:full-feature software for haplotype network construction[J]. Methods in Ecology and Evolution,2015,6(9):1110-1116
[26]GRANT W S,BOWEN B W. Shallow population histories in deep evolutionary lineages of marine fishes:Insights from sardines and anchovies and lessons for conservation [J]. Journal of Heredity,1998,89(5):415-426
[27]WRIGHT S. The interpretation of population structure by F-statistics with special regard to systems of mating[J].Evolution,1965,19(3):395-420
[28]LIU Z F,ZHAO X X,WANG C S,et al. Magnetostratigraphy of Tertiary sediments from the Hoh Xil Basin:implications for the Cenozoic tectonic history of the Tibetan Plateau [J]. Geophysical Journal International,2003,154(2):233-252
[29]WANG W M. On the origin and development of Artemisia (Asteraceae) in the geological past [J]. Botanical Journal of the Linnean Society,2004,145(3):331-336
[30]楊湘奎,杜紹敏,張烽龍. 呼倫貝爾高原晚更新世以來的古氣候演變[J]. 自然災害學報,2006,15(2):157-159
[31]裘善文. 試論科爾沁沙地的形成與演變[J]. 地理科學,1989,9(4):317-328
[32]LI Z H,CHEN J,ZHAO G F,et al. Response of a desert shrub to past geological and climatic change:A phylogeographic study of Reaumuria soongarica (Tamaricaceae) in western China[J]. Journal of Systematics and Evolution,2012,50 (4):351-361
[33]XU Z,ZHANG M L. The effect of past climatic oscillations on spatial genetic structure of Atraphaxis manshurica (Polygonoideae) in the Horqin sand lands,northern China[J]. Biochemical Systematics and Ecology,2015,60:88-94
(責任編輯 閔芝智)
