高州油茶與小果油茶種仁蛋白質組差異比較
2024-11-07 00:00:00魏尚霖李晨蕾廖柏勇程俊森代文魁姜維王溢李永泉
經濟林研究 2024年3期
關鍵詞:高州油茶;小果油茶;種仁;蛋白質組;非標記技術
中圖分類號:S601;S794.4 文獻標志碼:A 文章編號:1003—8981(2024)03—0010—15
油茶屬于山茶科Theaceae 山茶屬Camellia 小喬木或灌木,是我國南方地區重要的木本油料樹種[1-2]。油茶種質資源豐富,小果油茶Camelliameiocarpa 和高州油茶Camellia drupifera 是其中具有代表性的2 種。小果油茶的特點是適應性較強,抗炭疽病能力較強,產量穩定,單果出籽率和含油率優于普通油茶,栽培面積和年產量僅次于普通油茶,主要分布在江西、福建、湖南、廣西、貴州和廣東北部等地區[3]。高州油茶,又名華南油茶、大果油茶,具有果實大且樹體較高的特點,是培育果大皮薄、高產優質油茶的重要種質資源,主要分布在廣東、廣西、海南和香港等地,栽植面積和產量均排在全國第3 位[4]。
油茶具有廣泛的應用前景,不但可用于園林觀賞,而且非產油部位的果皮可用于堆漚作為肥料,果皮提取物質還可用于制作護膚產品和水果保鮮等,種仁是油茶產油的主要部位,其中富含脂質、蛋白質、淀粉、黃酮類、酚酸類、有機酸類、萜類、木脂素類等物質[5-6]。從油茶種仁中提取的油脂含有豐富的油酸、亞油酸、亞麻酸等不飽和脂肪酸及棕櫚酸、硬脂酸等飽和脂肪酸,還有生育酚、角鯊烯、多酚類化合物等主要成分,以及胡蘿卜素、維生素、礦物質、皂苷、三萜醇、鞣質和芝麻素等物質[7-11]。小果油茶油脂中包含約80% 的油酸,含有較多的棕櫚酸和亞油酸,少量硬脂酸、亞麻酸、棕櫚油酸和花生烯酸等,還含有多酚、角鯊烯、植物甾醇和維生素E 等生物活性物質,營養價值極高[12-15]。高州油茶油脂中,油酸含量同樣最高,其次為棕櫚酸、亞油酸、硬脂酸和亞麻酸,不飽和脂肪酸含量可達90%。高州油茶的油脂中,具有與小果油茶類似的活性物質,還富含黃酮類和酚酸類物質。研究結果表明,高州油茶油脂具有普通油茶油脂約3 倍的角鯊烯含量和更高的β- 谷甾醇含量,說明高州油茶種仁在抗氧化活性方面具有極高價值[6,16-18]。
油茶是以產油為主要種植目的的經濟林樹種,茶油的產量和品質決定了其價值。茶油產量受種仁大小、發育進程以及油脂積累過程等諸多因素影響,茶油品質則與抗氧化活性、脂肪酸組成及其生物合成水平等緊密相關。趙松子等[19]對普通油茶轉錄組的數據進行了分析,用油茶SPATULA/ALCATRAZ 基因的mRNA 序列搜索,發現了CoALC、CoSPT1、CoSPT2 等與油茶果實發育、大小相關的基因;趙松子[20] 通過對普通油茶CYP78A10同源基因的mRNA 序列搜索,發現CoCYP78A10a、CoCYP78A10b、CYP78A10、GmCYP78A10 等基因可能參與種子大小的調控;Ji等[21] 采用加權基因共表達網絡分析法研究了普通油茶種子大小相關基因,篩選到21 個樞紐基因,其中SPL4、ABI4、YAB1、KLU 等基因在不同時期的表達水平發生顯著改變,參與調控種子大小的過程;吳波等[22] 研究了油脂合成源基因(GPD1)和匯基因(DGAT1 和DGAT2)在高產油量與低產油量的油茶種子中的表達差異及其對油脂合成積累的影響,發現GPD1 基因為油脂合成積累了更多初始底物,從而促進種子油脂合成,DGAT1 和DGAT2基因在種子發育期間的高表達促進了油脂積累;Wu 等[23] 對高產和低產普通油茶進行轉錄組測序和qRT-PCR 分析,發現上游基因HAD、EAR 和KASI的協同高表達為油酸生物合成提供了前體資源,SAD 基因的持續高表達加速了油酸的合成和積累,而下游基因FAD2、FAD3 等的協同低表達降低了油酸轉化的消耗;郭鈺柬[24] 利用qRT-PCR 技術對越南油茶的4 個關鍵酶基因進行測定,發現了SAD、FAD2、FAD3 等基因參與脂肪酸合成的作用機制。這些研究結果揭示了影響油茶種仁大小、發育過程、油脂合成和油脂積累水平的相關基因及其表達情況,但仍未能完整地反映調控種仁性狀的作用機制。蛋白質能直接或間接地控制生理性狀的表達,通過比較2 種油茶種仁的蛋白質組差異,可以初步探索調控油茶油脂合成和積累的蛋白質組學機制。基于此,本研究中采用非標記(label-free)蛋白質組學技術,對小果油茶和高州油茶種仁的蛋白質組進行比較分析,挖掘2 種油茶種仁中與種仁大小、發育、油脂合成積累及其成分等相關的關鍵蛋白,分析差異蛋白的生物學功能,以期為油茶分子育種提供理論參考。
1 材料與方法
1.1 供試材料
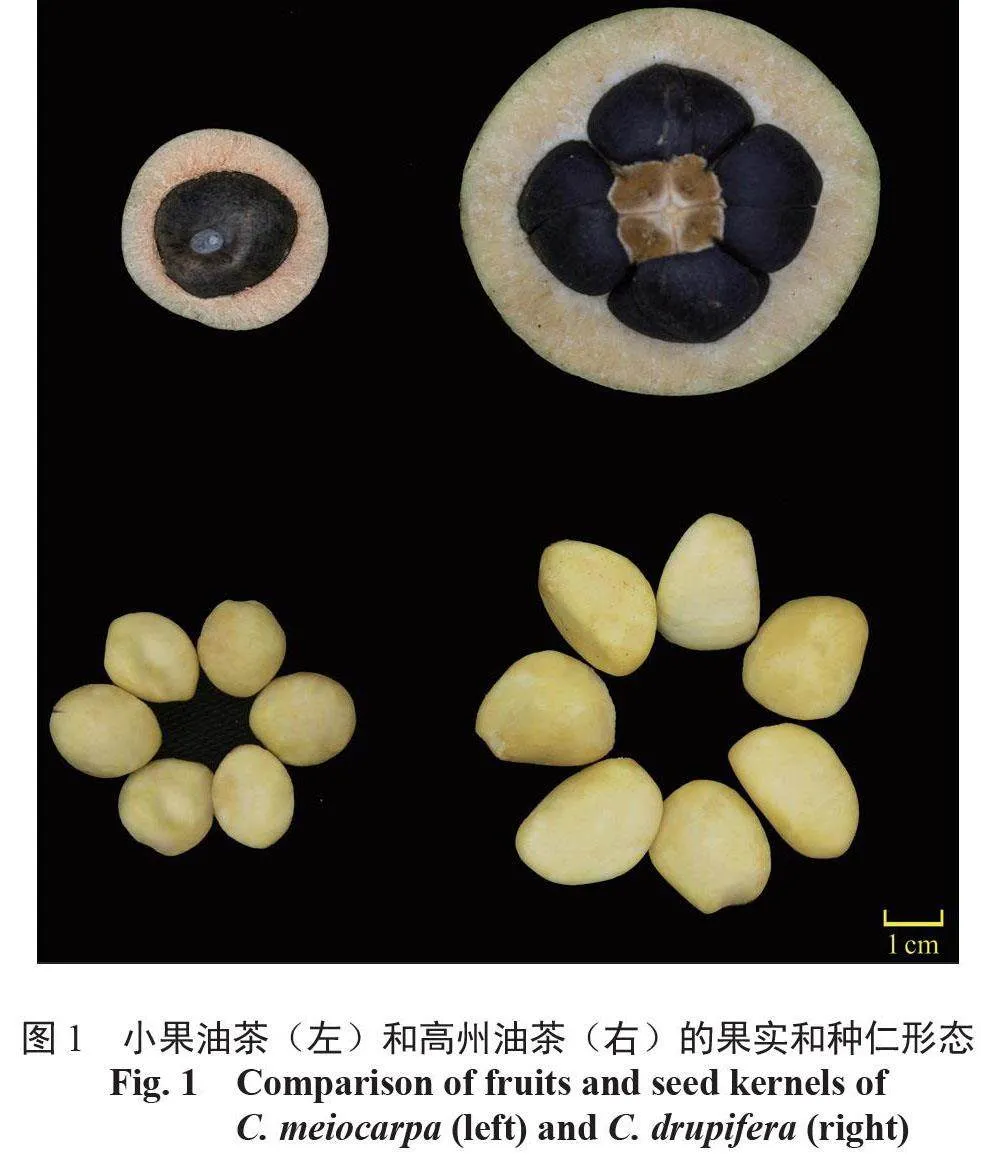
用于試驗的高州油茶和小果油茶的成熟果實及其種仁如圖1 所示,高州油茶于2020 年10 月中旬(成熟期)采自中國廣東省惠州市博羅林場(114°34′E,23°23′N), 小果油茶于2020 年11月中旬(成熟期)采自中國廣東省韶關市曲江小坑林場(113°35′E,24°15′N)。2 個品種均隨機選擇3 棵盛果期單株,采集樹冠中上部成熟果實,取其種仁放入凍存管,高州油茶種仁樣品標記為GZ255-2-1、GZ255-2-2、GZ255-2-3,小果油茶種仁樣品標記為YPZZ13-1、YPZZ13-2、YPZZ13-3,隨后置于液氮中快速冷凍,保存于-80 ℃超低溫冰箱中備用。
1.2 試驗方法
1.2.1 蛋白質提取和酶解
取1g種仁樣品置于預冷的研缽中,加入液氮充分研磨至粉末狀。參照文獻[25-26] 中的方法,進行蛋白質的提取。蛋白質濃度參照Bradford 法[27]測定。隨后,對蛋白質進行酶解,樣品的消化肽段在SPE Cartridges C18(標準密度)濾芯(Sigma-Aldrich公司)上進行脫鹽,管床內徑7 mm,體積3 mL。真空離心濃縮后用40 μL 體積比0.1% 的甲酸重構。
1.2.2 液相色譜串聯質譜(LC-MS/MS)分析
LC-MS/MS 分析在Q-Exactive 質譜儀上進行,該質譜儀與Easy nLC 聯用。根據定量后的酶解產物濃度,從各組酶解的樣品中分別取2 μg 上樣分析。
1.2.3 蛋白質鑒定和定量
使用MaxQuant(1.5.3.17)軟件對每個樣品的LC-MS/MS 原始數據進行數據搜索,對各組的原始數據進行鑒定和定量分析。使用公共蛋白數據庫UniProt(https://www.uniprot.org/)對蛋白質進行定性分析。
1.2.4 差異表達蛋白質篩選
使用公共蛋白數據庫UniProt 篩選出差異表達的蛋白質,包括顯著差異表達蛋白質和部分差異表達蛋白質。顯著差異表達蛋白質,是指組間比較中差異倍數(fold change,FC)大于2.0 或小于0.5 的差異蛋白質;部分差異表達蛋白質,指組間比較中一組樣品具有2 個及以上的非空值,而另一組樣品具有空值的差異蛋白質。
1.2.5 生物信息學分析
分別使用CELLO(http://cello.life.nctu.edu.tw/)軟件和在線網站NCBI Conserved Domain(https://www.ncbi.nlm.nih.gov/cdd/)的CDD 數據庫進行亞細胞定位分析和蛋白結構域分析。
小果油茶和高州油茶種仁中差異表達的蛋白質使用Cluster 3.0 軟件采用層次聚類法進行聚類,并將結果顯示在熱圖中。
采用GO 數據庫(http://www.geneo-ntology.org/) 和KEGG 通路數據庫(http://www.genome.ad.jp/kegg/)對差異蛋白進行GO 功能注釋和所參與代謝通路分析,獲得差異蛋白質的生物功能、所參與生物過程以及細胞定位等信息。
通過Fisher 精確檢驗進行富集分析,應用多重測試的Benjamini- Hochberg 校正來調整得到的P 值,P 值小于0.05 的功能類別和通路被認為是顯著的。
使用String 數據庫(https://string-db.org/)進行蛋白質互作網絡(protein-protein interaction networks,PPI)分析,通過Cytoscape軟件(http://www.cytoscape.org/)進行可視化和編輯。
2 結果與分析
2.1 種仁蛋白質的識別與鑒定
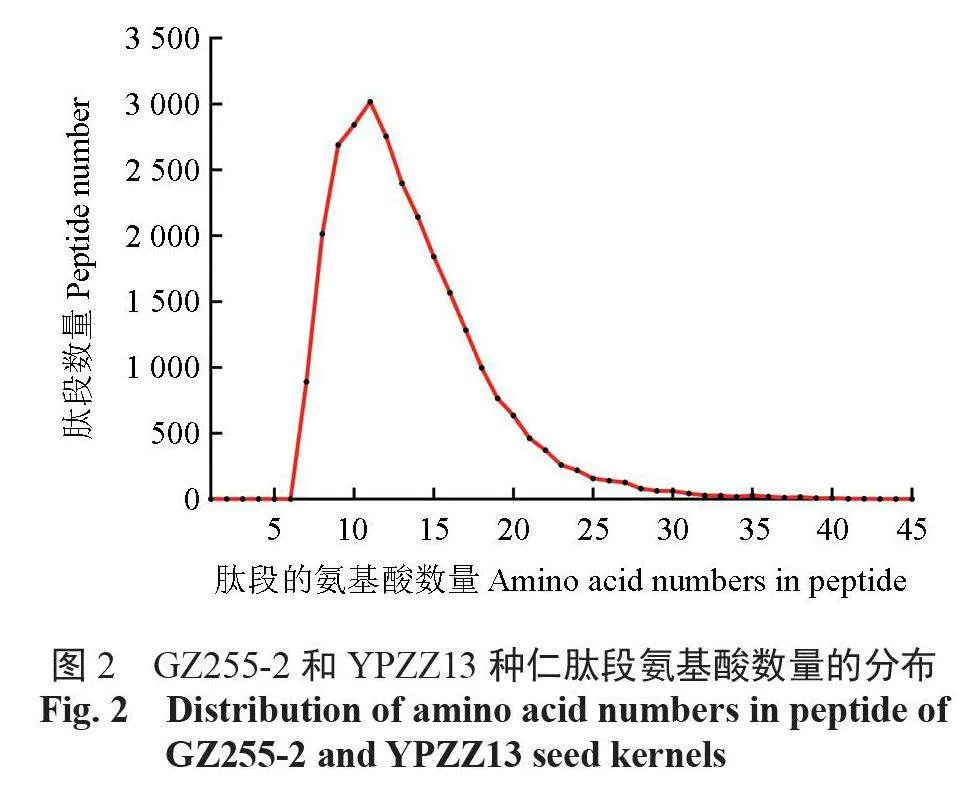
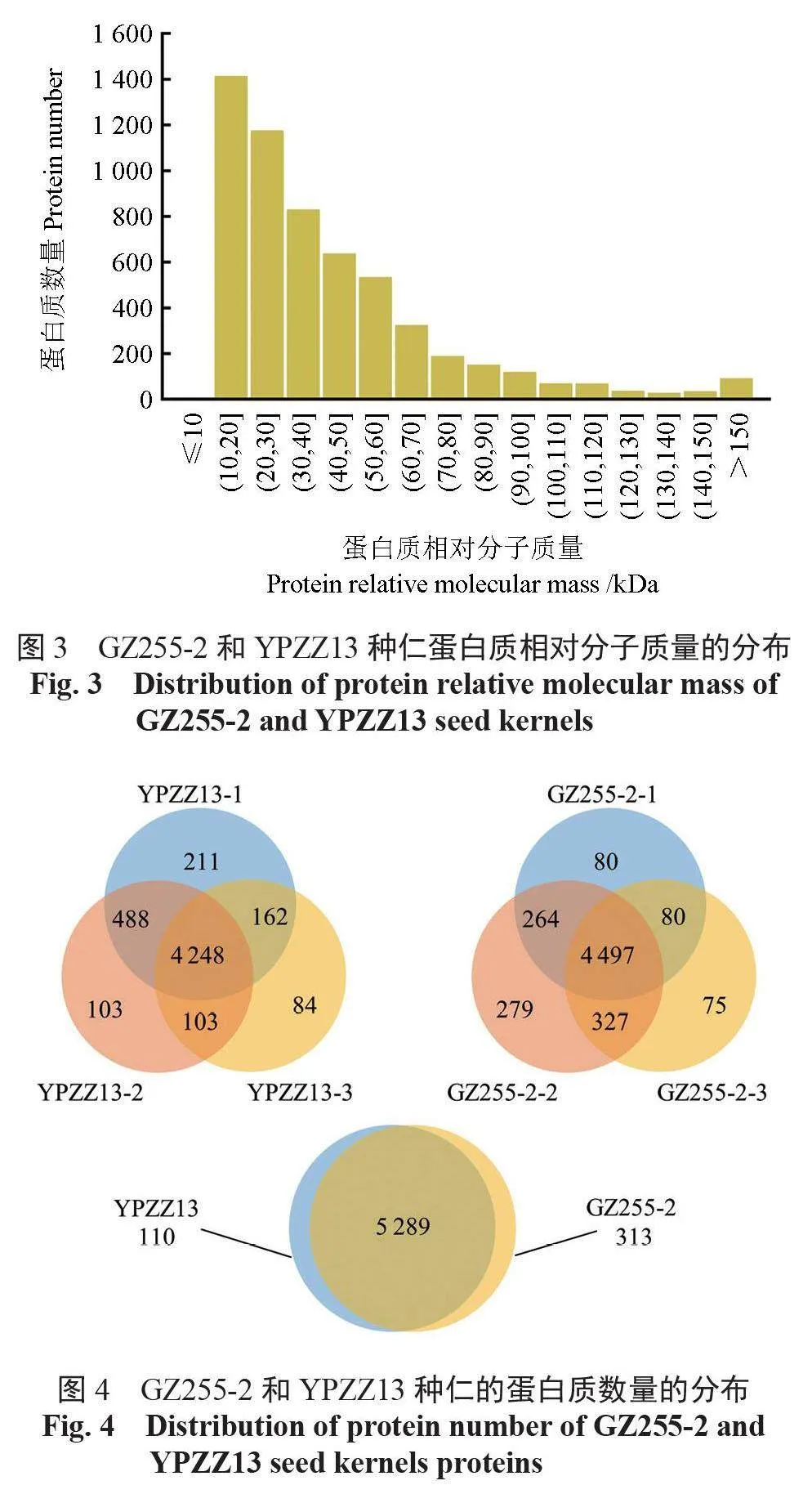
酶解后總蛋白的質譜鑒定結果顯示,共有27 983 個肽段被鑒定,肽段的氨基酸組成數量集中分布在7~22,占總數量的95.31%(圖2)。所有肽段對應5712個蛋白質,其相對分子質量主要為10 ~ 100 kDa,占總數量的94.15%(圖3)。
如圖4所示, 在YPZZ13中共鑒定出5399個蛋白質, 其中有4248個共有的蛋白質,在GZ255-2中共鑒定出5602個蛋白質, 其中有4497個共有的蛋白質。通過組間比較發現,GZ255-2和YPZZ13中共鑒定出5712個蛋白質,其中有5289個共有的蛋白質,差異蛋白質有110個屬于YPZZ13,313個屬于GZ255-2。
2.2 種仁蛋白質的亞細胞定位及結構域注釋
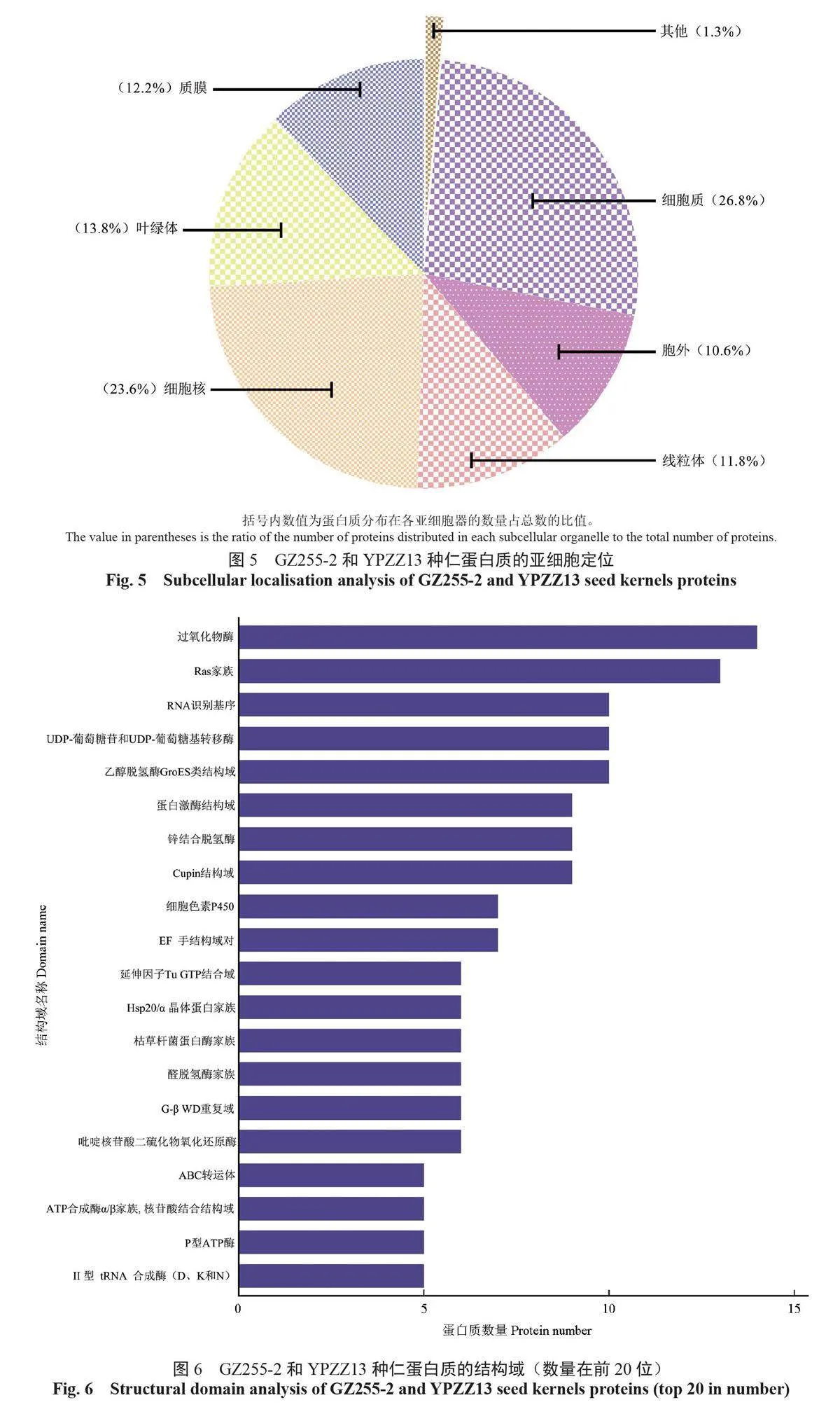
蛋白質的亞細胞定位可揭示其在細胞中發揮的功能,GZ255-2和YPZZ13中的蛋白質分布在細胞內的大部分亞細胞器中,體現功能的主要亞細胞器依次分別是細胞質、細胞核、葉綠體、質膜、線粒體和胞外,在溶酶體、過氧化物酶體、高爾基體、細胞骨架等其他亞細胞器中體現了部分功能(圖5)。
結構域是蛋白質中能獨立折疊和發揮功能的基本單位,GZ255-2和YPZZ13的蛋白質結構域分析結果顯示,蛋白質數量排名靠前的結構域主要為過氧化物酶、Ras家族、RNA 識別基序、UDP-葡萄糖苷和UDP-葡萄糖基轉移酶、乙醇脫氫酶GroES類結構域等(圖6)。
2.3 種仁差異表達蛋白的篩選
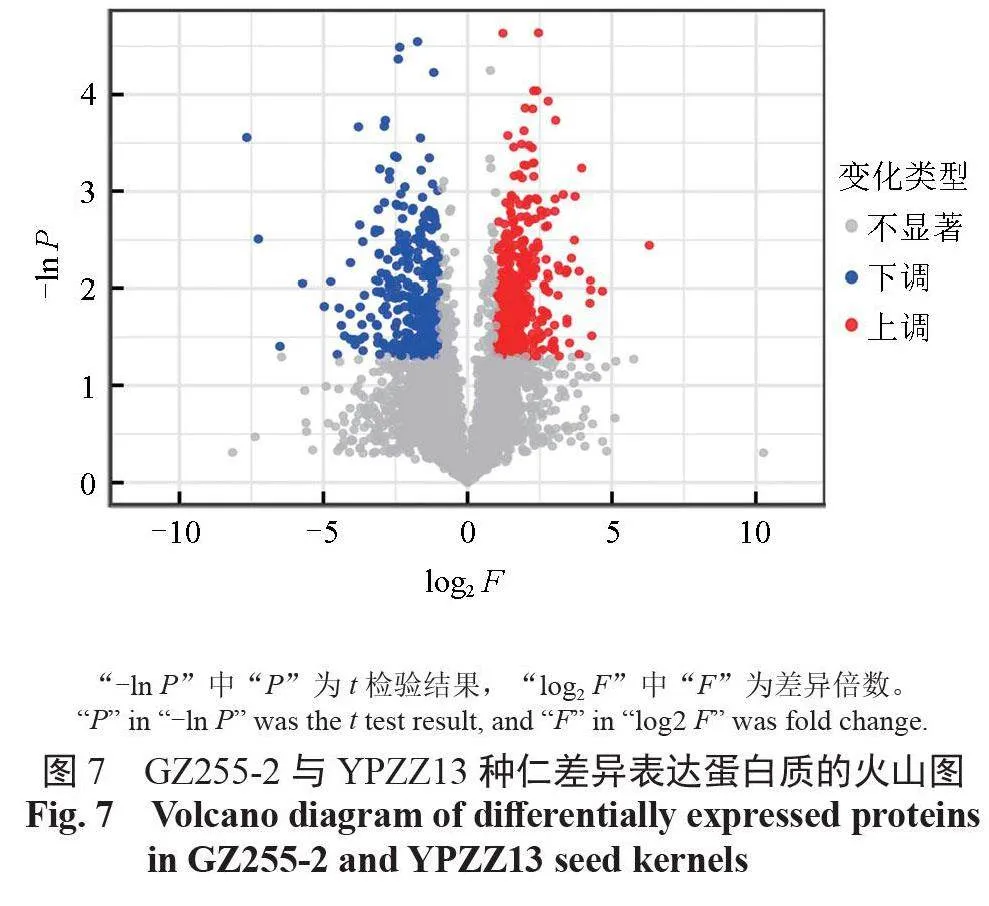
對在GZ255-2 和YPZZ13 種仁中鑒定出的5712個蛋白質進行差異表達蛋白(differentiallyexpressed proteins,DEPs)的篩選,將折疊變化的蛋白質進行t 檢驗得到P 值,篩選出顯著差異表達的蛋白質。與YPZZ13 種仁蛋白質相比,GZ255-2種仁中有403個上調的顯著差異表達蛋白質,有337 個下調的顯著差異表達蛋白質,有178 個上調的部分差異表達蛋白質,有68 個下調的部分差異表達蛋白質。GZ255-2 與YPZZ13 種仁中蛋白質表達的差異情況如圖7 所示。
2.4 種仁差異表達蛋白質的聚類
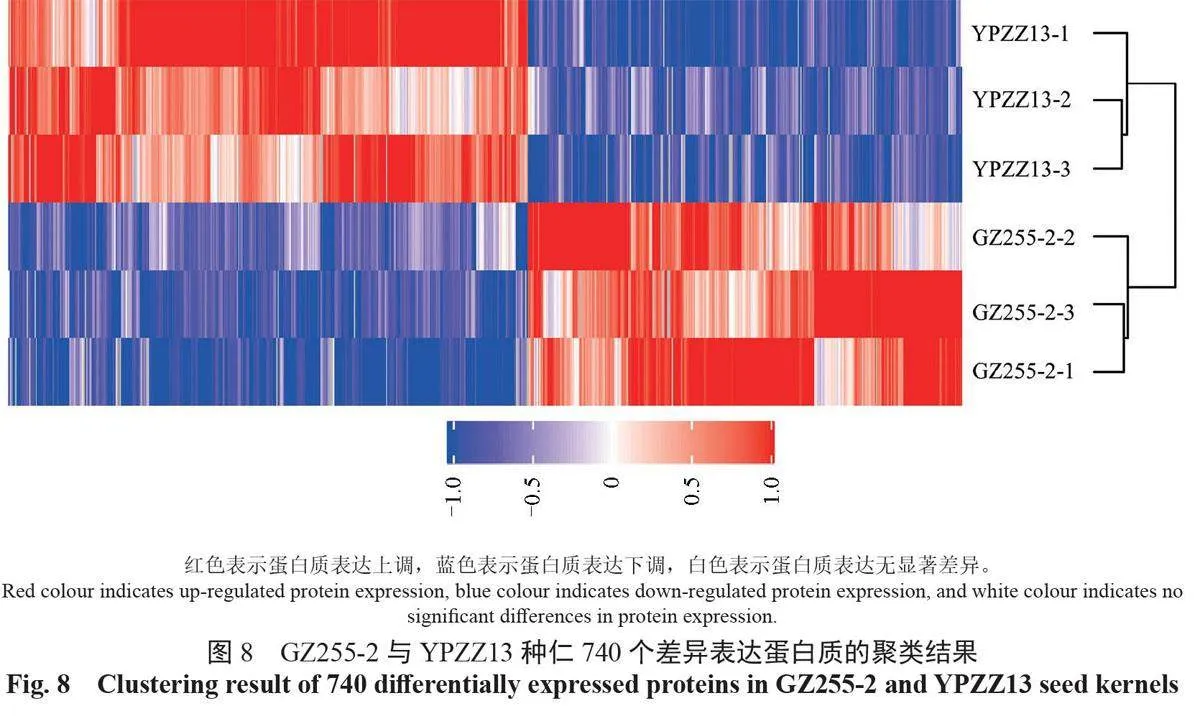
將GZ255-2和YPZZ13種仁中差異表達的740 個蛋白質采用層次聚類法進行聚類,結果如圖8所示。具有相似積累模式的蛋白質被聚在一起,展示了蛋白質的差異表達與生理功能、性狀的相關性。有403 個差異表達蛋白質在YPZZ13中上調,在GZ255-2 中下調;有337個差異表達蛋白質在YPZZ13 中下調,在GZ255-2中上調。高州油茶和小果油茶種仁中蛋白質聚類的結果表明,二者生理性狀的差異來源于蛋白質的積累模式差異。
2.5 種仁差異表達蛋白質的GO分析
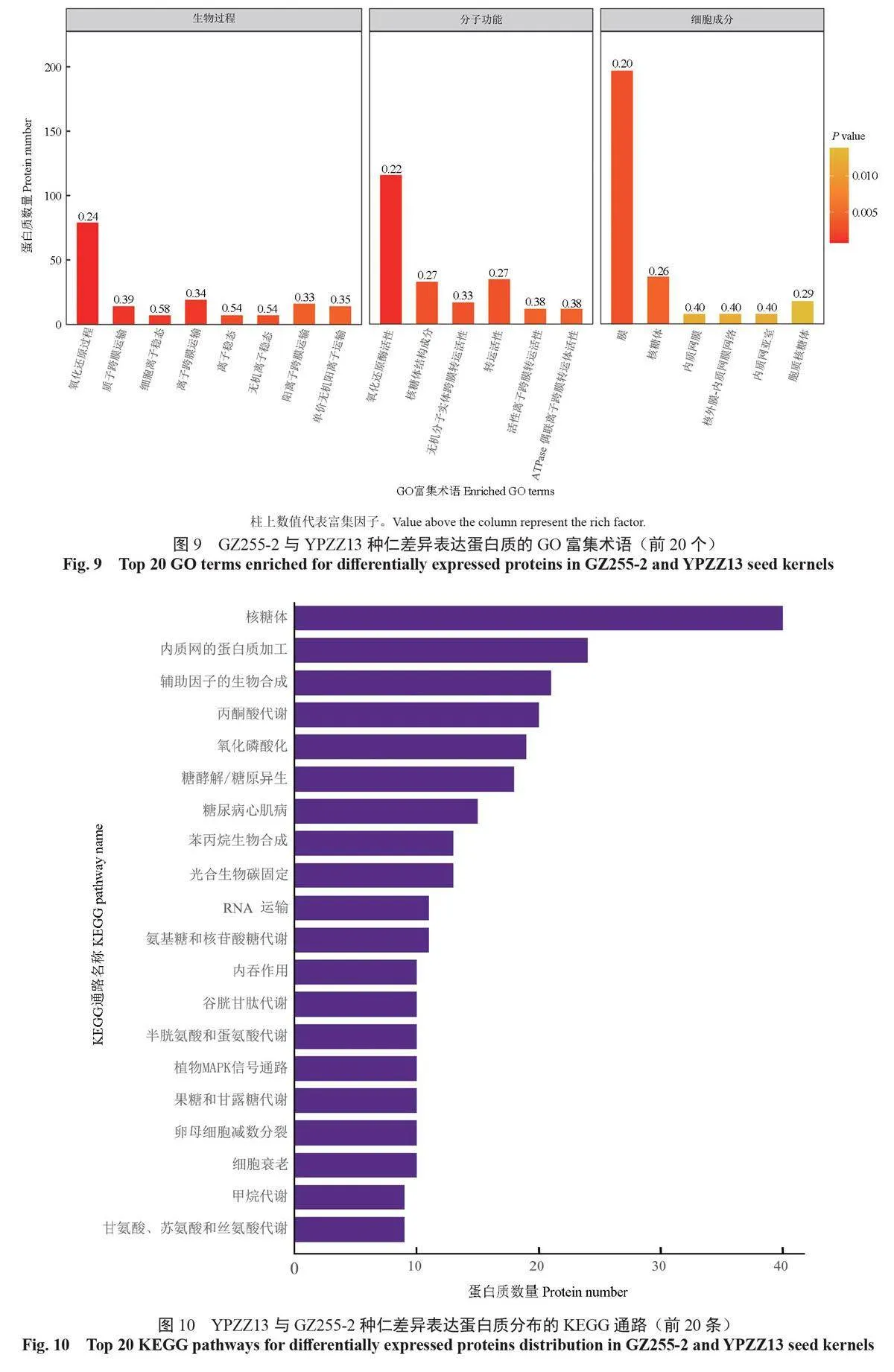
根據GO 數據庫對差異表達蛋白質進行生物過程(biological process,BP)、分子功能(molecularfunction,MF)和細胞成分(cellular component,CC)的分類,得到前20 個GO 功能富集術語(圖9)。生物過程方面,具有代表性的GO 術語是氧化還原過程(79 個DEPs)和質子跨膜運輸(14個DEPs);分子功能方面,氧化還原酶活性(116個DEPs)、核糖體結構成分(33 個DEPs)和無機分子實體跨膜轉運活性(17 個DEPs)是具有代表性的GO 術語;細胞成分方面,膜(197 個DEPs)和核糖體(37 個DEPs)是具有代表性的GO 術語。
2.6 種仁差異表達蛋白質的KEGG分析
2.6.1 差異表達蛋白質的KEGG通路注釋
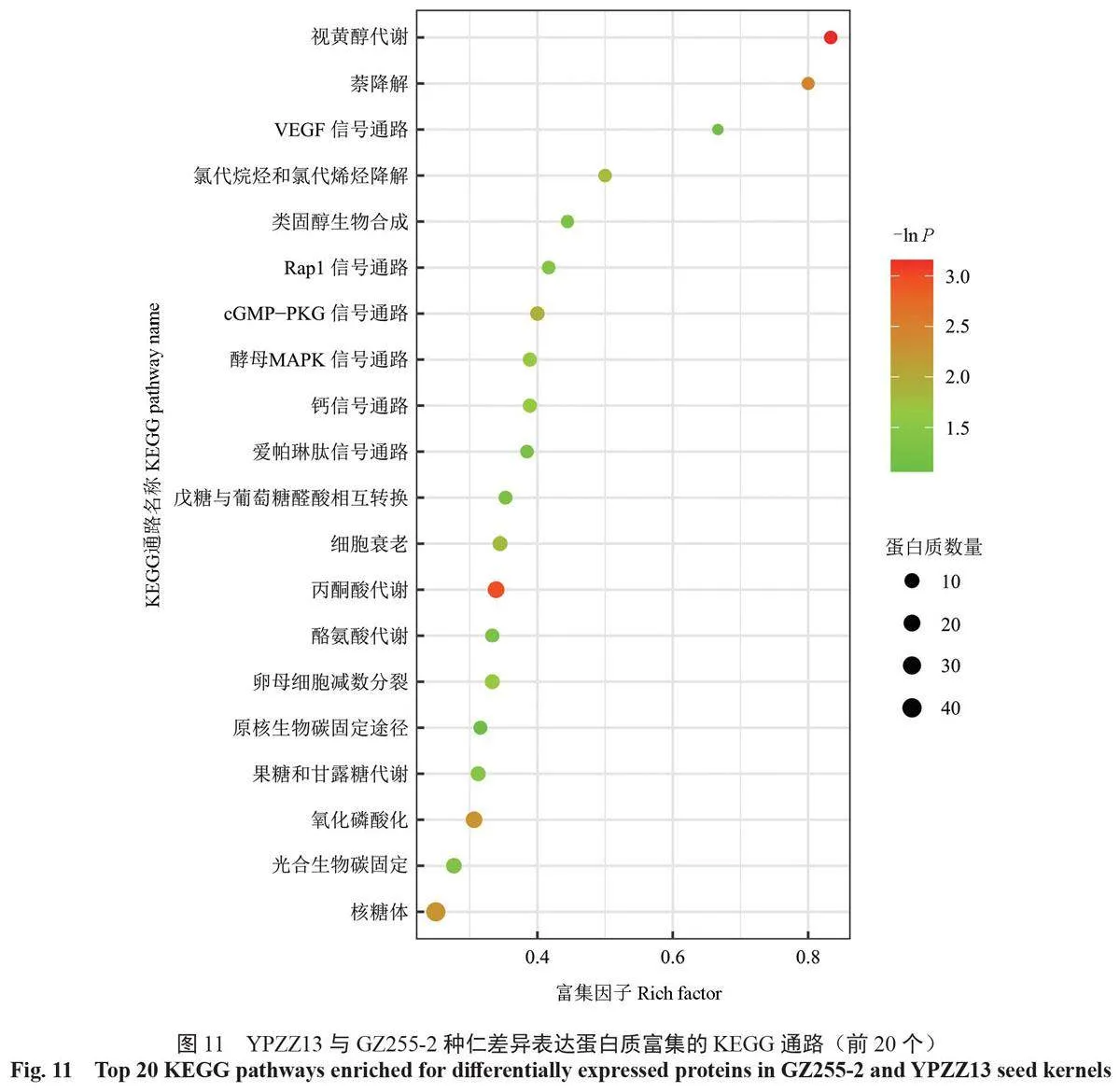
在GZ255-2 和YPZZ13種仁中共檢測出179條差異表達蛋白質富集的KEGG通路并進行注釋,篩選出差異表達蛋白質富集數量排名在前20 位的通路(圖10)。差異表達蛋白質富集的KEGG 通路中,富集數量最多的前5 條通路分別是核糖體通路(40個DEPs)、內質網的蛋白質加工通路(24個DEPs)、輔助因子的生物合成通路(21個DEPs)、丙酮酸代謝通路(20個DEPs)和氧化磷酸化通路(19個DEPs)。
2.6.2 差異表達蛋白質的KEGG富集分析
通過KEGG 富集分析, 篩選出GZ255-2 與YPZZ13 中差異表達蛋白質顯著富集的前20 條KEGG 富集通路(圖11)。視黃醇代謝通路(5個DEPs)、丙酮酸代謝通路(20 個DEPs)、萘降解通路(4 個DEPs)、氧化磷酸化通路(19 個DEPs)和核糖體通路(40 個DEPs)是顯著富集(P<0.05)的前5 條重要通路。其中,丙酮酸代謝通路、氧化磷酸化通路和核糖體通路是屬于差異表達蛋白質富集數量排名在前20 位的通路,而視黃醇代謝通路是差異表達蛋白質富集最顯著的通路。
2.7 種仁蛋白質互作的網絡分析
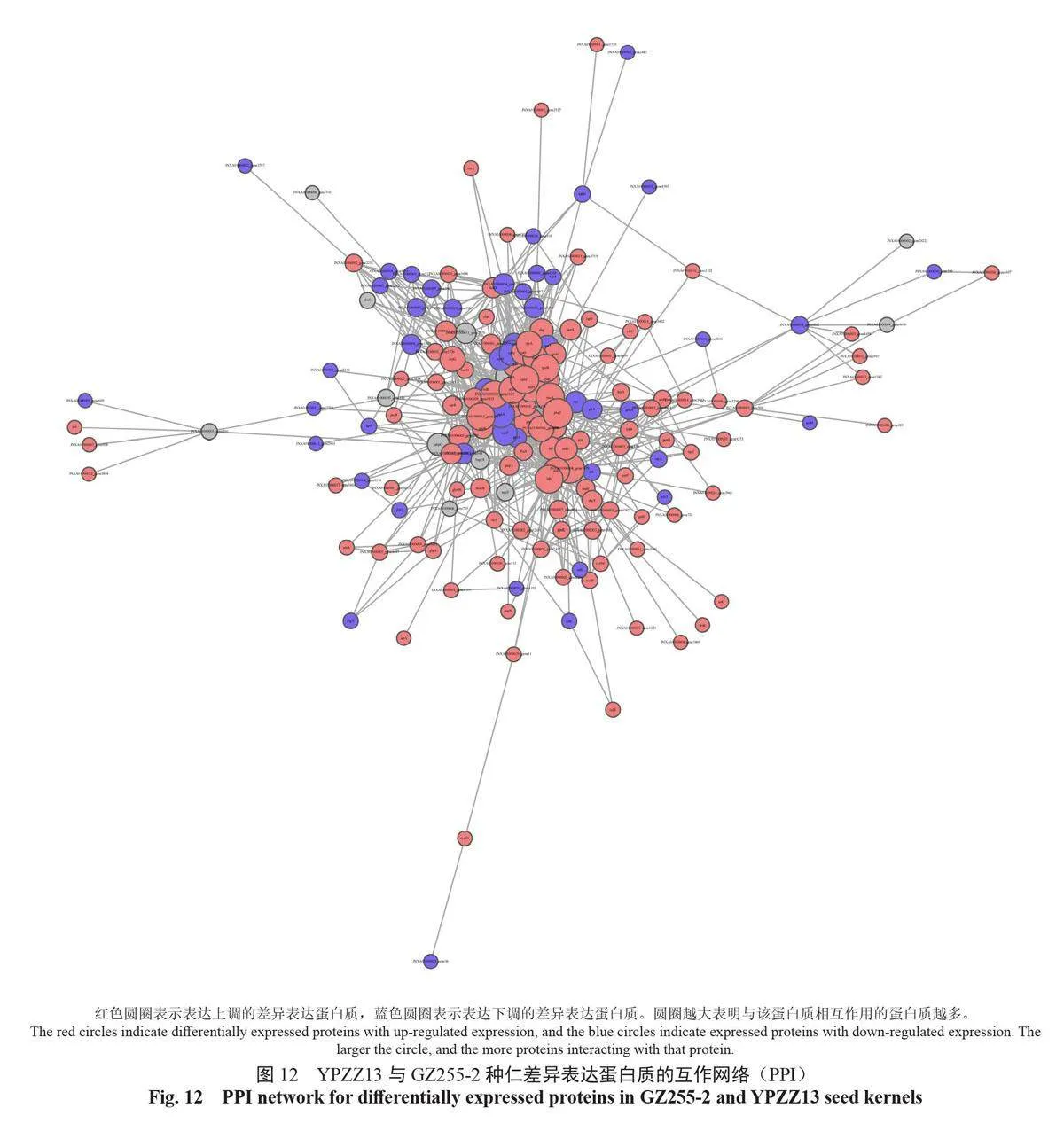
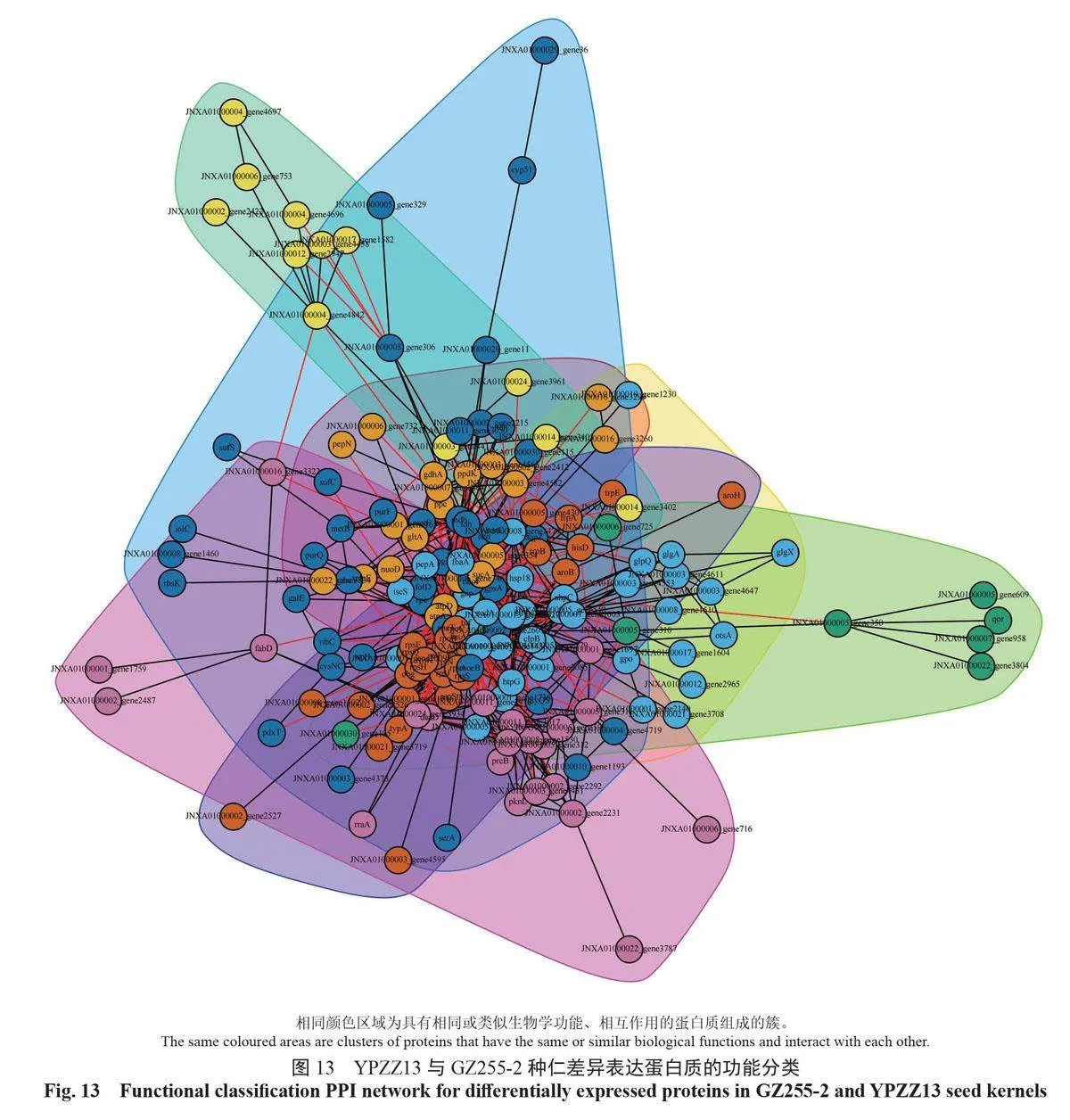
蛋白質互作網絡是蛋白之間介導的途徑,或其形成復合物從而發揮生物學調控作用的機制。以String 數據庫中的蛋白質相互作用關系為基礎,利用CytoScape 軟件構建GZ255-2 與YPZZ13 差異表達蛋白質的相互作用網絡圖,展示GZ255-2與YPZZ13 差異表達蛋白質之間的相互作用網絡(圖12)。不同蛋白質高度聚集說明其可能具有相同或相似的功能,能通過協同作用發揮生物學功能。將互作網絡中聚集程度高的蛋白質劃分為不同簇,進行功能歸類分組,如圖13所示。
2.8 種仁顯著差異表達蛋白質的代謝通路富集分析
通過比較高州油茶和小果油茶種仁中參與油脂合成積累、營養物質合成、活性成分合成等重要通路的顯著差異表達的蛋白質,篩選出部分關鍵的顯著差異表達蛋白質。
表1中展示了與油脂合成、積累相關的關鍵顯著差異表達蛋白質,及其參與的重要通路。在淀粉和蔗糖代謝、脂肪酸的生物合成、脂肪酸降解、脂肪酸延伸、不飽和脂肪酸的生物合成等通路中鑒定出多個關鍵的顯著差異表達蛋白質,其中長鏈酰基輔酶A 合成酶同時參與了脂肪酸的生物合成和脂肪酸降解通路,酰基-[ 酰基載體蛋白] 去飽和酶同時參與了脂肪酸的生物合成和不飽和脂肪酸的生物合成通路,烯酰基載體蛋白還原酶/ 反式-2-烯酰- 輔酶A 還原酶同時參與了脂肪酸的生物合成和脂肪酸延伸通路,而在脂肪酸降解通路中有3個S-( 羥甲基) 谷胱甘肽脫氫酶/ 乙醇脫氫酶。
維生素和人體不能合成的必需氨基酸在種仁中的表達情況是影響油茶種仁品質的重要因素。表2 中展示了與營養物質、活性成分合成相關的關鍵顯著差異表達蛋白質,及其參與的重要通路。在視黃醇代謝(維生素A)、抗壞血酸(維生素C)和醛酸代謝、硫胺素代謝(維生素B1)、核黃素代謝(維生素B2)、維生素B6代謝、生物素代謝(維生素B7、維生素H)等通路中共鑒定出14個關鍵的顯著差異表達蛋白質,其中參與視黃醇代謝通路的3個S-( 羥甲基) 谷胱甘肽脫氫酶/乙醇脫氫酶還參與了脂肪酸降解通路,參與生物素代謝的3- 羥酰基-[ 酰基載體蛋白] 脫水酶還參與了脂肪酸的生物合成通路。在必需氨基酸代謝和生物合成的不同通路中,共鑒定出16 個顯著差異表達蛋白質,其中酰胺酶同時參與了苯丙氨酸代謝通路和色氨酸代謝通路。
3 結論與討論
本研究中利用非標記蛋白質組學技術研究了小果油茶和高州油茶種仁的蛋白質組差異,發現740個蛋白質在2 種油茶種仁中呈現表達的差異。通過GO 分析發現,差異表達蛋白質主要參與氧化還原的過程以及核糖體的組成。KEGG 分析結果顯示,差異表達蛋白質顯著富集的通路是視黃醇代謝通路、丙酮酸代謝通路、萘降解通路、氧化磷酸化通路和核糖體通路,這與GO 分析結果相關。此外,通過對差異表達蛋白質進行挖掘,總結和分析了影響種仁中油脂合成積累、營養物質合成、活性成分合成相關的部分關鍵的顯著差異表達蛋白質,揭示了其在高州油茶和小果油茶種仁中積累模式的差異。
糖類物質是油脂積累的重要前體物質,可溶性糖和淀粉含量與油脂的積累正相關[28-30]。本研究結果表明,6個顯著差異表達的蛋白質包括淀粉合酶(glgA)、ADP- 葡萄糖焦磷酸化酶(glgC)、海藻糖-6- 磷酸合酶/ 磷酸酶(TPS)、UDP- 葡萄糖焦磷酸化酶(UGP2)、果糖激酶(scrK)和己糖激酶(HK)。其中,glgA 和glgC 是參與淀粉生物合成的關鍵酶,其基因表達水平與淀粉積累速率正相關[31-32]。TPS 參與海藻糖代謝,是合成海藻糖的關鍵酶,其酶活性影響植物新陳代謝、發育進程和脅迫應答的能力[33-35]。UGP2 則是多糖生物合成過程中的關鍵酶,參與調控植物體內蔗糖的合成與代謝[36],還參與催化生產UDP- 葡萄糖,從而合成蔗糖和促進細胞壁形成[37-38],UGP2 在植物發育和應答脅迫方面也具有重要的作用[39]。此外,Granot 等[40] 經鑒定發現,HK 家族和scrK 家族是催化葡萄糖和果糖基本不可逆磷酸化的2 個酶家族,游離果糖會被scrK 或HK 催化磷酸化,且scrK 與果糖的親和度更高[41]。與小果油茶相比,在高州油茶種仁中,這6 個顯著差異表達蛋白質在淀粉和蔗糖代謝通路中下調,這可能與高州油茶消耗了大量的糖類物質用于合成脂肪酸有關。莊瑞林[4] 經研究認為,在油脂合成和積累的能力上,高州油茶優于小果油茶,這與本研究結果基本一致,說明脂肪酸的合成和積累可能是影響糖類物質代謝的重要因素。
此外,本研究中發現長鏈酰基輔酶A 合成酶(ACSL)、烯酰基載體蛋白還原酶/ 反式-2- 烯酰- 輔酶A 還原酶(MECR)、酰基-[ 酰基載體蛋白] 去飽和酶(FAB2)參與了與脂肪酸代謝相關的多個通路。ACSL 通過形成酰基輔酶A 中間體激活脂肪酸,參與脂肪酸的生物合成、降解。ACSL 家族酶能將長鏈或超長鏈脂肪酸轉化為硫酯,用于不同的脂質代謝相關過程,而其活性的改變影響種子萌發、種子含油量以及植物應答脅迫的能力[42]。MECR 參與脂肪酸的生物合成和延伸,能促進脂肪酸的合成,并參與脂肪酸的延伸,使脂肪酸從C4 延伸到C16,甚至進一步拉長成為極長鏈脂肪酸,改變不同脂肪酸的相對含量[43-44]。FAB2 是一類可溶性酶,是參與不飽和脂肪酸和脂肪酸的生物合成的關鍵蛋白[45],其基因表達影響植物對生物脅迫調控的反應[46]。與小果油茶相比,高州油茶有9 個顯著差異表達蛋白質在脂肪酸的生物合成、脂肪酸降解和脂肪酸延伸通路中下調,有1 個顯著差異表達蛋白質在不飽和脂肪酸的生物合成通路中上調,這與糖類物質代謝促進脂肪酸合成的推測結果不一致。高州油茶作為性狀變異程度較大的大果油茶,其成熟時間常存在差異,采集時間較晚或種子成熟度較高有可能是導致種仁脂肪酸合成減弱的原因,但脂肪酸的降解也減弱,這與Wang 等[47] 的報道一致,即早熟大果油茶種子成熟末期,脂肪酸的含量先增加,后略有減少。此外,與小果油茶種仁相比,高州油茶種仁中FAB2 在不飽和脂肪酸的生物合成通路中上調,推測在種仁成熟末期,高州油茶種仁中不飽和脂肪酸的合成和積累增加,這與高州油茶油脂高不飽和脂肪酸含量的特征一致。
視黃醇是維生素A中的一種[48],與視覺功能、細胞分化和生長、上皮組織細胞健康、免疫功能及抗氧化能力等相關[49]。視黃醇通常與脂肪酸結合成視黃基酯,視黃基酯和植物性食物中的類胡蘿卜素又常與蛋白質結合形成復合物,最終在小腸中釋放出脂肪酸、游離的視黃醇及類胡蘿卜素[50]。KEGG 富集分析的前20條通路中,差異表達蛋白質最顯著富集的通路是視黃醇代謝通路。在視黃醇代謝通路中,共有4個顯著差異表達蛋白質富集,其中1個為脫氫酶/ 還原酶SDR家族成員4(DHRS4),另外3個顯著差異表達蛋白質均為S-( 羥甲基) 谷胱甘肽脫氫酶/ 乙醇脫氫酶(frmA)。其中,DHRS4 屬于短鏈脫氫酶/ 還原酶(SDR)大家族,SDR 是NAD(P)H 依賴性氧化還原酶,在脂質、氨基酸、糖、輔助因子、激素和異生物代謝以及氧化還原傳感機制中發揮著重要作用[51]。SDR基因在種子萌發過程中顯著表達,參與了種子的發育[52]。乙醇脫氫酶則參與視黃醇代謝,催化視黃醇和視黃醛的可逆相互轉化,視黃醛經還原得到視黃醇,經氧化后得到視黃酸[53]。視黃酸是視黃醇代謝的中間產物,在調節細胞分化和凋亡、胚胎發育和再生、視力發育等方面發揮重要作用[54-55]。與小果油茶種仁相比,高州油茶種仁中顯著差異表達蛋白質在視黃醇代謝通路下調,說明小果油茶種仁的維生素A含量可能較高。
試驗中筆者采樣時對油茶果實成熟度的判斷可能存在一定的偏差,這會對油茶種仁蛋白質的鑒定產生一定的影響。后續將進一步分析關鍵蛋白(酶)的調控基因和最終合成的代謝產物,以驗證本研究中的差異蛋白數據。
