三種濕地植物在高負荷養豬廢水脫氮過程中的根際效應
2024-11-07 00:00:00夏夢華劉銘羽葉磊李希李裕元吳金水
農業環境科學學報 2024年4期
關鍵詞:養豬廢水;人工濕地;氮;根系分泌物;根際微生物
農業面源污染已成為制約農業經濟和農村生態環境可持續發展的主要因素。其中,畜禽養殖廢水因排水時間較為集中,污染負荷較高等原因,難以快速地控制和利用。目前,養殖廢水處理技術主要包括二級處理工藝(預處理+活性污泥法/生物膜法)和生態環境工程法,生態環境工程投資相對較少,且資源回收利用率較高,因此更受利潤較低且易被市場影響的中小型養殖企業青睞。其中人工濕地組合模式較多,可滿足不同的污染消納需求,因此應用前景更加廣泛。
人工濕地對污水中氮的去除依靠微生物、植物和填料基質間的協同作用,微生物作用目前被認為是人工濕地中最主要的脫氮機制,氮脫除量可達到系統總量的43%-95%,植物同化貢獻率約為0.5%-60%。除了直接吸收氮素以外,植物根系還可以向根際釋放分泌物,根系分泌物不僅可以誘導和刺激根際特定菌群的生長,還可以作為有效碳源被根際微生物利用,從而使根際微生物的豐度及種類遠高于非根際,進而影響濕地的脫氮過程及脫氮效果。例如Zhai等研究顯示,在低污染負荷的潛流人工濕地中,植物根系分泌物可能會使反硝化速率增加94-267kg·hm-2·a-1(以N計);Salvato等發現香蒲和蘆葦根系能分泌更多的溶解性有機碳,反硝化率分別達到37%和34%,顯著高于其他試驗植物;Wu等研究表明根系分泌的低分子量有機碳能顯著提高根際反硝化基因(nirS和nirK)的豐度;Gu等研究表明水生植物的存在有利于氨氧化菌的生長繁殖。
目前關于濕地植物根系分泌物的釋放速率與種類對濕地系統氮去除效果及根際微生物群落組成影響的研究已較多,但是在污染負荷較高的養豬廢水中植物根系分泌物的釋放特征及其對根際脫氮過程影響的研究還較少。已有研究表明綠狐尾藻、鳳眼蓮和美人蕉等水生植物可在高負荷養豬廢水中正常存活且脫氮效果可觀,表明利用水生植物構建人工濕地治理養豬廢水具有可行性。基于此,本文通過野外小區控制試驗,綜合考慮耐受性及生長特性后,選擇美人蕉、梭魚草和黃菖蒲3種挺水植物為試驗對象,構建表面流人工濕地,研究3種濕地植物在處理養豬廢水過程中根系分泌物對根際氮循環功能基因豐度的影響,以期揭示濕地植物根際脫氮機理,探尋提高人工濕地脫氮能力的途徑,為高負荷養豬廢水的生態修復提供理論依據。
1材料與方法
1.1試驗區概況
試驗區位于中國科學院長沙農業環境觀測站(28°33' 4.01\"N,113°20' 4.75\"E),屬亞熱帶季風氣候,四季分明,年平均氣溫17℃,年平均降雨量1470mm,年平均無霜期為260d。試驗期間月平均溫度21℃,月平均降雨量100mm。
1.2試驗材料
試驗所用養豬廢水取自長沙縣開慧鎮錫福村某豬場,豬場占地面積約為5800m2,生豬存欄量約為1500頭。廢水為經過厭氧處理的沼液,其總氮濃度為294mg·L-1。試驗所選植物為當地較為典型且萌蘗能力較強的挺水植物,幼苗購于附近花卉市場,挑選株高大致相同且生長狀態良好的植株作為試驗材料。小區內填料基質選用當地典型水稻土,土壤砂粒(gt;0.05mm)含量為42%,黏粒(lt;0.002mm)含量為10%,質地為粉砂壤,土壤pH為5.2,全氮、全磷、全鉀和總有機質含量分別為1.2、0.4、22.5g·kg-1和18.3g·kg-1。
1.3試驗設計
試驗時間為2018年9-11月,為野外小區控制試驗,共4組處理,分別為美人蕉、梭魚草、黃菖蒲植物組及無植物對照組,每組設置3次重復,共12個試驗小區,小區長80cm、寬50cm、深40cm。為使小區正常排水,在一側墻壁距池底28cm的位置開兩個半徑為1cm的排水孔。小區內定植基質(水稻土)填裝厚度為12cm,且均已均質過篩(3mm),每個小區種植幼苗15叢,并控制其鮮質量保持一致(約1.6kg)。緩慢加入清水至剛剛淹沒土壤,穩定10d。待植物生長穩定后一次性加廢水至排水孔處,記作試驗正式開始的第1天,后使用兩臺蠕動泵(LabV3)從儲水桶向12個試驗小區同時泵入廢水,進水流速均為6.33mL.min-1,全天不間斷供水,共運行56d。各小區進水量為9.14L·d-1,即人工濕地表面水力負荷為0.02m3.m-2.d-1,水力停留時間為7dm,試驗期間根據自然狀況定期加水以補充因蒸散發而損失的水分。
1.4取樣與檢測
分別在人工濕地運行的第8、16、24、32、40、48天和56天采集水樣,共采集7次,沿小區對角線取3份水樣,混合后儲于100mL塑料瓶中,同時采集儲水桶中原液,過濾(0.45um)后取上清液待測。銨態氮(NH4-N)和硝態氮(N03-N)使用AA3流動分析儀直接測定(德國seal),總氮(TN)采用堿性過硫酸鉀消解后使用AA3流動分析儀測定。使用便攜式水質分析儀(美國HQ40d)測定水溫、pH及溶解氧(DO)。
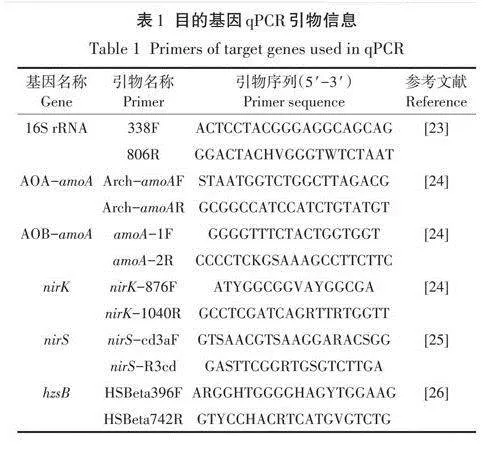
分別于試驗前(穩定期后,養豬廢水添加前)和試驗后(培養56d)對分泌物和根際微生物進行收集。每個小區選擇2株生長較好且具有代表性的植株,測定植物株高后將植物樣品帶底泥切出,裝在滅菌袋中,低溫保存并快速送回實驗室。清理根系附著的大量底泥后,用玻璃棒將根際土輕輕刮下,用錫紙包裹后放人紗網袋中-80℃保存。底泥樣品中微生物DNA用DNeasy PowerSoil kit(德國QIAGEN)試劑盒提取,用NanoDrop核酸蛋白測定儀(美國Thermo)測定DNA濃度。分別以16S rRNA、氨氧化古菌(AOA)和氨氧化細菌(AOB)的氨單加氧酶編碼基因(amoA)、亞硝酸鹽還原酶編碼基因(nirK、nirS)和聯氨合成酶編碼基因為目的基因進行qPCR(美國Roche)擴增。對含有目的基因且濃度已知的質粒載體進行梯度稀釋,制備成標準曲線,將標準曲線及樣品同時置于384孔板中進行擴增,默認標準曲線R2大于0.99,目的基因擴增效率在90%-110%時數據可用。擴增體系為10uL,其中DNA樣品1uL,上下游引物0.4uL,Takara SYBR Premix Ex Taq(2x)5uL.用滅菌去離子水補足10uL。qPCR反應條件:95℃預變性2min;隨后95℃變性Ss;57℃退火20 s;72℃延伸20 s。反應共進行40個循環,具體引物信息見表l。
取完根際土后,用超純水將植物根系表面清洗干凈,再用濾紙吸干表面水分,放入0.5mmol·L-1的CaCl2培養液中(已滅菌)室溫培養24 h,培養結束后采集100mL培養液,過0.45um有機相濾膜(Nvlon材質)純化后使用有機碳分析儀(日本島津)測定培養液中溶解性有機碳(DOC)含量。再取一份50mL培養液,冷凍干燥至近干后過0.22um有機相濾膜純化,用高效液相色譜儀(美國Agilent)測定培養液中乙酸(AA)、檸檬酸(CA)、琥珀酸(SA)及丙二酸(MA)的含量,4種有機酸(OA)含量總和計為總有機酸(TOA)含量。收集完分泌物后將植物在80℃下烘干至質量恒定,記錄植株生物量及根系生物量。植物體內全氮含量經H2S04-H202消解后使用AA3流動分析儀測定。
1.5數據計算與分析
人工濕地中污染物去除效率計算公式:
采用Excel 2010進行數據分析;采用SPSS 22進行單因素方差分析,差異顯著(Plt;0.05)則進一步進行多重比較;采用R語言4.1.3中的ggplot2包繪制柱狀圖,corrplot包繪制Pearson相關性熱圖。
2結果與分析
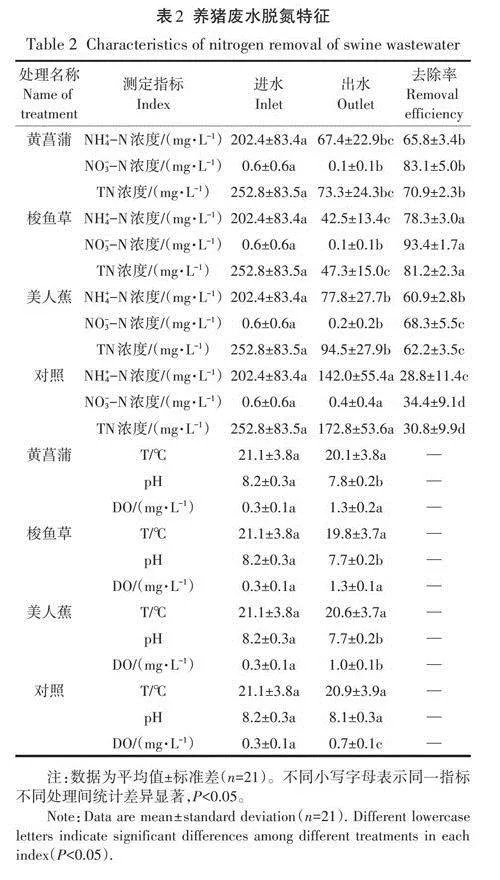
2.1養豬廢水脫氮特征
試驗期間人工濕地系統中水溫呈下降趨勢,變化范圍為14.3-27.5℃,與氣溫(13.7-30.7℃)變化趨勢一致,不同處理間水溫無顯著差異(Pgt;0.05),詳見表2。水體呈微堿性,植物處理組pH均值為7.7,對照組pH均值為8.1,植物處理組pH顯著低于對照組(Plt;0.05)。水體中DO含量偏低,植物處理組DO均值為1.2 mg·L-1,對照組DO均值為0.7mg·L-1,植物處理組DO顯著高于對照組(Plt;0.05)。試驗期間NH;-N、N03-N和TN進水濃度變化范圍分別為127.0~384.8、0.2-1.6mg·L-1和170.0-429.9mg·L-1,養豬廢水中N的主要形態為NH;-N,N03-N含量極低。植物處理組對NH;-N、N03-N和TN的平均去除率分別為68.3%、81.6%和71.4%,出水濃度為62.6、0.1mg·L-1和71.7mg·L-1,效果均顯著優于對照組(Plt;0.05)。試驗期間梭魚草處理脫氮效果最好。
2.2根際氮循環功能基因的定量分析
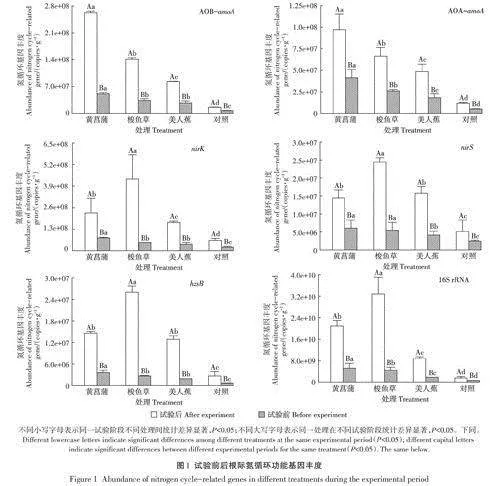
試驗期間人工濕地系統中AOB-amoA基因豐度顯著高于AOA-amoA(Plt;0.05),詳見圖1,表明濕地系統中氨氧化過程AOB占主導地位。植物組根際土中AOB-amoA基因豐度為2.8x107~2.6x108 copies·g-1,對照組中為7.1x106~1.6x107copies,根際土中AOB-amoA基因豐度顯著高于非根際土(Plt;0.05)。4組處理中AOB-amoA基因豐度試驗前為7.1x106~5.1x107copies·g-1,試驗后為1.6x107~2.6x108 copies·g-1,試驗后AOB-amoA基因豐度顯著高于試驗前(Plt;0.05)。AOB-amoA基因豐度在黃菖蒲處理中最高,試驗前為5.1X107 copies·g-1,試驗后為2.6x108 copies·g-1。
試驗期間人工濕地系統中nirK基因豐度顯著高于nirS (Plt;0.05),可見根際N02-N還原過程主要靠nirK編碼的還原酶催化轉化。試驗期間nirK基因豐度變化范圍為2.1x107~4.3x108 copies·g-1,同樣地,根際土中nirK基因豐度顯著高于非根際土(Plt;0.05),試驗后nirK基因豐度顯著高于試驗前(Plt;0.05)。nirK基因豐度試驗前在黃菖蒲處理中最高,為7.7x 107copies.g-1;試驗后在梭魚草處理中最高,為4.3x108 copies·g-1。
試驗期間hz.sB基因豐度變化范圍為6.4×105-2.6x107 copies·g-1,說明在植物根際也存在較明顯的厭氧氨氧化反應。
2.3植物根系分泌物釋放特征
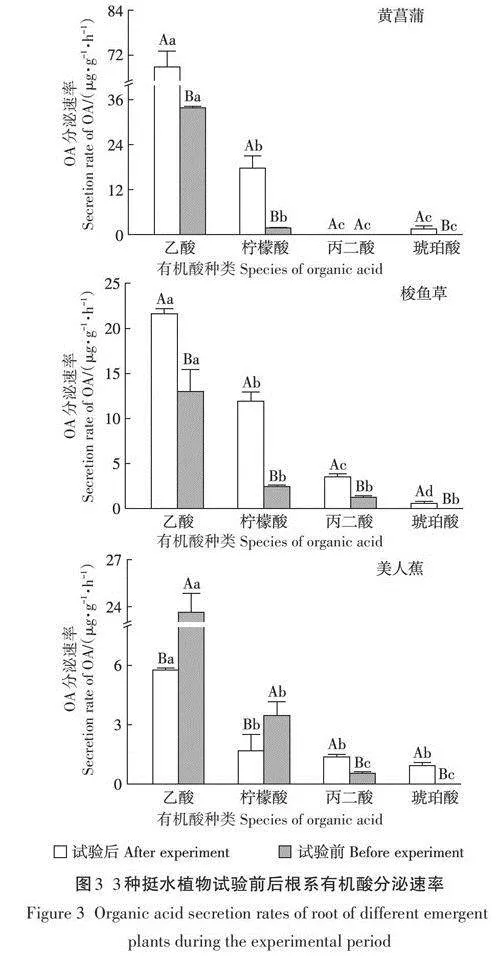
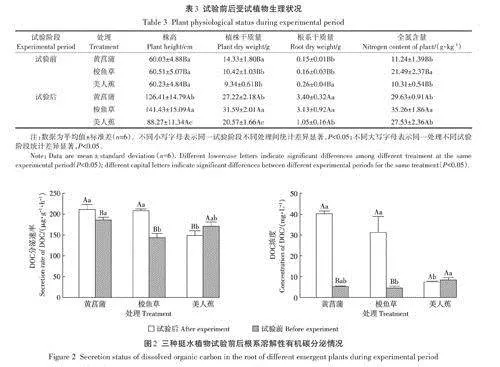
試驗前后受試植物生理狀況如表3所示,可見梭魚草生物量較大,黃菖蒲根系較發達。試驗前植物根系DOC分泌速率黃菖蒲gt;美人蕉gt;梭魚草(圖2),分別為185.8、170.6ug·g-1.h-1和143.3ug·g-1.h-1,黃菖蒲與梭魚草根系DOC分泌速率差異極顯著(Plt;0.01);試驗后分泌速率為黃菖蒲gt;梭魚草gt;美人蕉,分別為210.9、207.7ug·1-h-1和148.8ug·g-1.h-1,黃菖蒲、梭魚草與美人蕉根系DOC分泌速率差異極顯著(Plt;0.01)。黃菖蒲和梭魚草根系DOC分泌速率在試驗后顯著升高,美人蕉則是顯著降低(Plt;0.05)。培養液中DOC濃度試驗前在美人蕉處理中最高,為8.4mg·L-1;試驗后在黃菖蒲處理中最高,為40.2mg·L-1,培養液中DOC濃度與根系生物量呈正相關(Plt;0.05)。
試驗前在美人蕉和梭魚草根際檢測出AA、CA和MA,TOA分泌速率分別為黃菖蒲根際檢測出AA和CA,TOA分泌速率為35.6ug·g-1·h-1,如圖3所示。試驗后三種植物根際均新增檢測出SA,黃菖蒲和梭魚草根系TOA分泌速率試驗后顯著高于試驗前(Plt;0.05),分別為88.1而美人蕉根系TOA分泌速率試驗后顯著低于試驗前(Plt;0.05),試驗期間黃菖蒲根系TOA分泌能力最強,梭魚草根系分泌有機酸的種類最多,且CA、SA和MA的分泌能力均較強。
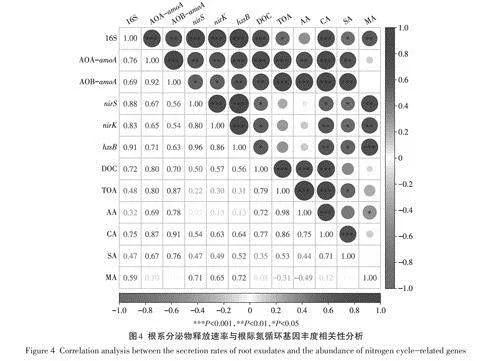
2.4根系分泌物與氮循環功能基因豐度的相關性
植物根系分泌物釋放速率與根際氮循環基因豐度的Pearson相關性分析如圖4所示,結果表明根系DOC分泌速率與5種氮循環功能基因豐度均呈正相關(Plt;0.05),可見根系分泌DOC對根際硝化菌、反硝化菌及厭氧氨氧化菌的生長有著促進作用。TOA分泌速率與AOA和AOB基因豐度呈顯著正相關(Plt;0.01),表明根系分泌TOA對硝化菌的生長影響更大。CA、SA也與5種氮循環功能基因豐度均呈正相關(Plt;0.05),可見有機酸種類不同,對根際氮循環微生物的影響也不同。
3討論
3.1影響養豬廢水脫氮特征的主要因素
試驗期間植物處理組pH顯著低于對照組,這是因為相比于對照組,植物處理組脫氮效果更好,NH4-N濃度相對較低;再者,植物根系有機酸等物質的分泌、微生物轉化脫氮等生理活動也會對水體pH產生一定影響。濕地系統內水體DO含量整體較低可能是因為微生物在分解有機污染物的過程中消耗了溶解的氧氣。缺氧環境有利于反硝化過程的進行,因此水體中N03-N濃度整體較低。植物處理組中DO含量明顯高于對照組,除了可能因為其對污染物的去除效果更好,致使大氣復氧速率增強以外,還可能因為植物根系具有泌氧能力。
試驗期間植物處理組對NH4-N、N03-N和TN的去除效果均高于無植物對照組,與陽光等研究結果一致,這是因為植物可以吸收廢水中的氮以供自身生長,研究表明NH4-N、N03-N及溶解態有機氮均能直接被植物吸收。再者,植物根系可以為微生物提供一個更適宜生長的環境,加快根際的氮轉化進程。先前已發表的關于養豬廢水中氮去向的研究表明在3種植物構建的人工濕地系統中,微生物轉化途徑脫氮量為1.9-2.7g·m-2·d-1,顯著高于對照組(1.4g·m-2.d-1) (Plt;0.05),可見植物的存在可以提高微生物轉化途徑的脫氮效能。梭魚草處理脫氮效果最好,不僅是因為其生物量較大,可吸收儲存更多氮素,也可能因為其根際細菌豐度及氮循環功能基因豐度較高。
3.2驅動根際氮循環功能基因豐度產生差異的環境因子
根系分泌DOC對根際氮循環微生物的生長有著促進作用,這是根際土中氮循環基因豐度高于非根際土的主要原因之一,與先前學者的研究結論一致。試驗后人工濕地系統中氮循環基因豐度升高,表明系統中氮轉化過程加快。氨氧化作為硝化過程的第一步和限速步驟,主要由AOA和AOB催化轉化,濕地系統中氨氧化過程主要靠AOB來完成,與Chen等、Di等研究結果一致,其原因可能是AOA銨耐受性較差。AOA和AOB均在黃菖蒲處理中最高,這可能是因為黃菖蒲根系DOC及TOA分泌能力較強。Ajibade等研究表明外源碳的添加可以為氨氧化細菌創造更合適的生存環境;Gu等的研究也表明,部分氨氧化細菌和植物根部可形成共生關系,根系分泌有機碳可促進這部分細菌的生長代謝。其次黃菖蒲根系生物量較大,可以擴大污染物與微生物的接觸面積。
nirS和nirK是亞硝酸鹽還原酶的編碼基因,可將N02轉化為NO,是反硝化過程的限速步驟,該人工濕地中的N02的還原過程主要靠nirK編碼的還原酶催化轉化,與Dandie等研究結果一致,型基因結構差異較大,在環境中也占據不同的生態位,濕地土壤中pH可能是驅動nirK和nirS基因豐度產生差異最主要的原因。nirS和nirK基因豐度均在梭魚草處理中最高,結合有機酸檢測結果來看,可能是因為梭魚草根系分泌的有機酸種類較多,而反硝化菌對碳源的利用具有選擇性。
hzsB是負責編譯聯氨合成酶的一個亞基,該酶可催化NO與NH4轉化為N2H4,是厭氧氨氧化過程的步驟之一,在缺氧環境較為常見且效能較高。研究結果表明厭氧氨氧化也是該濕地系統中一個重要的脫氮途徑,相比于Zhang等的研究,本研究中hzsB基因豐度較高,這可能是因為養豬廢水環境更適合厭氧氨氧化菌生長。
3.3影響根系DOC和OA分泌特征的主要因素
本研究表明黃菖蒲根系分泌DOC的能力最強,與Zhai等研究結果一致,這可能是因為植物的物理特征和根系分布策略導致的。試驗后期黃菖蒲與梭魚草根系DOC分泌速率增大,可能是因為根系DOC的釋放受外界營養狀況的強烈影響,Xu等研究表明在閾值范圍內,NH4-N的存在會促進根系DOC的釋放。試驗后期美人蕉根系DOC分泌速率降低,這可能是因為高銨環境破壞了美人蕉的根系結構或抑制了根系的生理代謝。本研究結果與馬濤等的研究結果存在一定差異,這可能是因為植物生長在野外,外部環境因素、苗齡均對DOC分泌速率有較大影響。
OA屬于小分子初級代謝物,一般通過擴散或離子通道轉移到根際環境,不受植物本身控制。試驗后所檢出的有機酸種類更多,這可能是由于試驗結束時植物正處于生長相對旺盛的階段,有相關研究表明植物所處的生長階段是導致根系分泌物種類存在差異的主要原因。其次,根系OA的分泌可能也受外界營養狀況的影響。CA和SA作為三羧酸(TCA)循環過程中的中間代謝產物,后期分泌速率加快可能意味著植物根系營養物質代謝速率增加。梭魚草CA、SA和MA的分泌能力均較好,這可能是其根際氮循環功能基因豐度較高的原因之一。
3.4根系分泌物與根際微生物相互作用關系
根系DOC分泌速率與試驗中5種氮循環功能基因豐度均呈正相關,這可能是因為根系分泌DOC組分較為復雜,可滿足不同類型氮循環微生物對碳源的需求。TOA與AOA、AOB相關性較強,表明TOA可以促進硝化菌在根際的活性,這可能是因為濕地系統為缺氧環境,相比于反硝化菌而言,硝化菌的生長受限,而根系分泌TOA可以為硝化菌的生長提供一個更適宜的環境,也可以為其代謝和繁殖提供能量。CA和SA與5種氮循環功能基因豐度呈正相關,可能是因為其作為TCA循環的中間產物,分泌速率越快表明根系生理代謝越強烈,進而植物根系對根際氮循環微生物的影響也越強烈;再者,CA可提高其他營養物質的生物可利用性,間接促進氮循環微生物的生長。
4結論
(1)3種挺水植物在處理養豬廢水(沼液)過程中具有不同的脫氮特征,其中梭魚草對氮的脫除能力最好,根際反硝化菌和厭氧氨氧化菌豐度最高;黃菖蒲脫氮能力次之,根際硝化菌豐度最高。
(2)濕地系統中微生物介導的脫氮過程以硝化一反硝化為主,同時伴隨著明顯的厭氧氨氧化作用。硝化過程AOB-amoA占主導地位,反硝化過程nirK占主導地位。
(3)黃菖蒲根系溶解性有機碳和總有機酸分泌能力最強,梭魚草根系分泌有機酸種類較多。根系分泌物可以促進根際氮循環微生物的生長繁殖,進而提高人工濕地系統的脫氮效率。