組蛋白去乙酰化酶抑制劑MGCD0103對小反芻獸疫病毒體外復制的影響
2024-11-08 00:00:00鄧瑞雪潘春容朱學亮胡林杰孫躍峰曾巧英蒙學蓮
畜牧獸醫學報 2024年10期
摘 要: 旨在探究組蛋白去乙酰化酶(HDACs)選擇性抑制劑MGCD0103(Mocetinostat)對小反芻獸疫病毒(peste des petits ruminants virus,PPRV)在山羊子宮內膜上皮細胞(EEC細胞)復制的影響。本研究通過RT-qPCR法測定PPRV感染后宿主細胞HDACs和抑制劑處理細胞中PPRV N基因的轉錄水平,通過Time of Addition Assay確定MGCD0103在PPRV復制過程的作用階段,用Western blot檢測PPRV N蛋白的相對表達量,并采用TCID50方法測定了病毒滴度。結果表明,PPRV感染引起EEC細胞HDAC1(Plt;0.001)和HDAC2(Plt;0.002)轉錄水平明顯上升;MGCD0103在病毒入侵和復制階段均發揮了抑制作用,且在復制階段抑制作用更明顯;MGCD0103顯著降低PPRV N基因轉錄水平和表達水平及病毒滴度(Plt;0.002),且其抑制PPRV在EEC中的半數有效濃度(EC50)和藥物選擇指數(SI)分別是0.43 μmol·L-1和4.35。本研究確定MGCD0103顯著抑制PPRV的復制,這對研發有效抗PPRV藥物具有重要意義,為高效率產毒細胞株的構建提供新思路。
關鍵詞: 組蛋白去乙酰化酶抑制劑;MGCD0103;抑制;小反芻獸疫病毒;病毒復制
中圖分類號:R978.7;S852.659.5
文獻標志碼:A
文章編號:0366-6964(2024)10-4542-11
收稿日期:2023-12-11
基金項目:2022年甘肅省動物用生物制品創新聯合體項目(22ZD6NA012-1);蘭州國家高新區“百園”行動專項
作者簡介:鄧瑞雪(1990-),男,甘肅臨洮人,博士,主要從事病毒與分子免疫學研究作,Email:drxd1@163.com
*通信作者:曾巧英,主要從事畜禽重大疫病病原微生物的分子致病機理及其免疫學研究,E-mail: zengqy@gsau.edu.cn;蒙學蓮,主要從事動物病毒分子生物學與免疫學研究,E-mail: mengxuelian@caas.cn
Suppressive Effect of Histone Deacetylase Inhibitor MGCD0103 on Peste Des Petits Rumants
Virus Replication in vitro
DENG "Ruixue1,2,3, PAN" Chunrong1,2,3, ZHU" Xueliang2, HU" Linjie2, SUN" Yuefeng
2, ZENG" Qiaoying1*, MENG" Xuelian2*
(1.College of Veterinary Medicine, Gansu Agricultural University, Lanzhou 730070," China;
2.
State Key Laboratory for Animal Disease Control and Prevention, Lanzhou Veterinary Research Institute,
Chinese Academy of Agricultural Sciences, Lanzhou 730046," China;
3.China Agricultural Vet Biology
and Technology Co., Ltd, Lanzhou 730046," China)
Abstract:" In this study, the effect of MGCD0103 (Mocetinostat), a selective HDACi, on the replication of peste des petits ruminants virus (PPRV) in goat endometrial epithelial cells (EEC cells) was investigated, aiming to clarify the role and activity of MGCD0103 in PPRV multiplication. The transcription levels of HDACs and PPRV N gene in different treated cells were determined by RT-qPCR, and the action stage of MGCD0103 in the viral proliferation cycle of PPRV was determined by the time of addition assay. The effect of MGCD0103 on the expression level of PPRV N protein and viral titer were further analyzed by Western blot and TCID50, respectively. The results showed that the mRNA expression levels of HDAC1 (Plt;0.001) and HDAC2 (Plt;0.002) were increased significantly in PPRV-infected EEC cells. Although MGCD0103 has an excellent inhibitory effect on PPRV multiplication, its inhibitory effect in the virus entry is limited. MGCD0103 significantly reduced the transcription and expression levels of PPRV N gene, as well as the viral titer (Plt;0.002). The half-maximal effective concentration (EC50) and selectivity index (SI) of MGCD0103 against PPRV were 0.43 μmol·L-1 and 4.35, respectively. These results indicated that MGCD0103 significantly inhibited PPRV replication, which is of great significance for the development of anti-PPRV drugs and provides a new idea for constructing sensitive cell lines for PPRV proliferation.
Key words: histone deacetylase inhibitor; MGCD0103; inhibit; peste des petits ruminants virus; replication
*Corresponding authors:" ZENG Qiaoying, E-mail: zengqy@gsau.edu.cn; MENG Xuelian, E-mail: mengxuelian@caas.cn
小反芻獸疫(peste des petits ruminants,PPR)是由小反芻獸疫病毒(peste des petits ruminants virus,PPRV)感染山羊、綿羊等家養和野生小反芻動物引起的一種急性、接觸性、烈性傳染病,被世界動物衛生組織(WOAH)認定為必須申報的疫病,在我國被列為一類動物傳染病。PPR主要流行于以農牧業為主的區域,這些區域經濟發展較為落后、單一,抗風險能力較差,PPR的流行嚴重影響了當地經濟穩定 (DIALLO, 2006)。此外,PPR的感染物種和流行范圍仍在擴大,野生反芻動物感染事件也時有發生。目前PPR的防控主要依靠減毒活疫苗免疫接種,疫苗接種對PPRV的暴發或大流行起很好的控制作用,但仍無可用的特效藥。有研究表明,一些植物提取物或者小分子化合物具有抗PPRV作用,如:金合歡或長葉蓼提取物和表沒食子兒茶素沒食子酸酯(ECGG)[1-3]、銀納米顆粒(silver nanoparticles)[4]、化合物 4,4′-(arylmethylene)bis(1H-pyrazol-5-ols)[5];霉酚酸(MPA)[6]、布喹那(brequinar,BQR)、來氟米特(leflunomide,LFM)、6-氮雜脲嘧啶(6-azauracil,6-AU)[7]等。因此,在PPR疫情防控態勢仍然嚴峻的情況下,開發一些可有效預防或/和治療PPR的藥物具有重要意義。
組蛋白乙酰化酶(HATs)與組蛋白去乙酰化酶(HDACs)是調控蛋白質乙酰化修飾水平的兩種拮抗作用的酶,通過添加或去除蛋白質側鏈賴氨酸殘基上的乙酰基團,改變染色體結構,調節靶基因轉錄。乙酰化修飾不僅參與調控細胞增殖、分化和凋亡等很多細胞過程,還影響免疫信號通路傳導,調節炎癥和免疫反應[8],這使得蛋白質乙酰化位點成了眾多疾病新藥設計的有利靶標。HDACs共分為四類,其中Ⅰ類(HDAC1、HDAC2、HDAC3和HDAC8)、Ⅱ類(HDAC4、HDAC5、HDAC6、HDAC7、HDAC9和HDAC10)、IV類(HDAC11)都屬于Zn2+依賴酶,而Ⅲ類(SirT1-7)屬于NAD+依賴酶。HDACs在控制病毒感染中扮演著關鍵角色,其抑制劑(HDACi)在調節病原體感染方面的研究也屢見不鮮。HDACi可抑制新冠病毒(SARS-CoV-2)RNA轉錄[9-10]、減少皰疹病毒(HSV)基因組的表達[11-13]、可抑制日本腦炎病毒(JEV)[14]和呼吸道合胞體病毒(RSV)[15]的復制,應用HDACi激活潛伏期病毒,聯合其他藥物,以達到治愈持續性病毒感染疾病的目的[16-18]。MGCD0103是一種化學合成的選擇性小分子HDACi,對Ⅰ類和Ⅳ類HDAC具有高度特異性,對HDAC1抑制作用最強。研究表明,MGCD0103可通過線粒體途徑、自噬作用、與蛋白酶抑制劑協同作用或破壞微管的穩定誘導細胞死亡[19-22]。此外,MGCD0103可通過下調SARS-CoV-2的主要受體ACE2以及宿主蛋白-病原體蛋白相互作用網絡(PPI)的其他重要蛋白抑制病毒復制[23]。盡管越來越多的證據表明HDACi可影響多種病原的侵染過程,但關于MGCD0103能否影響PPRV在易感宿主細胞增殖的研究還未見報道。本研究將以山羊子宮內膜上皮細胞(EEC細胞)作為靶細胞,從轉錄水平、蛋白水平以及病毒效價來評價MGCD0103對PPRV復制的作用及抗病毒活性,為明確HDAC在PPRV感染中的作用和研發抗PPRV藥物提供理論依據。
1 材料與方法
1.1 材料
1.1.1 病毒株和細胞系
山羊子宮內膜上皮細胞(EEC細胞)由本實驗室根據文獻[24]制備,小反芻獸疫病毒疫苗株Nigeria 75/1由中國獸醫藥品監察所支海兵研究員饋贈。
1.1.2 主要試劑
小鼠抗PPRV核衣殼蛋白(N蛋白)單克隆抗體由本實驗室制備;小鼠抗β-Actin單克隆抗體、FITC標記山羊抗小鼠IgG、HRP標記山羊抗小鼠IgG均購自Abcam公司(香港);總RNA提取試劑Trizol購自生工生物工程(上海)股份有限公司;StarScript Ⅲ去基因組DNA反轉錄預混試劑和2×RealStar Fast染料法qPCR預混液購自康潤誠業生物科技有限公司(北京);CCK-8 試劑盒購自白鯊生命科學(安徽);Western及IP細胞裂解液購自碧云天生物技術(上海);BCA法蛋白質濃度測定試劑盒購自生工生物工程技術服務有限公司(上海);SDS-PAGE蛋白上樣緩沖液(5X)購自博士德生物(武漢);SuperKineTM增強型ECL發光液(皮克級)購自Abbkine Scientific Co., Ltd(武漢)。
1.1.3 引物合成
利用oligo 6.31引物設計軟件,分別根據PPRV N基因序列和山羊的相關基因序列設計熒光定量PCR引物,所有引物均由上海生工生物工程技術服務有限公司合成(表1)。
1.2 方法
1.2.1 細胞培養和病毒增殖
從液氮中取出凍存的細胞,置于37 ℃水浴鍋中快速解凍,離心棄掉上清,加入5 mL含10%胎牛血清(FBS)的DMEM/F12培養液重懸,轉移至T25細胞培養瓶,置于37 ℃、5% CO2恒溫箱培養,當細胞豐度達90%時進行傳代。
細胞豐度達到80%接種PPRV(MOI=0.01),置于37 ℃、5% CO2恒溫培養箱孵育2 h,而后棄掉含PPRV的培養液,用PBS洗滌,重新加入含2% FBS的DMEM/F12培養液培養。每隔12 h觀察細胞病變(CPE)情況,當CPE達80%左右,收集病毒培養液,分裝后于-80 ℃保存待用。
1.2.2 Time-of-addition (TOA) assay
利用time-of-addition (TOA) assay確定MGCD0103對PPRV復制周期的作用階段。在PPRV感染宿主細胞的不同時間段加入MGCD0103,具體分組如下。
A.“Full-time”處理組:用MGCD0103預處理細胞4 h,接種PPRV(MOI=0.01),換用新的含MGCD0103和2% FBS的DMEM/F12培養液培養3 d。
B.“Entry”處理組:用MGCD0103預處理細胞4 h,接種PPRV(MOI=0.01),換用含2% FBS的DMEM/F12培養液培養3 d。
C.“Post-entry”處理組:PPRV(MOI=0.01)接種細胞,換用含MGCD0103和2% FBS的DMEM/F12培養液培養3 d。收集病毒培養物,分別檢測N基因轉錄水平、N蛋白表達量和病毒滴度。
1.2.3 熒光定量PCR檢測
收集病毒感染或者藥物處理的細胞樣品,提取總RNA,參照StarScript Ⅲ去基因組 DNA反轉錄預混試劑操作說明書將RNA反轉錄成cDNA;以cDNA為模板,利用2×RealStar Fast SYBR qPCR Mix分別擴增目的基因,以ACTB作為內參基因,使用2-ΔΔCt計算目的基因相對表達量。
1.2.4 免疫印記(Western blot)分析
利用細胞裂解液處理病毒感染或者藥物處理的細胞樣品,測定蛋白濃度,SDS-PAGE電泳后轉印至PVDF膜,3% BSA室溫封閉2 h,TBST清洗后置于抗PPRV-N蛋白抗體(1∶1 000)或者抗ACTB(1∶5 000)抗體中4℃孵育過夜;同上洗滌后將PVDF膜置于山羊抗小鼠抗體(1∶5 000)中室溫孵育1 h;洗滌,ECL發光液顯色,Gel Doc XR+蛋白凝膠成像分析儀分析。
1.2.5 病毒滴度檢測
收集病毒培養物,凍融處理,10倍梯度稀釋后待檢。將Vero細胞鋪至96孔板,每孔3 000個細胞,培養24 h后棄掉上清,每孔加入50 μL稀釋好的病毒液和150 μL含2% FBS的DMEM/F12,逐日觀察并記錄細胞病變情況,至連續3 d無新培養孔細胞出現CPE,終止試驗,使用Reed-Muench法統計病毒滴度(TCID50·mL-1)。
1.2.6 抗病毒活性評價
1.2.6.1 細胞半數毒性濃度(CC50)測定:用含10% FBS的DMEM/F12培養基稀釋細胞后鋪至96孔細胞培養板,每孔100 μL(含1×104個細胞),37 ℃、5%濃度CO2恒溫培養箱培養。當細胞豐度達70%時,試驗組每孔加入1 μL DMSO稀釋的不同濃度MGCD0103,對照組每孔加入1 μL DMSO,5% CO2、37 ℃培養。3 d后每孔加入10 μL CCK-8試劑,孵育1 h后用酶標儀測定450 nm吸光度,計算出各試驗組細胞活力。以藥物濃度為橫坐標,細胞活力為縱坐標,利用GraphPad 8.0軟件進行統計分析,計算細胞半數毒性濃度(CC50)。
1.2.6.2 半數有效濃度(EC50)測定:
利巴韋林(ribavirin)作為一種廣譜性抗病毒藥物,能夠抑制PPRV的感染[6,25],選擇其為陽性對照藥物。當細胞豐度達70%,分別用0.125、0.25、0.5、1和2 μmol·L-1 MGCD0103處理,4 h后試驗組換用含PPRV的培養液孵育,2 h后換為含不同濃度MGCD0103的新培養液,同時設置細胞對照組、未用藥物處理的PPRV對照組和濃度分別為64、128、256、512、1 024和2 048 μmol·L-1的陽性藥物利巴韋林對照組,每個濃度設置3個重復。于37 ℃、5% CO2恒溫培養箱培養3 d,收集病毒培養物,提取RNA,用RT-qPCR檢測PPRV N基因轉錄水平。根據PPRV N基因相對轉錄水平計算藥物對PPRV抑制率,以藥物濃度為橫坐標,細胞病變抑制率為縱坐標,利用Graphpad 8.0軟件繪制細胞病變抑制率的劑量依賴曲線,進行回歸分析,計算藥物半數有效濃度(EC50)。
根據細胞半數毒性濃度(CC50)和抑制細胞產毒半數有效濃度(EC50)計算藥物選擇指數(SI)。
1.2.7 數據統計分析
所有試驗均進行三次獨立重復,利用Graphpad Prism 8.0進行單因素方差分析,組間差異用t檢驗分析,試驗結果用“x-±s”表示,Alpha 為 0.05,Plt;0.033 (*)、Plt;0.002 (**) 和Plt;0.001 (***)被認為統計具有不同的顯著性。
2 結 果
2.1 PPRV感染對宿主細胞組蛋白去乙酰化酶轉錄的影響
為了明確宿主細胞組蛋白去乙酰化酶(HDACs)是否影響PPRV感染過程,作者收集了PPRV感染3 d后的宿主細胞,提取總RNA,通過SYBR Green實時熒光定量RT-PCR檢測HDACs的相對轉錄水平,其中以未感染PPRV的宿主細胞組作為對照。分析qPCR數據可知,PPRV明顯提高了宿主細胞HDAC1(Plt;0.001)和HDAC2的轉錄水平(Plt;0.002)(圖1)。
2.2 MGCD0103對EEC細胞毒性作用檢測
為了確定MGCD0103在EEC細胞的最高工作濃度,使用CCK-8試劑盒對不同濃度MGCD0103對EEC細胞的毒性作用進行了檢測。結果表明:MGCD0103濃度等于2 μmol·L-1時,細胞活力為44.5%;MGCD0103濃度等于1 μmol·L-1時,細胞活力為65.9%;MGCD0103濃度小于0.5 μmol·L-1時,EEC細胞活力大于95%(圖2)。因此,選擇0.5 μmol·L-1作為MGCD0103在EEC細胞中的最高工作濃度。
2.3 MGCD0103對PPRV感染過程的影響
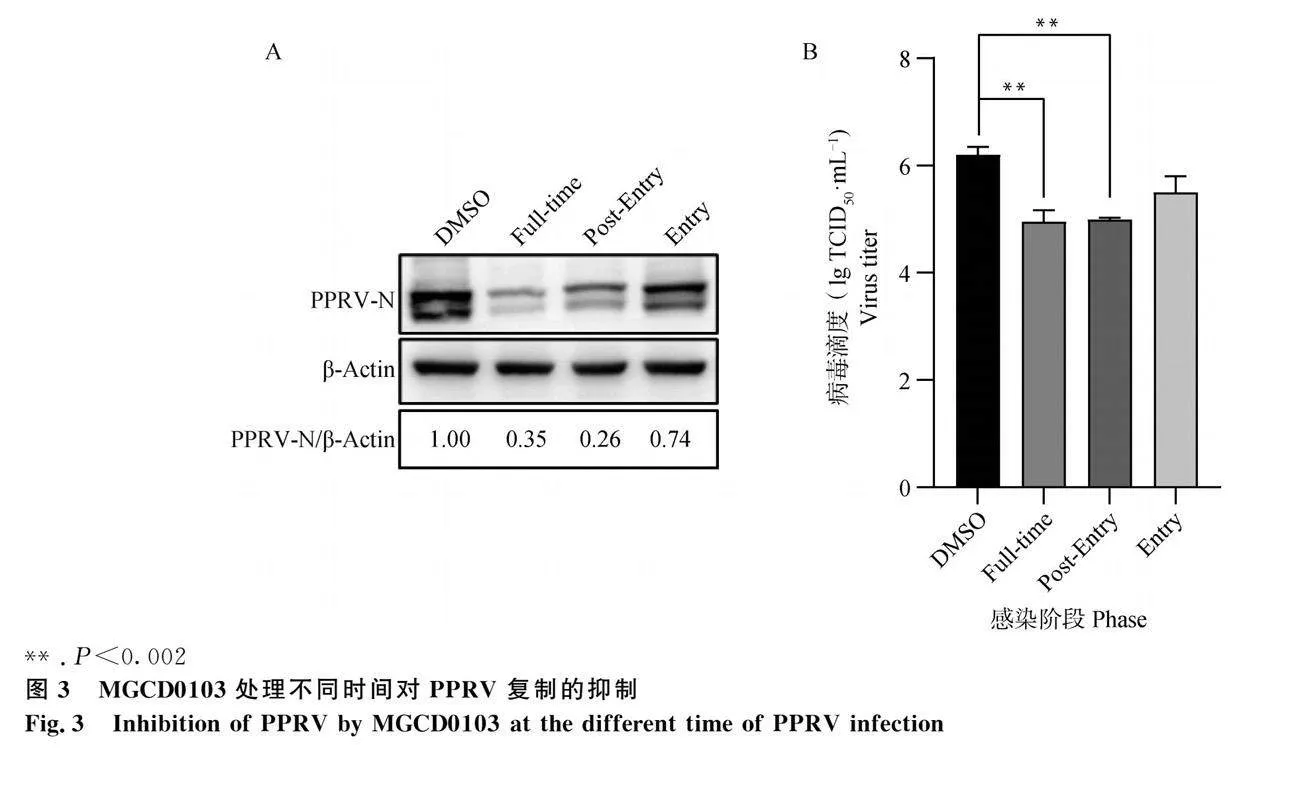
使用0.5 μmol·L-1 MGCD0103進行TOA試驗,以確定MGCD0103影響PPRV侵染過程的具體階段。TOA結果表明,MGCD0103對PPRV復制具有明顯的抑制作用,其中對病毒吸附和入侵環節的抑制作用較弱,對進入細胞后病毒復制的抑制作用較強(圖3)。
2.4 MGCD0103對PPRV N基因轉錄具有抑制作用
用0.125、0.25、0.5 μmol·L-1MGCD0103處理細胞并感染PPRV,培養3 d后收集樣品,提取RNA,使用SYBR Green實時定量PCR檢測PPRV N基因mRNA相對轉錄水平。通過分析可知,當MGCD0103濃度大于0.5 μmol·L-1,MGCD0103處理組PPRV N基因相對轉錄水平顯著低于對照組(DMSO),差異顯著(0.25 μmol·L-1,Plt;0.002; 0.5μmol·L-1,Plt;0.001)(圖4A)。同時,用0.5 μmol·L-1 MGCD0103處理細胞并感染PPRV培養不同時間(1、2、3、4和5 d)的結果表明,感染3 d后藥物處理組PPRV N基因相對轉錄水平與對照組差異顯著(3 d,Plt;0.001;4 d和5 d,Plt;0.002)(圖4B)。
2.5 MGCD0103對PPRV N蛋白表達具有抑制作用
為了進一步確認MGCD0103對PPRV在EEC細胞中復制的作用,用0.125、0.25、0.5 μmol·L-1 MGCD0103處理細胞并感染PPRV,培養3 d后收集樣品,進行Western blot檢測。結果表明,藥物處理組PPRV N蛋白表達量低于對照組,其中 0.50 μmol·L-1 MGCD0103作用引起的差異最大(圖5)。同時,用0.5 μmol·L-1 MGCD0103處理細胞并感染PPRV培養不同時間(1、2、3、4和5 d)的結果表明,藥物處理組PPRV N蛋白表達量低于對照組,感染病毒3 d時的差異最大(圖6)。
2.6 MGCD0103降低了PPRV的病毒滴度
病毒滴度是藥物抗病毒作用最重要的評判指標。本試驗用0.125、0.25、0.5 μmol·L-1 MGCD0103處理細胞并感染PPRV,培養5 d后收集樣品,測算TCID50。結果表明,MGCD0103濃度大于0.25 μmol·L-1組的病毒滴度明顯低于對照組,差異顯著(0.25 μmol·L-1,Plt;0.002; 0.5 μmol·L-1,Plt;0.001)(圖7)。
2.7 MGCD0103抗PPRV活性
利巴韋林可以抑制DNA或RNA的合成,從而抑制病毒復制。因此,以利巴韋林作為抑制PPRV復制的陽性對照藥,通過添加不同劑量的MGCD0103,確定MGCD0103對50%宿主細胞產生毒性的濃度(CC50)和抑制50% PPRV復制的藥物半數有效濃度(EC50)。結果表明,MGCD0103對EEC細胞的CC50為1.87 μmol·L-1,而利巴韋林對EEC細胞的CC50大于2047.4 μmol·L-1;MGCD0103抑制PPRV復制的EC50為0.43 μmol·L-1,而利巴韋林對PPRV的EC50為450.4 μmol·L-1;計算得到MGCD0103在EEC細胞中抑制PPRV復制的藥物選擇常數為4.35,而利巴韋林在EEC細胞中抑制PPRV復制的藥物選擇常數為gt;4.55(圖8)。
3 討 論
HDACi主要被作為抗腫瘤藥物進行廣泛使用。隨著對HDACi作用研究的不斷深入和擴展,發現其在病毒復制方面也表現出重要作用。一些HDACi可抑制病毒復制,如:Vorinostat(SAHA)抑制人巨細胞病毒(HCMV)[26],SAHA、CI994I、 RGFP966可抑制HCV病毒復制[27-28],Tubacin可減弱JEV的復制[14],Trichostatin A(TSA)/SAHA可顯著抑制RSV復制[15]。但值得關注的是,這些小分子化合物也可通過抑制HDAC的表達而促進病毒復制,如:SAHA、TSA、MGCD0103和PCI34051顯著增加口蹄疫病毒(FMDV)復制[29]。此外,HDACi還表現出清除病毒感染的作用[30-31]。作為一種選擇性抑制劑,MGCD0103主要抑制HDAC1、2、3和11,其中對HDAC1抑制作用最強。Xu等[32]研究證明HDAC1顯著抑制豬流行性腹瀉病毒(PEDV)復制,同時影響ISG15和OAS等多種抗病毒基因的表達。本研究表明,PPRV感染可引起宿主細胞HDAC1和HDAC2轉錄水平明顯上升,而用MGCD0103處理宿主細胞可以抑制PPRV N基因轉錄、表達且降低了病毒滴度,這提示MGCD0103對PPRV復制的抑制效果可能是其對HDAC1、2的抑制作用所致。PPRV和RSV同屬于副黏病毒科,Feng等[15]研究表明,RSV感染可促進呼吸道上皮細胞(ACEs)中HDAC2的表達,降低了組蛋白乙酰化;用TSA和SAHA處理ACEs,可通過上調IFN-α相關通路顯著抑制RSV復制;用TSA和SAHA處理RSV感染的ACEs能明顯抑制RSV誘導的促炎性細胞因子(IL-6和IL-8)和氧化應激相關分子(MDA和NO)的釋放,并且顯著抑制調控促炎基因表達和氧化應激損傷的NF-κB、COX-2、MAPK和Stat3的活化,從而減少RSV誘導的呼吸道炎癥和氧化應激。這與本研究的結果一致。Icardi等[33]研究表明HDAC1和HDAC2對Ⅰ型干擾素誘導的STAT3和ISGF3轉錄活性的調控存在相反作用,這表明HDAC1和HDAC2參與調控宿主細胞天然免疫應答。本研究表明,MGCD0103對PPRV的抑制作用貫穿病毒的整個生命周期,但對病毒進入細胞后復制過程的作用最強。作者猜測:PPRV感染宿主細胞后,激活天然免疫從而抑制病毒復制,而MGCD0103可能對病毒激活宿主細胞天然免疫起到協同作用,這需要進一步深入研究HDACs對PPRV復制機制的調控機制。
目前研究者主要利用Vero細胞或一些表達PPRV受體的細胞系研究PPRV的致病機制,借助易感宿主源細胞的研究較少[34-36]。PPRV的自然宿主主要是山羊、綿羊等小型反芻動物,EEC細胞是山羊子宮內膜上皮細胞,它能支持PPRV的增殖,并具有明顯的細胞病理作用。此外,Tang等[24]研究發現,PPRV能有效誘導EEC細胞天然免疫反應,EEC細胞是研究PPRV的一個合適的細胞模型,可用于病毒感染和先天免疫機制的研究。因此,本研究選擇山羊EEC細胞系比較MGCD0103處理后PPRV增殖變化,這更能反映MGCD0103對PPRV的實際作用。
雖然PPRV的防控主要依賴于疫苗接種,但也有研究表明一些藥物可以抑制PPRV的增殖[1-7,37-39],這些藥物主要通過抑制病毒進入[2-4]、影響病毒RNA合成[6-7,37,39]或者調控天然免疫通路[37]抑制PPRV復制。隨著對HDACi研究的不斷深入,發現HDACi通過競爭性抑制HDAC活性,從而提高組蛋白乙酰化水平、改變特定基因的轉錄與表達、調控免疫通路,可作為一類新的防治病毒的藥物[14-15,26-28]。本試驗研究表明HDACs抑制劑MGCD0103對PPRV的復制具有抑制作用;在EEC細胞中,MGCD0103抑制PPRV復制的EC50為0.43 μmol·L-1,僅約為陽性對照藥物利巴韋林EC50(450.4 μmol·L-1)的千分之一,且MGCD0103的SI值(4.35)近似于利巴韋林的SI值(gt;4.55)。因此,以上結果提示MGCD0103是一種潛在的PPRV抑制劑,HDAC1和HDAC2可以作為新的藥物靶點用于抗PPRV藥物研發和育種。
4 結 論
PPRV感染可引起宿主HDAC1和HDAC2轉錄水平明顯上升,抑制HDAC1、HDAC2的選擇性抑制劑MGCD0103可抑制小反芻獸疫病毒N基因轉錄、表達和病毒復制,其EC50僅約為陽性對照藥物利巴韋林的千分之一,SI值近似于利巴韋林;作為一種抗腫瘤藥物,MGCD0103已用于臨床Phase 2研究階段;綜上所述,MGCD0103可顯著抑制PPRV的復制,可作為一種抗PPRV的候選藥物。
參考文獻(References):
[1] RAHEEL R, ASHRAF M, EJAZ S, et al. Assessment of the cytotoxic and anti-viral potential of aqueous extracts from different parts of Acacia nilotica (Linn) Delile against Peste des petits ruminants virus[J]. Environ Toxicol Pharmacol, 2013, 35(1):72-81.
[2] YADAV P, CHOUDHURY S, BARUA S, et al. Polyalthia longifolia leaves methanolic extract targets entry and budding of viruses-an in vitro experimental study against paramyxoviruses[J]. J Ethnopharmacol, 2020, 248:112279.
[3] SAADH M. Epigallocatechin gallate (EGCG) combined with zinc sulfate inhibits Peste des petits ruminants virus entry and replication[J]. Saudi J Biol Sci, 2021, 28(11):6674-6678.
[4] KHANDELWAL N, KAUR G, CHAUBEY K K, et al. Silver nanoparticles impair Peste des petits ruminants virus replication[J]. Virus Res, 2014, 190:1-7.
[5] SUJATHA K, SHANTHI G, SELVAM N P, et al. Synthesis and antiviral activity of 4, 4′-(arylmethylene)bis(1H-pyrazol-5-ols) against peste des petits ruminant virus (PPRV)[J]. Bioorg Med Chem Lett, 2009, 19(15):4501-4503.
[6] CHANG Q Y, GUO F C, LI X R, et al. The IMPDH inhibitors, ribavirin and mycophenolic acid, inhibit peste des petits ruminants virus infection[J]. Vet Res Commun, 2018, 42(4):309-313.
[7] JIN L, LI Y C, PU F Y, et al. Inhibiting pyrimidine biosynthesis impairs Peste des Petits Ruminants Virus replication through depletion of nucleoside pools and activation of cellular immunity[J]. Vet Microbiol, 2021, 260:109186.
[8] GUISE A J, BUDAYEVA H G, DINER B A, et al. Histone deacetylases in herpesvirus replication and virus-stimulated host defense[J]. Viruses, 2013, 5(7):1607-1632.
[9] PHIEL C J, ZHANG F, HUANG E Y, et al. Histone deacetylase is a direct target of valproic acid, a potent anticonvulsant, mood stabilizer, and teratogen[J]. J Biol Chem, 2001, 276(39):36734-36741.
[10] UNAL G, TURAN B, BALCIOGLU Y H. Immunopharmacological management of COVID-19:Potential therapeutic role of valproic acid[J]. Med Hypotheses, 2020, 143:109891.
[11] JONES K, NOURSE J, CORBETT G, et al. Sodium valproate in combination with ganciclovir induces lysis of EBV-infected lymphoma cells without impairing EBV-specific T-cell immunity[J]. Int J Lab Hematol, 2010, 32(1 Pt 1):e169-e174.
[12] GHOSH S K, PERRINE S P, WILLIAMS R M, et al. Histone deacetylase inhibitors are potent inducers of gene expression in latent EBV and sensitize lymphoma cells to nucleoside antiviral agents[J]. Blood, 2012, 119(4):1008-1017.
[13] SHAPIRA L, RALPH M, TOMER E, et al. Histone deacetylase inhibitors reduce the number of herpes simplex virus-1 genomes initiating expression in individual cells[J]. Front Microbiol, 2016, 7:1970.
[14] LU C Y, CHANG Y C, HUA C H, et al. Tubacin, an HDAC6 selective inhibitor, reduces the replication of the japanese encephalitis virus via the decrease of viral RNA synthesis[J]. Int J Mol Sci, 2017, 18(5):954.
[15] FENG Q Q, SU Z L, SONG S Y, et al. Histone deacetylase inhibitors suppress RSV infection and alleviate virus-induced airway inflammation[J]. Int J Mol Med, 2016, 38(3):812-822.
[16] WIGHTMAN F, ELLENBERG P, CHURCHILL M, et al. HDAC inhibitors in HIV[J]. Immunol Cell Biol, 2012, 90(1):47-54.
[17] LI Y X, SETO E. HDACs and HDAC inhibitors in cancer development and therapy[J]. Cold Spring Harb Perspect Med, 2016, 6(10):a026831.
[18] 潘春容, 鄧瑞雪, 胡林杰, 等. 組蛋白去乙酰化酶抑制劑對小反芻獸疫病毒復制的影響[J]. 畜牧獸醫學報, 2022, 53(7): 2307-2316.
PAN C R, DENG R X, HU L J, et al. Effect of histone deacetylase inhibitors on Peste Des Petits ruminants virus replication[J]. Acta Veterinaria et Zootechnica Sinica, 2022, 53(7): 2307-2316.(in Chinese)
[19] BUGLIO D, MAMIDIPUDI V, KHASKHELY N M, et al. The class-I HDAC inhibitor MGCD0103 induces apoptosis in Hodgkin lymphoma cell lines and synergizes with proteasome inhibitors by an HDAC6-independent mechanism[J]. Br J Haematol, 2010, 151(4):387-396.
[20] EL-KHOURY V, MOUSSAY E, JANJI B, et al. The histone deacetylase inhibitor MGCD0103 induces apoptosis in B-cell chronic lymphocytic leukemia cells through a mitochondria-mediated caspase activation cascade[J]. Mol Cancer Ther, 2010, 9(5):1349-1360.
[21] WEI Y, KADIA T, TONG W G, et al. The combination of a histone deacetylase inhibitor with the Bcl-2 homology domain-3 mimetic GX15-070 has synergistic antileukemia activity by activating both apoptosis and autophagy[J]. Clin Cancer Res, 2010, 16(15):3923-3932.
[22] CHIA K M, BEAMISH H, JAFFERI K, et al. The histone deacetylase inhibitor MGCD0103 has both deacetylase and microtubule inhibitory activity[J]. Mol Pharmacol, 2010, 78(3):436-443.
[23] CHAKRABARTY B, DAS D, BULUSU G, et al. Network-based analysis of fatal comorbidities of COVID-19 and potential therapeutics[J]. IEEE/ACM Trans Comput Biol Bioinform, 2021, 18(4):1271-1280.
[24] TANG J Y, TANG A X, DU H Y, et al. Peste des petits ruminants virus exhibits cell-dependent interferon active response[J]. Front Cell Infect Microbiol, 2022, 12:874936.
[25] 王丹陽, 張 康, 王旭榮, 等. 訶子、矮紫堇、甘青烏頭提取物對牛病毒性腹瀉病毒的體外抑制作用[J]. 畜牧獸醫學報, 2018, 49(9): 2036-2043.
WANG D Y, ZHANG K, WANG X R, et al. Inhibitory effects of extracts from Terminalia chebula, Corydalis hendersonii, Aconitum tanguticum on bovine viral diarrhea virus in vitro[J]. Acta Veterinaria et Zootechnica Sinica, 2018, 49(9): 2036-2043. (in Chinese)
[26] LIU Z S, XUAN B Q, TANG S B, et al. Histone deacetylase inhibitor SAHA induces expression of fatty acid-binding protein 4 and inhibits replication of human cytomegalovirus[J]. Virol Sin, 2021, 36(6):1352-1362.
[27] ZHOU Y, WANG Q, YANG Q, et al. Histone deacetylase 3 inhibitor suppresses hepatitis c virus replication by regulating apo-A1 and LEAP-1 expression[J]. Virol Sin, 2018, 33(5):418-428.
[28] SATO A, SAITO Y, SUGIYAMA K, et al. Suppressive effect of the histone deacetylase inhibitor suberoylanilide hydroxamic acid (SAHA) on hepatitis C virus replication[J]. J Cell Biochem, 2013, 114(9):1987-1996.
[29] ZHANG H J, WANG X W, QU M, et al. Foot-and-mouth disease virus structural protein VP3 interacts with HDAC8 and promotes its autophagic degradation to facilitate viral replication[J]. Autophagy, 2023, 19(11):2869-2883.
[30] KOBAYASHI Y, GéLINAS C, DOUGHERTY J P. Histone deacetylase inhibitors containing a benzamide functional group and a pyridyl cap are preferentially effective human immunodeficiency virus-1 latency-reversing agents in primary resting CD4+ T cells[J]. J Gen Virol, 2017, 98(4):799-809.
[31] ARCHIN N M, LIBERTY A L, KASHUBA A D, et al. Administration of vorinostat disrupts HIV-1 latency in patients on antiretroviral therapy[J]. Nature, 2012, 487(7408):482-485.
[32] XU J D, MAO J Y, HAN X, et al. Porcine epidemic diarrhea virus inhibits HDAC1 expression to facilitate its replication via binding of its nucleocapsid protein to host transcription factor Sp1[J]. J Virol, 2021, 95(18):e0085321.
[33] ICARDI L, LIEVENS S, MORI R, et al. Opposed regulation of type I IFN-induced STAT3 and ISGF3 transcriptional activities by histone deacetylases (HDACS) 1 and 2[J]. FASEB J, 2012, 26(1):240-249.
[34] BEGUM S, NOORUZZAMAN M, HASNAT A, et al. Isolation of peste des petits ruminants virus using primary goat kidney cell culture from kidneys obtained at slaughter[J]. Vet Med Sci, 2021, 7(3):915-922.
[35] CLARKE B D, ISLAM M R, YUSUF M A, et al. Molecular detection, isolation and characterization of Peste-des-petits ruminants virus from goat milk from outbreaks in Bangladesh and its implication for eradication strategy[J]. Transbound Emerg Dis, 2018, 65(6):1597-1604.
[36] KUMAR N, CHAUBEY K K, CHAUDHARY K, et al. Isolation, identification and characterization of a Peste des Petits Ruminants virus from an outbreak in Nanakpur, India[J]. J Virol Methods, 2013, 189(2):388-392.
[37] EL-SABBAGH O I, EL-SADEK M E, EL-KALYOUBI S, et al. Synthesis, DNA binding and antiviral activity of new uracil, xanthine, and pteridine derivatives[J]. Arch Pharm (Weinheim), 2007, 340(1):26-31.
[38] ZHANG W F, DENG H L, LIU Y F, et al. Antiviral effectivity of favipiravir against peste des petits ruminants virus is mediated by the JAK/STAT and PI3K/AKT pathways[J]. Front Vet Sci, 2021, 8:722840.
[39] KHANDELWAL N, CHANDER Y, RAWAT K D, et al. Emetine inhibits replication of RNA and DNA viruses without generating drug-resistant virus variants[J]. Antiviral Res, 2017, 144:196-204.
(編輯 白永平)
