間性豬垂體的全轉錄組學特征分析
2024-12-18 00:00:00于聰穎吳金華鐘秉洲趙海全譚淑雯于輝李華
畜牧獸醫學報 2024年11期

摘 要: 旨在探索間性豬垂體的編碼與非編碼RNAs的表達特征,為解析間性豬垂體功能紊亂的分子機制提供數據支持。以5月齡正常母豬和間性豬各3頭為研究對象,進行血清激素檢測和垂體組織的全轉錄組測序,分析鑒定間性豬垂體差異表達的mRNAs、lncRNAs、miRNAs,并構建間性豬垂體中相關基因調控的競爭性內源RNAs(ceRNAs)。結果表明,間性豬血清激素分泌紊亂,垂體功能異常。與正常母豬對比,間性豬垂體差異表達的mRNAs有1 451個,差異表達的lncRNAs有277個,差異表達的miRNAs有17個。其中差異表達mRNAs主要富集在MAPK信號通路、孕酮介導的卵母細胞成熟、PRL信號通路等生物通路上;ceRNAs網絡分析發現,TCONS_00175477-novel_265-CCNB3、TCONS_00134726-novel_265-ZNF366和TCONS_00212783-novel_265-ZNF366競爭組合可能與間性豬垂體激素分泌異常有關。綜上所述,本研究揭示了間性豬垂體mRNAs、lncRNAs和miRNAs差異表達并構建ceRNAs,其特定lncRNA-miRNA-mRNA可能參與間性豬垂體的激素合成與分泌的調控,為解析間性豬垂體功能紊亂的分子機制提供了理論參考。
關鍵詞: 間性豬;垂體;全轉錄組;ceRNA;激素
中圖分類號:S828.2
文獻標志碼:A
文章編號:0366-6964(2024)11-4925-13
收稿日期:2024-04-28
基金項目:國家自然科學基金(32072699);廣東省粵佛聯合基金重點項目(2020B1515120057)
作者簡介:于聰穎(2001-),女,吉林靖宇人,碩士生,主要從事動物遺傳育種與繁殖研究,E-mail:13415551629@163.com
*通信作者:于 輝,主要從事畜牧生產研究,E-mail: yu71hui@aliyun.com;李 華,主要從事動物遺傳育種研究,E-mail: okhuali@fosu.edu.cn
Analysis of Whole Transcriptome Characteristics of the Hermaphroditic Pig’s Pituitary
Gland
YU" Congying, WU" Jinhua, ZHONG" Bingzhou, ZHAO" Haiquan, TAN" Shuwen, YU" Hui*, LI" Hua*
(Guangdong Provincial Key Laboratory of Animal Molecular Design and Precision Breeding, Foshan University,
Foshan 528225," China)
Abstract:" The aim of this study was to explore the expression characteristics of coding and non-coding RNAs in the pituitary gland of hermaphroditic pigs, thereby providing supporting data for the analysis of the molecular mechanisms underlying pituitary dysfunction in these pigs. Three five-month-old normal sows and three hermaphroditic pigs were used for the study. Serum hormone assays and whole transcriptome sequencing of pituitary tissues were carried out to analyze and identify differentially expressed mRNAs, lncRNAs, and miRNAs in the pituitary of hermaphroditic pigs. Additionally, a competing endogenous RNA (ceRNA) network was established to elucidate the regulatory relationships of related genes in the pituitary of hermaphroditic pigs. Disrupted serum hormone secretion and abnormal pituitary function were observed in hermaphroditic pigs. Comparison with normal sows, 1 451 differentially expressed mRNAs, 277 differentially expressed lncRNAs, and 17 differentially expressed miRNAs were identified in the pituitary gland of hermaphroditic pigs, which were enriched in biological pathways, such as the MAPK signaling pathway, the progesterone-mediated oocyte maturation, and the PRL signaling pathway. Through the analysis of the ceRNA network, competitive combinations of TCONS_00175477-novel_265-CCNB3, TCONS_00134726-novel_265-ZNF366 and TCONS_00212783-novel_265-ZNF366 were identified possibly related to abnormal pituitary hormone secretion in hermaphroditic pigs. In summary, the differentially expressed mRNAs, lncRNAs and miRNAs in the pituitary gland of hermaphroditic pigs were revealed, thereby ceRNA networks were constructed, which may be involved in regulating hormone synthesis and secretion in this study. All these were aid to support the elucidation of the molecular mechanisms underlying the dysfunction of the pituitary gland in hermaphroditic pigs.
Key words: hermaphroditic pigs; pituitary; whole transcriptome; ceRNA; hormones
*Corresponding authors:YU Hui, E-mail: yu71hui@aliyun.com;LI Hua, E-mail: okhuali@fosu.edu.cn
性別發育異常(disorders of sex development,DSD)是一組先天性疾病,其呈現出染色體、性腺或解剖的性別不典型[1]。間性豬又稱為38,XX-DSD(SRY-),擁有正常雌性染色體結構,不含有雄性性別決定基因(sex determining region Y,SRY)[2]。間性豬生殖器官發育畸形,繁殖力低,死亡率高,限制了優質種豬的培育,給畜牧業帶來嚴重的經濟損失[3]。內分泌系統下丘腦-垂體-性腺軸在哺乳動物的生殖發育過程中發揮著重要作用[4],其合成分泌的各種生殖激素介導生殖器官的分化和成熟,其中垂體是該軸中承上啟下的內分泌腺體,其分泌的催乳素(prolactin,PRL)、促卵泡素(follicle-stimulating hormone,FSH)和促黃體素(luteinizing hormone,LH)在生殖發育中發揮著不可缺少的作用[5-6]。激素水平是診斷DSD的一個標準,因為它通常與激素分泌紊亂有關。間性豬的孕酮(progesterone,P)水平顯著高于正常母豬,達到5~90倍[7],睪酮(testosterone,TE)和雌二醇(estradiol,E2)水平均介于正常公、母豬之間,同時間性豬睪丸或卵泡中產生的雄烯酮可影響肉質[8]。課題組前期通過透射電鏡觀察到5月齡間性豬垂體中促性腺激素細胞和催乳素細胞分泌顆粒增多,分泌顆粒電子密度大[9],揭示間性豬垂體的結構功能異常,但具體分子機制尚不明確。
非編碼RNA(ncRNAs)可以影響調節性別決定和性腺發育基因的表達和功能[10],如非編碼RNA PISRT1(PIS regulated transcript number1)是一種抗睪丸因子,可抑制睪丸分化的關鍵調節因子SOX9[11],也有研究發現,lncRNA SM2可作為ceRNA,通過靶向Oar-miR-16b/TGF-β/SMAD2信號通路從而在湖羊垂體前葉調控促性腺激素的分泌[12];在卵巢分化過程中miR-124抑制XX性腺SOX9基因的表達[13],He等[14]研究發現miR-7通過與靶基因RAF1作用,介導GnRH和雌激素對促性腺激素的調節作用。此外,不同的蛋白質編碼RNA(mRNA)和ncRNAs之間的相互作用也在各個生物過程中發揮重要作用,如癌癥進展[15]、卵巢功能[16]和下丘腦-垂體-性腺軸功能[17]。
因此,本研究通過對間性豬和正常母豬的垂體進行全轉錄組測序,鑒定垂體中差異表達的mRNAs、lncRNAs和miRNAs,進一步分析差異表達基因的生物學功能并構建ceRNA網絡,為揭示間性豬垂體功能異常的分子調控機制提供重要基礎,為動物DSD等相關疾病研究提供重要的理論依據。
1 材料與方法
1.1 試驗動物和樣品采集
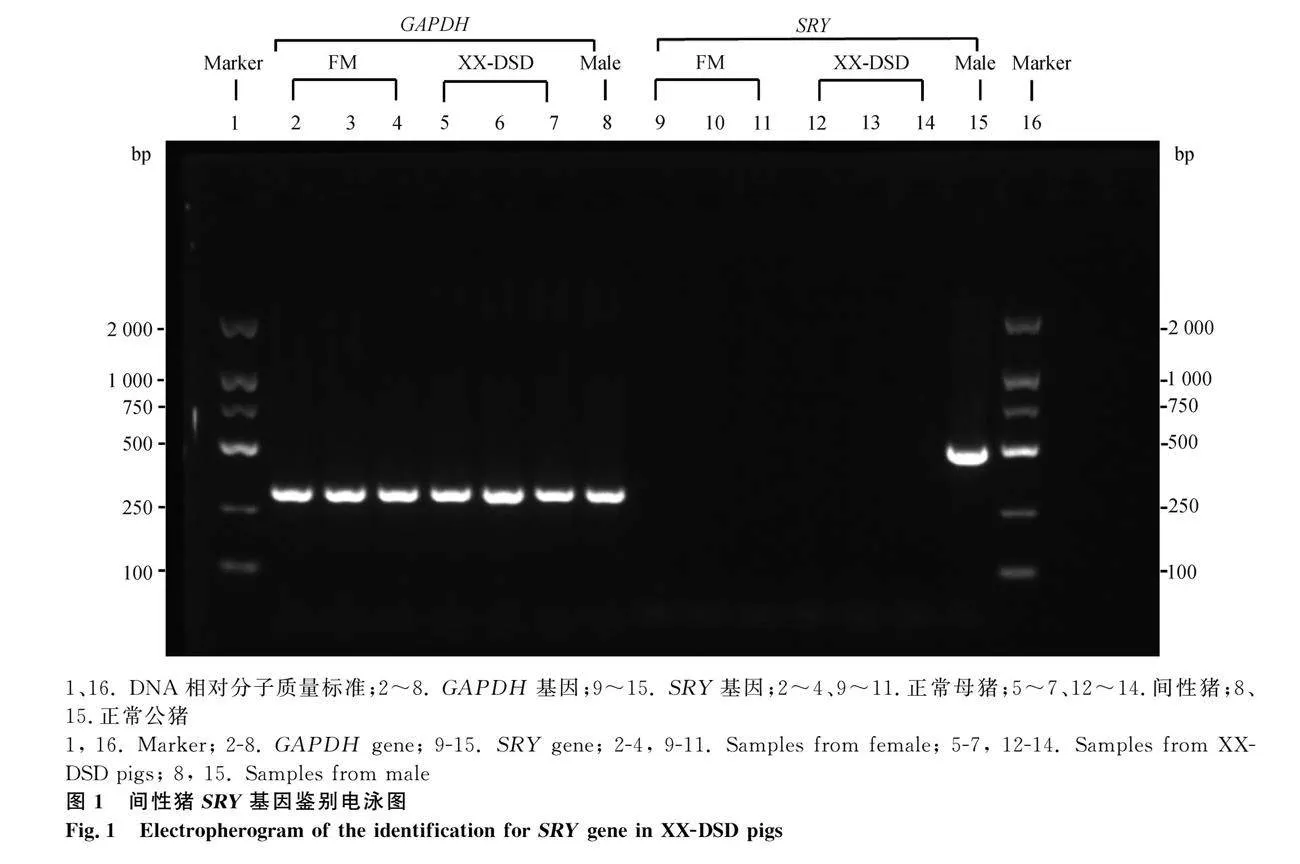
本試驗選取廣東某豬場5月齡約克夏豬的正常母豬3頭(Female,FM1、FM2、FM3)和38, XX-DSD(SRY-)的間性豬3頭(XX-DSD1、XX-DSD2、XX-DSD3)為試驗動物。外生殖器檢查后,通過引物(SRY-F:TGAAAGCGGACGATTACAGC, SRY-R:GGCTTTCTGTTCCTGAGCAC)進行SRY檢測(500 bp)和核型分析。屠宰后采集垂體組織樣品,裝入無RNA酶凍存管中并迅速置于液氮中保存備用。
1.2 激素檢測
取正常母豬以及間性豬血樣并分離血清,利用放射免疫法檢測TE、E2、LH、FSH、P和PRL。
1.3 RNA提取、cDNA文庫構建和Illumina測序
使用TRIzol試劑(Invitrogen,USA)分別提取6個樣品RNA,經測定的RNA提取質量合格,分別用于RNA文庫構建。使用the epicenter Ribo-zero TM rRNA Removal Kit (Epicenter, USA)從每個樣品中去除核糖體RNA(Ribosomal, rRNA)。根據制造商的推薦,使用the NEBNext UltraTM Directional RNA Library Prep Kit for Illumina (NEB, USA)制備cDNA文庫。對構建好的文庫在Illumina HiSeq2500平臺上進行測序,得到raw reads。
1.4 垂體組織差異mRNAs、lncRNAs、miRNAs篩選
對原始數據進行質量控制后得到clean reads,使用HTSeq(v0.6.0)、DESeq2軟件對正常母豬(FM)垂體與間性豬(XX-DSD)垂體的表達基因進行兩兩比較,進行lncRNAs、mRNAs表達定量和差異表達篩選(log2FoldChange(FC)≥1且 P<0.05)。miRDeeP2量化器模塊用于量化已知和新型miRNA的表達和檢索計數。miRNA的表達水平通過百萬轉錄物(TPM)進行表達量歸一處理后,對各樣本進行TPM密度分布統計,使用DESeq2軟件對差異表達miRNAs進行篩選(log2FoldChange(FC)≥1且 P<0.05)。
1.5 差異mRNAs的功能分析
為更好了解XX-DSD垂體中mRNAs的功能,本研究將兩個比較組 (FM vs. XX-DSD)的差異mRNAs分別做了GO(Gene Ontology)、KEGG(Kyoto encyclopedia of genes and genomes)分析,GO與KEGG分析均采用clusterProfiler R包完成,篩選顯著富集結果(P<0.05)。GO富集分為生物過程、細胞組成和分子功能3個部分;KEGG富集選取最顯著的20個KEGG通路繪制柱狀圖進行分析。
1.6 ceRNA互作調控網絡圖的繪制
lncRNA不僅可以通過直接與靶基因結合或者共表達調控靶基因,也可以作為ceRNA與miRNA結合,參與靶基因的表達調控。因此,運用miRanda預測lncRNA和mRNA上的miRNA結合位點,并通過COR篩選出miRNA-lncRNA和miRNA-mRNA對(COR<-0.5, P<0.05),而共表達的lncRNA-mRNA對則以COR>0.99、P<0.05標準篩選。通過分別整合miRNA-lncRNA、miRNA-mRNA和lncRNA-mRNA,獲得的miRNA-mRNA-lncRNA作為一個共競爭表達三聯體(co-expression competing triplet)。運用Cytoscape 3.7.1繪制miRNA-mRNA-lncRNA的網絡。
1.7 實時熒光定量PCR驗證測序結果
為了進一步保證測序結果的可靠性,對隨機挑選的4個mRNAs、4個lncRNAs和4個miRNAs進行qRT-PCR分析。樣品RNA提取與“1.2”相同,采用PrimeScriptTM RT reagent Kit with gDNA Eraser(Takara)試劑盒去除基因組DNA并合成cDNA,按照TB Green Premix Ex TaqTM II試劑盒指示在ABI 7500實時熒光定量PCR儀上進行檢測。驗證測序數據集。相對表達量結果采用2-ΔΔCt法計算得到。mRNAs和lncRNAs定量檢測以GAPDH為內參,miRNAs定量檢測以U6作為內參(表1)。
2 結 果
2.1 間性豬SRY基因的檢測
正常公豬(male)DNA能檢測出SRY基因,而母豬(FM)和間性豬(DSD)未檢測到SRY基因,表明間性豬不含SRY基因(圖1)。
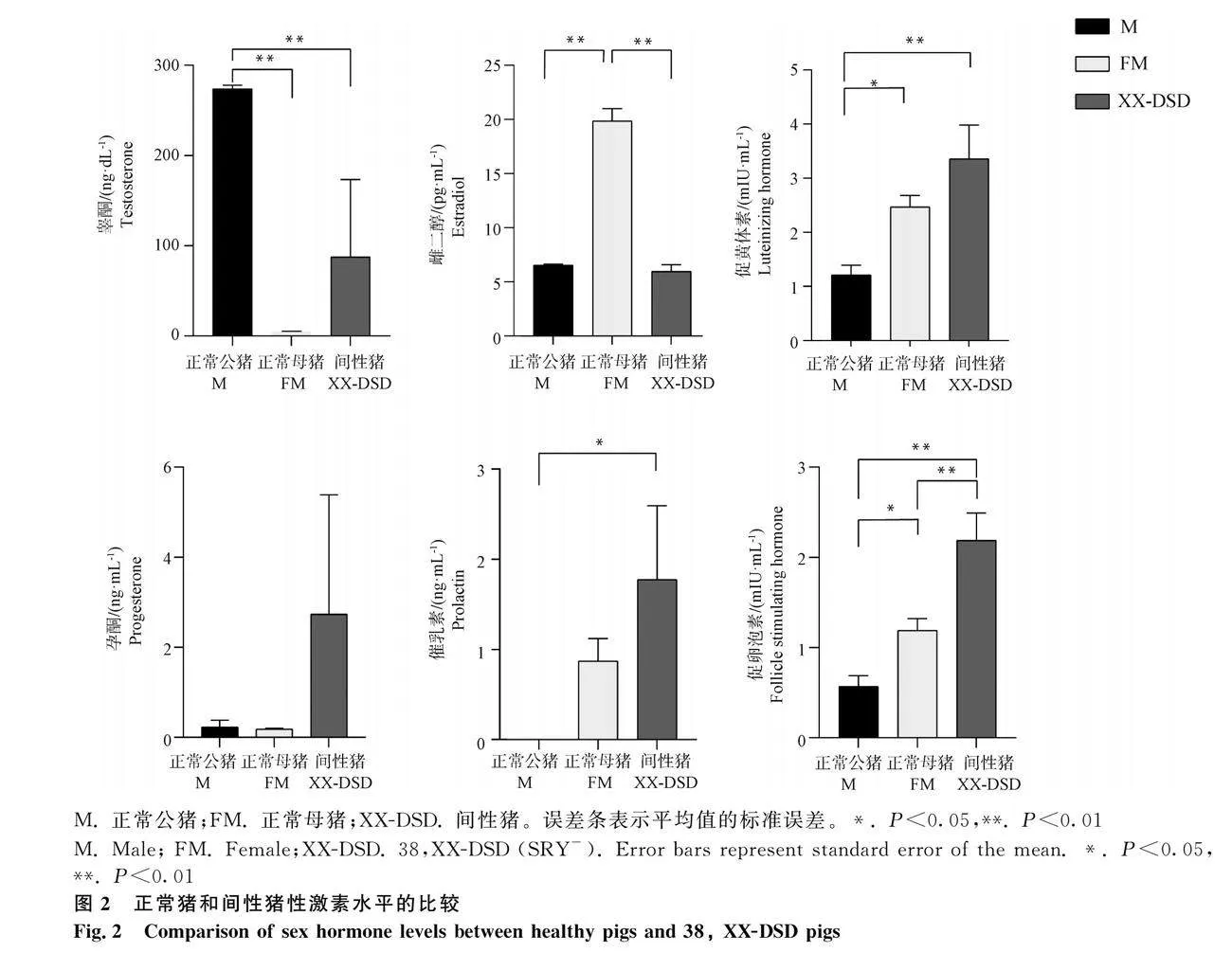
2.2 間性豬性激素水平檢測
由圖2可見,XX-DSD血清中的TE含量處于正常公豬與正常母豬之間,而XX-DSD血清中的E2含量則與TE相反;XX-DSD血清中LH、FSH、PRL和P含量均高于正常豬。
2.3 間性豬和母豬垂體組織全轉錄組差異表達分析
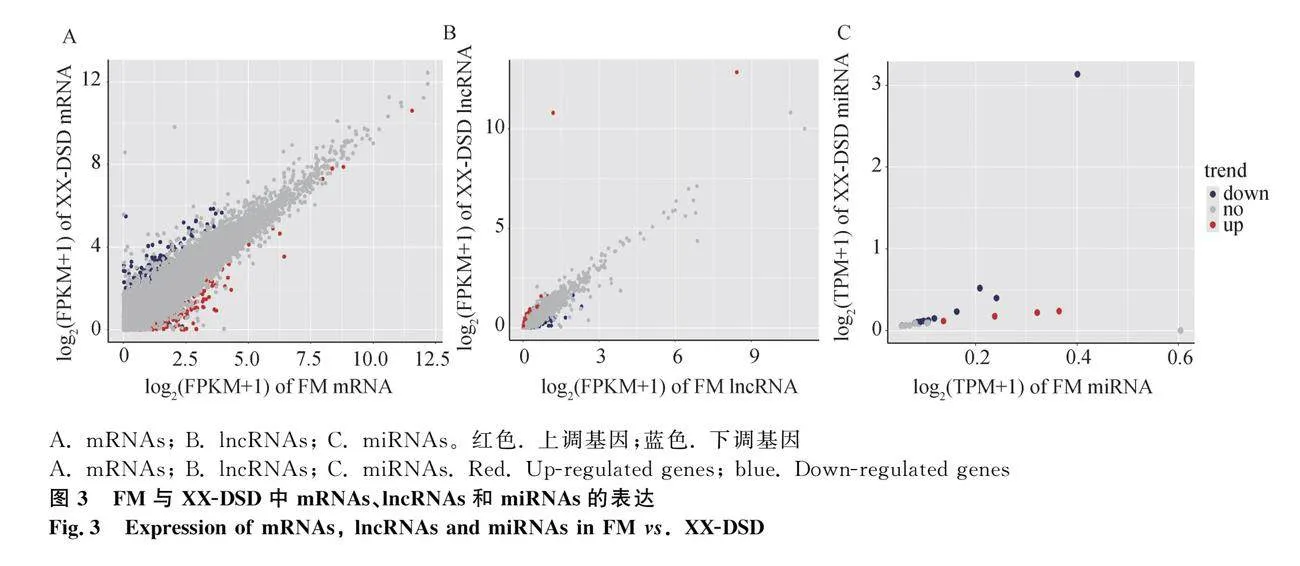
DESeq2分析結果顯示,以正常母豬垂體為對照,間性豬垂體差異表達mRNAs有1 451個,其中528個上調,923個下調(圖3A);差異表達lncRNAs有277個,其中117個上調,160個下調(圖3B);差異表達miRNAs有17個,其中4個上調,13個下調(圖3C)。
2.4 DSD差異mRNAs功能富集分析
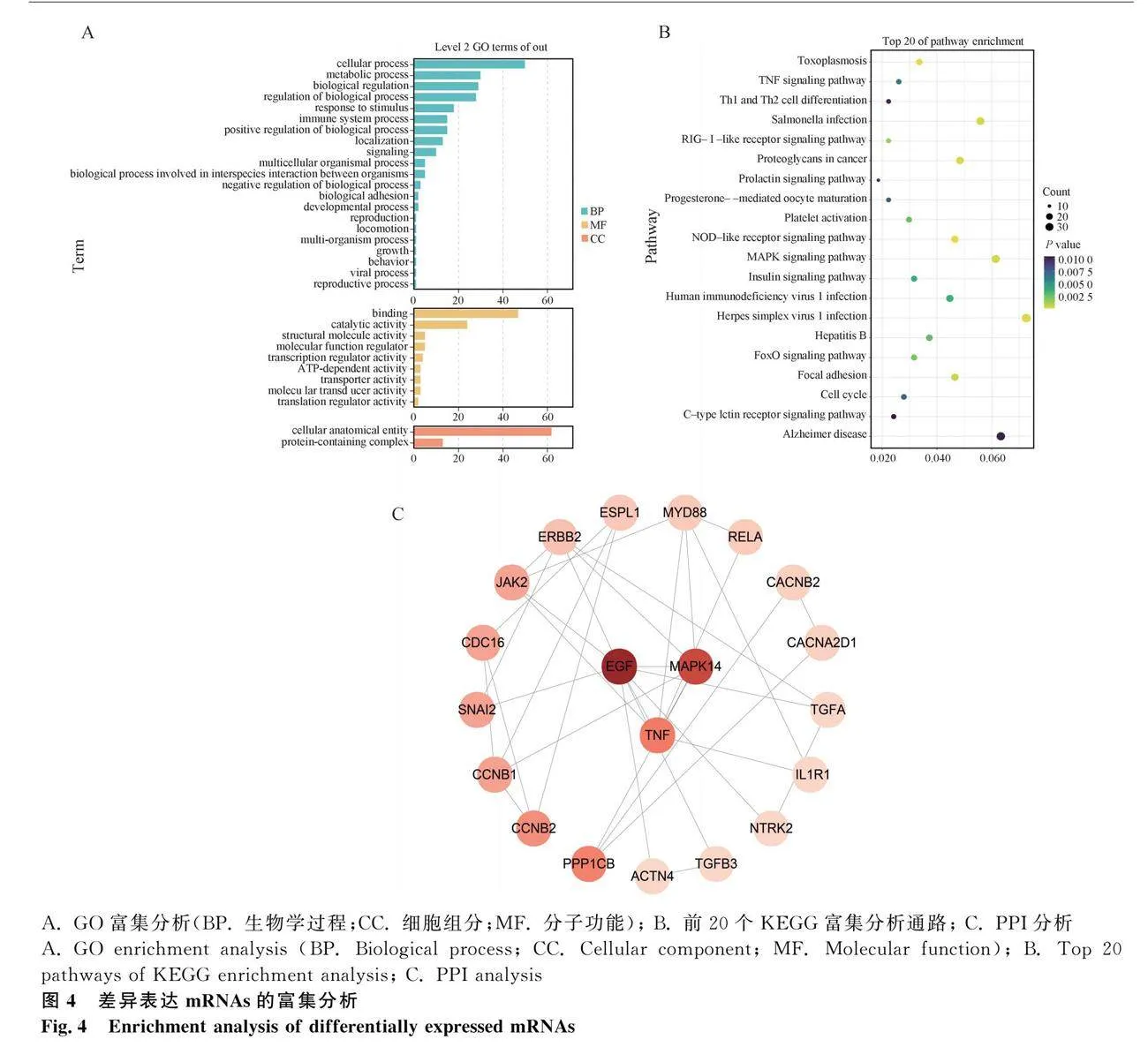
2.4.1 差異mRNAs的富集分析
對差異mRNAs進行GO(圖4A)和KEGG(圖4B)富集分析。GO結果顯示,基因顯著富集于細胞運動、分子轉導活性、生物調控等264個通路上(P<0.05)。KEGG結果顯示,差異mRNAs富集到68個KEGG通路中,并篩選出富集于MAPK信號通路、孕酮介導的卵母細胞成熟、PRL信號通路、卵母細胞減數分裂通路中的DEMs共49個,其中MAPK12為4個通路共同表達的基因。
2.4.2 差異mRNAs的PPI分析
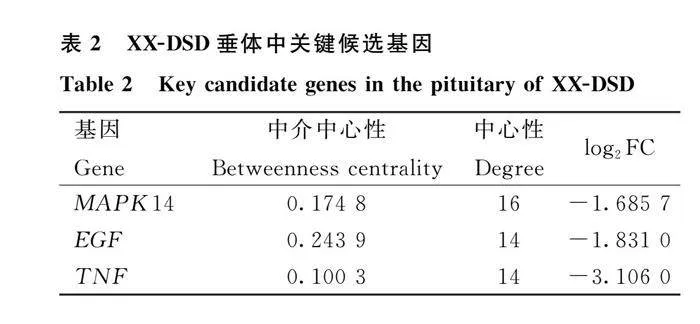
整合GO和KEGG結果,以候選mRNAs制作蛋白互作網絡圖(protein-protein interaction,PPI),如圖4C所示,同時篩選出3個與細胞信號傳導相關的關鍵候選基因,分別為絲裂原活化蛋白激酶14(mitogen-activated protein kinase 14,MAPK14)、上皮生長因子基因(epidermal growth factor,EGF)、腫瘤壞死因子基因(tumour necrosis factor,TNF)(表2)。
2.5 ceRNA互作調控網絡分析
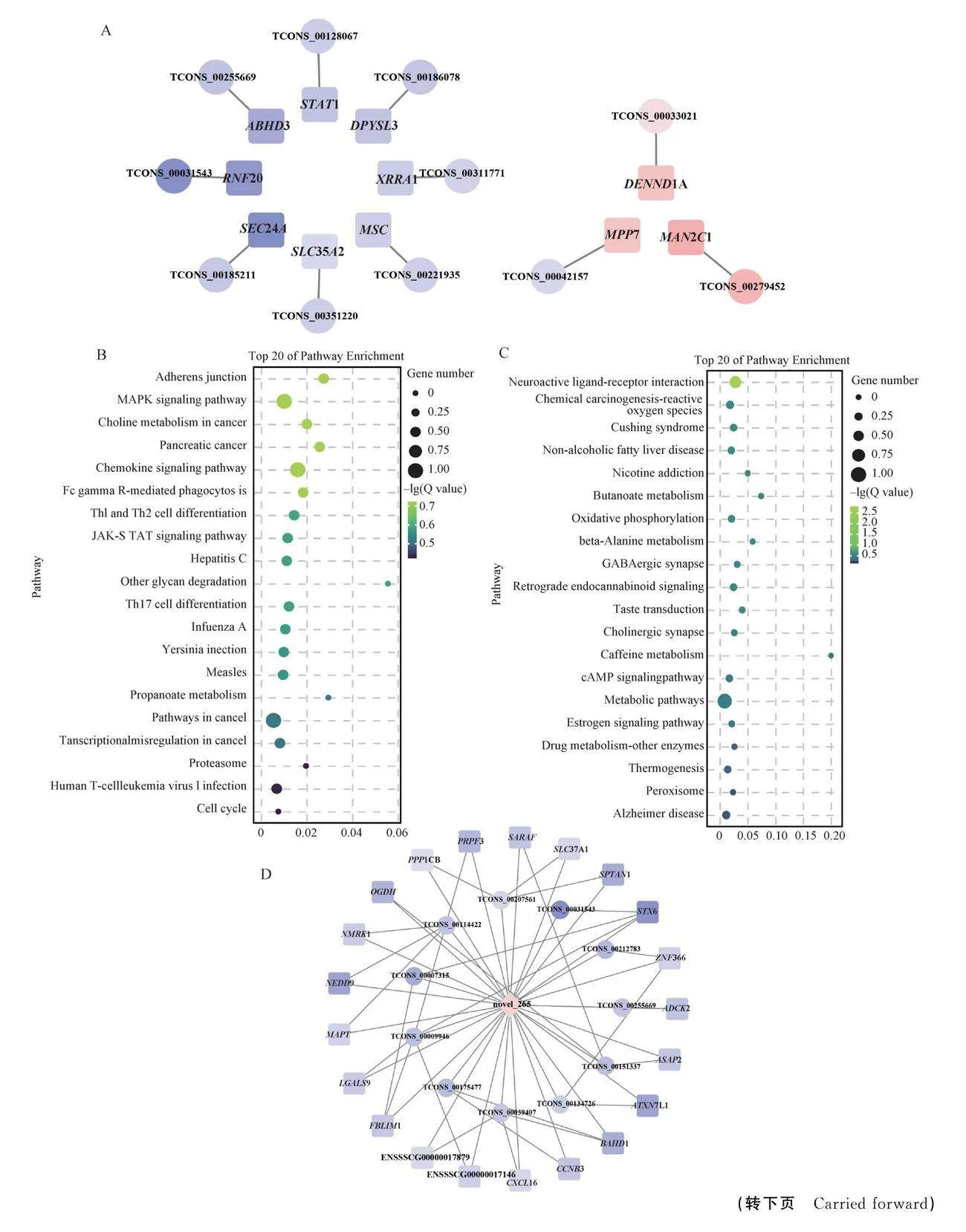
本研究首先篩選出垂體277個lncRNAs上下游100 kb的差異表達基因。lncRNAs可與miRNA競爭性結合,解除miRNA對靶基因的抑制。選擇共表達且符合篩選條件的miRNAs-mRNAs(圖5A)、miRNAs-lncRNAs(圖5B)和miRNAs-lncRNAs(圖5C)。首先,構建了差異mRNAs和lncRNAs的互作網絡圖如圖6A所示,垂體篩選出11對mRNAs-lncRNAs,包含了mRNAs和lncRNAs各11個,僅TCONS_00042157-MPP7表現為負調控,其余均為正調。值得注意的是,STAT1參與垂體PRL信號通路,TCONS_00128067-STAT1能參與間性豬垂體PRL的合成與分泌。同時對lncRNAs(圖6B)和miRNAs(圖6C)預測的靶基因進行KEGG分析,發現在cAMP信號通路、MAPK信號通路和雌激素信號通路等與垂體合成或分泌直接有關的信號通路。進一步地,整合上述數據,篩選競爭性共表達triplets,并以之建立ceRNA網絡。在FM和XX-DSD組中只篩選出26個競爭性共表達triplets。
如圖6D所示,構建的ceRNA網絡僅包含1個miRNA、21個mRNAs和11個lncRNAs。GO分析發現,CCNB3和ZNF366與激素結合、減數分裂調節相關。
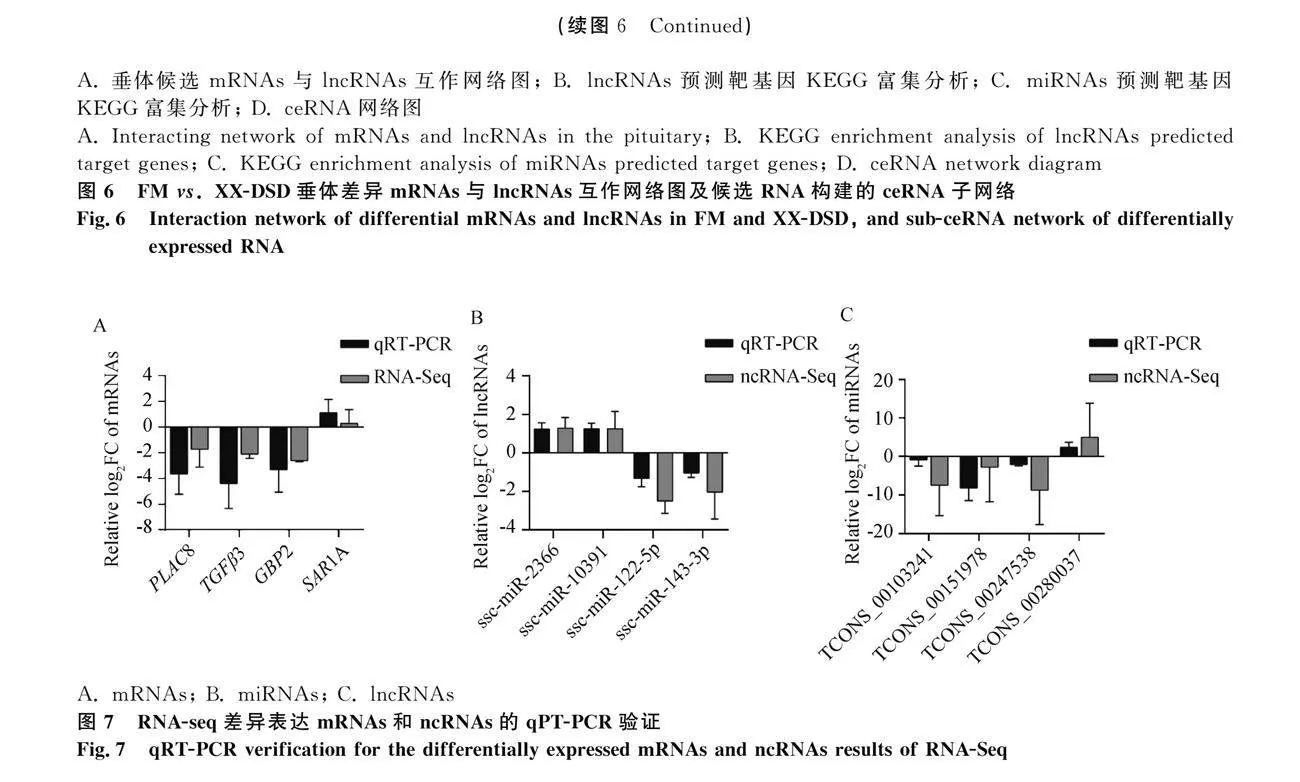
2.6 全轉錄組測序結果的qRT-PCR驗證
為驗證全轉錄組測序結果的準確性,對FM和XX-DSD垂體全轉錄組差異表達譜中隨機選擇的4個mRNAs(圖7A)、4個miRNAs(圖7B)和4個lncRNAs(圖7C)進行驗證,結果表明,所選擇的編碼RNA的qRT-PCR結果與全轉錄組測序結果相近,表明本研究的測序數據具較高準確性。
3 討 論
性腺發育是實現生殖能力的關鍵步驟,多個基因參與其過程。性別發育異常(DSD)是一種先天性疾病[18],導致生殖器官發育畸形,影響了生殖功能。研究表明在WT1的ZF4域發現的突變是46,XX-DSD (SRY-)的發病原因之一[19]。在46,XX中,腎上腺類固醇生成障礙是生殖器異常的常見原因[20]。激素紊亂是DSD中主要的臨床癥狀之一。
下丘腦-垂體-性腺軸(hypothalamic-pituitary-gonadal axis,HPGA)是控制性腺發育和性激素合成的主要內分泌系統[21],在人類和豬中,HPGA的異常可能導致性發育和生殖功能的異常[22]。課題組前期針對間性豬下丘腦的研究顯示,激素受體基因ESR1、前列腺素F受體(PTGFR)、促甲狀腺激素受體(TSHR)、孕激素受體(PGR)顯著上調,XX-DSD內分泌系統已經失常[22]。在本研究中,XX-DSD血清LH和FSH水平均高于正常的雌性和雄性,進一步說明其性腺功能不全。FSH和LH分別由一個共有的α-亞基(αGSU)和一個獨特的β-亞基(FSHβ或LHβ)組成的糖蛋白二聚體[23],其中αGSU由CGA基因編碼,β-亞基由FSHβ和LHβ基因編碼[24]。下丘腦通過控制垂體分泌的激素來調節機體的內環境穩定[25],下丘腦分泌的促性腺激素釋放激素(gonadotropin-releasing hormone,GnRH)在垂體能通過激活一系列通路,包括cAMP信號通路、Calcineurin/NFAT(nuclear factor of activated T cells)信號通路和MAPK信號通路、CaMK II(calcium/calmodulin-dependent kinase II)信號等,促進垂體CGA、FSHβ、LHβ基因的表達,從而合成和分泌LH和FSH[24]。此外,對差異表達的mRNAs進行富集分析,這些mRNAs在細胞通訊、信號傳導、細胞運動等通路上表達,特別是在MAPK通路中,MAPK1/3、MAPK8/9、MAPK14基因的表達下調,可能涉及參與GnRH信號通路的傳遞[26-27],參與誘導LHβ和αGSU的合成[28]。通過XX-DSD垂體中相關DEMs的PPI網絡分析,篩選出3個關鍵候選基因MAPK14、EGF和TNF,其表達量均下降。但是,Haisenleder等[29]研究指出,抑制MAPK14的表達對FSHβ和LHβ的表達以及促性腺激素的表達未產生顯著影響,而MAPK8/9則參與LHβ基因的調控[30]。在本研究中,MAPK9以及MAPK信號通路的基因多數為下調表達趨勢,但是LH含量仍然升高,這可能是由于LH和FSH的分泌受多種通路共同參與調控[31-32]。例如,近年越來越多的研究發現,由腺苷酸環化酶激活肽基因(adenylate cyclase activating polypeptide 1,ADCYAP1)編碼的PACAP可以單獨或者與GnRH共同調節促性腺激素的分泌[33-35],其在多個器官中表達,也可在GnRH和性腺激素的誘導下在促性腺激素細胞中合成[34,36]。PACAP可以增加促性腺激素亞基的啟動子活性[37],而PACAP受體1(ADCYAP1R1)則可以增強GnRH的調節作用[38]。ADCYAP1/ADCYAP1R1表達上調可能是造成GNRH誘發LH、FSH分泌異常升高的主要原因。
此外,TGFβ與垂體前葉功能的調節有關[39]。TGFβ3參與垂體前葉PRL細胞和Folliculo-stellate(FS)細胞之間相互作用[40]。FS細胞為非分泌細胞,通過細胞縫隙連接影響 PRL細胞的增殖和性腺釋放因子的釋放[41-42]。而TGFβ3 能通過促進FS細胞堿性成纖維細胞生長因子(basic fibroblast growth factor, bFGF)的分泌,增強細胞縫隙連接通訊[43]。而本研究發現, TGFβ3 表達量減少或將干擾 FS細胞和 PRL細胞間的溝通,影響PRL細胞的增殖。盡管 XX-DSD垂體內 PRL的含量升高,但是 PRL信號通路表達量卻下降。在PRL通路中,酪氨酸蛋白激酶2基因(Janus kinase 2, JAK2)參與PRL介導的細胞凋亡,是細胞凋亡途徑的起始激酶。當JAK2 被抑制時,內源性 PRL誘導的PRLR激活的效應未被觀察到[44]。信號轉導及轉錄激活蛋白家族(signal transducer and activator of transcription, STAT)是PRL的重要傳感器,包括了STAT1、STAT3和STAT5a/b[45]。在XX-DSD垂體中JAK2、STAT1基因表達量明顯下降進一步說明PRL通路受到抑制。結合課題組前期研究發現,XX-DSD垂體 PRL細胞較FM數量多,推測盡管TGFβ3表達量減少影響 PRL細胞的增殖,但PRL信號通路基因降低,使PRL細胞自噬減少,導致PRL的高分泌[9]。這些發現將JAK2、STAT1等基因可作為探索XX-DSD垂體功能異常的候選基因。
通過構建ceRNA網絡圖,僅發現novel_265參與ceRNA的調控,TCONS_00175477-novel_265-CCNB3、TCONS_00134726-novel_265-ZNF366和TCONS_00212783-novel_ 265-ZNF366這3個競爭組合與激素、細胞減數分裂相關。CCNB3與卵母細胞第二次減數分裂相關[46]。ZNF366是多種核受體的共調節因子[47-50],可直接與DNA結合,從而介導參與多種生物過程的基因轉錄,如增殖、分化、代謝、繁殖[51-52]。ZNF366能抑制I類核受體(nuclear receptors,NRs)介導的基因表達,促進II類NRs介導的基因表達[52]。研究指出,其鋅指區域能與ERα的鋅指區域(DNA binding domain,DBD)相互作用,召集抑制雌激素受體(estrogen receptor,ERα)表達的因子,如c末端結合蛋白(C-terminal binding protein,CtBP)、RIP140等,抑制ERα的活性,從而抑制雌激素反應基因的表達[50]。在XX-DSD垂體中ZNF366的表達降低,然而ESR1的表達量也同樣降低。此外,其他ERα的共同抑制因子,如N-CoR[53]、LCoR[54]、CtBP[55]表達量同樣有所下降。在COS-1細胞中添加E2時,ERα活性提高,但是隨著ZNF366的轉染濃度升高,ERα活性100%消失[50]。結合本研究中的XX-DSD激素水平檢測,發現其E2水平較FM的低,推測激素水平的低下是造成ERα活性下降的首要因素[56-57]。
4 結 論
間性豬對生豬種業構成了嚴重的影響,全轉錄組測序鑒定出間性豬垂體中有1 451個mRNAs、17個miRNAs和277個lncRNAs,共鑒定了26個lncRNA-miRNA-mRNA共表達競爭三聯體。其中TCONS_00175477-novel_265-CCNB3、TCONS_00134726-novel_265-ZNF366和TCONS_00212783-novel_ 265-ZNF366可能參與間性豬垂體的激素合成與分泌的調控。
參考文獻(References):
[1] REYES A P,LEN N Y,FROST E R,et al.Genetic control of typical and atypical sex development[J].Nat Rev Urol, 2023,20(7):434-451.
[2] 鐘秉洲,吳金華,于聰穎,等.38,XX-DSD(SRY-)間性豬分子病因學的研究進展[J].中國畜牧雜志,2024,60(6):107-112.
ZHONG B Z,WU J H,YU C Y,et al.Advances in the molecular pathogenesis of hermaphroditic pigs(38,XX-DSD,SRY-)[J]. Chinese Journal of Animal Science,2024,60(6):107-112.(in Chinese)
[3] HUGHES I A,HOUK C,AHMED S F,et al.Consensus statement on management of intersex disorders[J].Arch Dis Child,2006,91(7):554-563.
[4] MAMGAIN A,SAWYER I L,TIMAJO D A M,et al.RFamide-related peptide neurons modulate reproductive function and stress responses[J].J Neurosci,2021,41(3):474-488.
[5] ZHANG S,CUI Y L,MA X Y,et al.Single-cell transcriptomics identifies divergent developmental lineage trajectories during human pituitary development[J].Nat Commun,2020,11(1):5275.
[6] PECULIS R,MANDRIKA I,PETROVSKA R,et al.Pituispheres contain genetic variants characteristic to pituitary adenoma tumor tissue[J].Front Endocrinol,2020,11:313.
[7] EYAREFE O D,ATAWALNA J,EMIKPE B O,et al.Intersex piglet with bilobed urinary bladder in Kumasi,Ghana:A case report[J].Anim Res Int,2017,14(2):2720-2724.
[8] NOWACKA-WOSZUK J,SZCZERBAL I,STACHOWIAK M,et al.Association between polymorphisms in the SOX9 region and canine disorder of sex development (78,XX; SRY-negative) revisited in a multibreed case-control study[J].PLoS One,2019,14(6):e0218565.
[9] 周 怡,趙海全,劉玉清,等.一例真間性豬的研究[J].中國農業科學,2014,47(10):2021-2029.
ZHOU Y,ZHAO H Q,LIU Y Q,et al.Study of a true hermaphrodite pig[J].Scientia Agricultura Sinica,2014,47(10): 2021-2029.(in Chinese)
[10] CHEN X,SUN Y Z,GUAN N N,et al.Computational models for lncRNA function prediction and functional similarity calculation[J].Brief Funct Genomics,2019,18(1):58-82.
[11] PAILHOUX E,VIGIER B,SCHIBLER L,et al.Positional cloning of the PIS mutation in goats and its impact on understanding mammalian sex-differentiation[J].Genet Sel Evol,2005,37 Suppl 1(S1):S55-S64.
[12] WAN Z,YANG H,CHEN P Y,et al.The novel competing endogenous long noncoding RNA SM2 regulates gonadotropin secretion in the hu sheep anterior pituitary by targeting the oar-miR-16b/TGF-β/SMAD2 signaling pathway[J]. Cells,2022,11(6):985.
[13] MOKABBER H,NAJAFZADEH N,VARDIN M M.miR-124 promotes neural differentiation in mouse bulge stem cells by repressing Ptbp1 and Sox9[J].J Cell Physiol,2019,234(6):8941-8950.
[14] HE J,XU S R,JI Z J,et al.The role of miR-7 as a potential switch in the mouse hypothalamus-pituitary-ovary axis through regulation of gonadotropins[J].Mol Cell Endocrinol,2020,518:110969.
[15] ZHAO H Y,YIN X Z,XU H T,et al.LncTarD 2.0:an updated comprehensive database for experimentally-supported functional lncRNA-target regulations in human diseases[J].Nucleic Acids Res,2023,51(D1):D199-D207.
[16] CHEN X,SHI W.Genome-wide characterization of coding and non-coding RNAs in the ovary of honeybee workers and queens[J].Apidologie,2020,51(5):777-792.
[17] CAPRA E,LAZZARI B,FRATTINI S,et al.Distribution of ncRNAs expression across hypothalamic-pituitary-gonadal axis in Capra hircus[J].BMC Genomics,2018,19(1):417.
[18] EGGERS S,SADEDIN S,VAN DEN BERGEN J A,et al.Disorders of sex development:insights from targeted gene sequencing of a large international patient cohort[J].Genome Biol,2016,17(1):243.
[19] EOZENOU C,GONEN N,TOUZON M S,et al.Testis formation in XX individuals resulting from novel pathogenic variants in Wilms’ tumor 1 (WT1) gene[J].Proc Natl Acad Sci U S A,2020,117(24):13680-13688.
[20] FINKIELSTAIN G P,VIEITES A,BERGAD I,et al.Disorders of sex development of adrenal origin[J].Front Endocrinol,2021,12:770782.
[21] TSUKAMURA H.Kobayashi Award 2019:The neuroendocrine regulation of the mammalian reproduction[J].Gen Comp Endocrinol,2022,315:113755.
[22] TAN S W,ZHOU Y,ZHAO H Q,et al.Comprehensive transcriptome analysis of hypothalamus reveals genes associated with disorders of sex development in pigs[J].J Steroid Biochem Mol Biol,2021,210:105875.
[23] RIVERA-HERNNDEZ A,MADRIGAL-GONZLEZ M M,ESPINOSA-PENICHE R,et al.Van Wyk-Grumbach syndrome and trisomy 21[J].Proc (Bayl Univ Med Cent),2022,35(4):569-571.
[24] STAMATIADES G A,CARROLL R S,KAISER U B.GnRH-A key regulator of FSH[J].Endocrinology,2019,160(1):57-67.
[25] MARTINEZ-ARMENTA M,DE LEN-GUERRERO S D,CATALN A,et al.TGFβ2 regulates hypothalamic Trh expression through the TGFβ inducible early gene-1 (TIEG1) during fetal development[J].Mol Cell Endocrinol, 2015,400:129-139.
[26] SHAN B B,LIU Y,YANG C P,et al.Comparative transcriptomic analysis for identification of candidate sex-related genes and pathways in Crimson seabream (Parargyrops edita)[J].Sci Rep,2021,11(1):1077.
[27] 徐俊杰,張璐通,王津潔,等.基于多組學與網絡藥理學探究淫羊藿對后備母豬發情的作用[J].畜牧獸醫學報,2024, 55(4):1615-1628.
XU J J,ZHANG L T,WANG J J,et al.Exploring the effect of epimedium on estrus of gilts based on multiomics and network pharmacology[J].Acta Veterinaria et Zootechnica Sinica,2024,55(4):1615-1628.(in Chinese)
[28] YANG L Q,WANG H,SONG S J,et al.Systematic understanding of anti-aging effect of coenzyme Q10 on oocyte through a network pharmacology approach[J].Front Endocrinol (Lausanne),2022,13:813772.
[29] HAISENLEDER D J,BURGER L L,WALSH H E,et al.Pulsatile gonadotropin-releasing hormone stimulation of gonadotropin subunit transcription in rat pituitaries:evidence for the involvement of Jun N-terminal kinase but not p38[J].Endocrinology,2008,149(1):139-145.
[30] BURGER L L,HAISENLEDER D J,AYLOR K W,et al.Regulation of Lhb and Egr1 gene expression by GNRH pulses in rat pituitaries is both c-Jun N-terminal kinase (JNK)- and extracellular signal-regulated kinase (ERK)-dependent[J].Biol Reprod,2009,81(6):1206-1215.
[31] WANG Y,BERNARD D J.Activin A induction of murine and ovine follicle-stimulating hormone β transcription is SMAD-dependent and TAK1 (MAP3K7)/p38 MAPK-independent in gonadotrope-like cells[J].Cell Signal,2012,24(8): 1632-1640.
[32] HOLLANDER-COHEN L,GOLAN M,LEVAVI-SIVAN B.Differential regulation of gonadotropins as revealed by transcriptomes of distinct LH and FSH cells of fish pituitary[J].Int J Mol Sci,2021,22(12):6478.
[33] KANASAKI H,MIJIDDORJ T,SUKHBAATAR U,et al.Pituitary adenylate cyclase-activating poly-peptide (PACAP) increases expression of the gonadotropin-releasing hormone (GnRH) receptor in GnRH-producing GT1-7 cells overexpressing PACAP type I receptor[J].Gen Comp Endocrinol,2013,193:95-102.
[34] ZHENG W M,GRAFER C M,HALVORSON L M.Interaction of gonadal steroids and gonadotropin-releasing hormone on pituitary adenylate cyclase-activating polypeptide (PACAP) and PACAP receptor expression in cultured rat anterior pituitary cells[J].Reprod Sci,2014,21(1):41-51.
[35] TAO W J,CHEN J L,TAN D J,et al.Transcriptome display during tilapia sex determination and differentiation as revealed by RNA-Seq analysis[J].BMC Genomics,2018,19(1):363.
[36] PURWANA I N,KANASAKI H,ORIDE A,et al.GnRH-induced PACAP and PAC1 receptor expression in pituitary gonadotrophs:a possible role in the regulation of gonadotropin subunit gene expression[J].Peptides,2010,31(9):1748-1755.
[37] MIJIDDORJ T,KANASAKI H,ORIDE A,et al.Interaction between kisspeptin and adenylate cyclase-activating polypeptide 1 on the expression of pituitary gonadotropin subunits:a study using mouse pituitary lbetaT2 cells[J].Biol Reprod,2017,96(5):1043-1051.
[38] PURWANA I N,KANASAKI H,ORIDE A,et al.Expression of the pituitary adenylate cyclase-activating polypeptide (PACAP) type 1 receptor (PAC1R) potentiates the effects of GnRH on gonadotropin subunit gene expression[J].Biochem Biophys Res Commun,2011,410(2):295-300.
[39] QIAN X,JIN L,LLOYD R V.Expression and regulation of transforming growth factor β1 in cultured normal and Neoplastic rat pituitary cells[J].Endocr Pathol,1996,7(1):77-90.
[40] HENTGES S,BOYADJIEVA N,SARKAR D K.Transforming growth factor-β3 stimulates lactotrope cell growth by increasing basic fibroblast growth factor from folliculo-stellate cells[J].Endocrinology,2000,141(3):859-867.
[41] OOMIZU S,CHATURVEDI K,SARKAR D K.Folliculostellate cells determine the susceptibility of lactotropes to estradiol’s mitogenic action[J].Endocrinology,2004,145(3):1473-1480.
[42] CHRISTIAN H C,IMIRTZIADIS L,TORTONESE D.Ultrastructural changes in lactotrophs and folliculo-stellate cells in the ovine pituitary during the annual reproductive cycle[J].J Neuroendocrinol,2015,27(4):277-284.
[43] KABIR N,CHATURVEDI K,LIU L S,et al.Transforming growth factor-β3 increases gap-junctional communication among folliculostellate cells to release basic fibroblast growth factor[J].Endocrinology,2005,146(9):4054-4060.
[44] DE DIOS N,ORRILLO S,IRIZARRI M,et al.JAK2/STAT5 pathway mediates prolactin-induced apoptosis of lactotropes[J].Neuroendocrinology,2019,108(2):84-97.
[45] DASILVA L,RUI H,ERWIN R A,et al.Prolactin recruits STAT1,STAT3 and STAT5 independent of conserved receptor tyrosines TYR402,TYR479,TYR515 and TYR580[J].Mol Cell Endocrinol,1996,117(2):131-140.
[46] 周 怡,趙海全,劉玉清,等.一例真間性豬的研究[J].中國農業科學,2014,47(10):2021-2029.
ZHOU Y,ZHAO H Q,LIU Y Q,et al.Study of a true hermaphrodite pig[J].Scientia Agricultura Sinica,2014,47(10): 2021-2029.(in Chinese)
[46] MENG T G,LEI W L,LI J,et al.Degradation of Ccnb3 is essential for maintenance of MII arrest in oocyte[J].Biochem Biophys Res Commun,2020,521(1):265-269.
[47] HONTELEZ S,KARTHAUS N,LOOMAN M W,et al.DC-SCRIPT regulates glucocorticoid receptor function and expression of its target GILZ in dendritic cells[J].J Immunol,2013,190(7):3172-3179.
[48] ANSEMS M,KARTHAUS N,HONTELEZ S,et al.DC-SCRIPT:AR and VDR regulator lost upon transformation of prostate epithelial cells[J].Prostate,2012,72(16):1708-1717.
[49] ANSEMS M,HONTELEZ S,LOOMAN M W G,et al.DC-SCRIPT:nuclear receptor modulation and prognostic significance in primary breast cancer[J].J Natl Cancer Inst,2010,102(1):54-68.
[50] LOPEZ-GARCIA J,PERIYASAMY M,THOMAS R S,et al.ZNF366 is an estrogen receptor corepressor that acts through CtBP and histone deacetylases[J].Nucleic Acids Res,2006,34(21):6126-6136.
[51] GRONEMEYER H,GUSTAFSSON J ,LAUDET V.Principles for modulation of the nuclear receptor superfamily[J].Nat Rev Drug Discov,2004,3(11):950-964.
[52] ANSEMS M,SNDERGAARD J N,SIEUWERTS A M,et al.DC-SCRIPT is a novel regulator of the tumor suppressor gene CDKN2B and induces cell cycle arrest in ERα-positive breast cancer cells[J].Breast Cancer Res Treat,2015,149(3): 693-703.
[53] JACKSON T A,RICHER J K,BAIN D L,et al.The partial agonist activity of antagonist-occupied steroid receptors is controlled by a novel hinge domain-binding coactivator L7/SPA and the corepressors N-CoR or SMRT[J].Mol Endocrinol,1997,11(6):693-705.
[54] FERNANDES I,BASTIEN Y,WAI T,et al.Ligand-dependent nuclear receptor corepressor LCoR functions by histone deacetylase-dependent and -independent mechanisms[J].Mol Cell,2003,11(1):139-150.
[55] VO N,FJELD C,GOODMAN R H.Acetylation of nuclear hormone receptor-interacting protein RIP140 regulates binding of the transcriptional corepressor CtBP[J].Mol Cell Biol,2001,21(18):6181-6188.
[56] PREZ P A,TOLEDO J,DEL VALLE SOSA L,et al.The phthalate DEHP modulates the estrogen receptors α and β increasing lactotroph cell population in female pituitary glands[J].Chemosphere,2020,258:127304.
[57] GUZMN J M,LUCKENBACH J A,DA SILVA D A M,et al.Seasonal variation of pituitary gonadotropin subunit,brain-type aromatase and sex steroid receptor mRNAs,and plasma steroids during gametogenesis in wild sablefish[J].Comp Biochem Physiol Part A Mol Integr Physiol,2018,219-220:48-57.
(編輯 郭云雁)
