轉(zhuǎn)錄組和蛋白組篩選就巢雞卵巢發(fā)育候選基因及其調(diào)控網(wǎng)絡(luò)構(gòu)建
2024-12-18 00:00:00蔣婷李文東李興起黃雨王啟貴王海威楊朝武劉凌斌
畜牧獸醫(yī)學(xué)報 2024年11期





摘 要: 為了研究蛋雞就巢性的潛在調(diào)控機制,本研究選用300日齡體況良好、體重一致的城口山地雞12只,其中3只個體處于正常產(chǎn)蛋期,其余9只個體分別處于就巢期10、20和30天。采集各組蛋雞卵巢組織進(jìn)行組織解剖學(xué)形態(tài)觀察,然后利用轉(zhuǎn)錄組(RNA-seq)和蛋白組(iTRAQ)測序技術(shù)分別對產(chǎn)蛋組正常卵巢(normal ovary,NO)和就巢組萎縮卵巢(atrophic ovary,AO)進(jìn)行測序分析。篩選差異表達(dá)基因(DEGs)和差異表達(dá)蛋白(DEPs)并進(jìn)行功能富集分析,鑒定與家禽就巢性相關(guān)的候選基因。結(jié)果表明:就巢導(dǎo)致蛋雞卵巢發(fā)生萎縮,卵泡大量閉鎖。在卵巢組織中鑒定出930個DEGs,其中430個基因上調(diào),500個基因下調(diào);同時鑒定出546個DEPs,其中178個蛋白上調(diào),368個蛋白下調(diào)。通過功能注釋和富集分析發(fā)現(xiàn),這些DEGs和DEPs顯著富集在細(xì)胞外基質(zhì)(extracellular matrix,ECM)受體相互作用、黏著和PI3K-Akt等信號通路。最后通過轉(zhuǎn)錄組和蛋白組聯(lián)合分析,篩選出7個蛋雞就巢性的候選基因:COMP、FN1、ITGA8、THBS1、COL4A2、COL4A1和COL1A1。綜上所述,本研究通過分析轉(zhuǎn)錄組測序和蛋白組測序結(jié)果篩選了蛋雞就巢性關(guān)鍵候選基因,并構(gòu)建了家禽就巢狀態(tài)下卵巢發(fā)育的調(diào)控網(wǎng)絡(luò)。本研究為深入解析就巢期卵巢萎縮的分子調(diào)控機制提供了理論參考,豐富了調(diào)控家禽就巢性狀的候選基因,為蛋雞的遺傳改良和分子育種研究提供理論依據(jù)。
關(guān)鍵詞: 就巢;卵巢;轉(zhuǎn)錄組學(xué);蛋白組學(xué);城口山地雞
中圖分類號:S831.2
文獻(xiàn)標(biāo)志碼:A
文章編號:0366-6964(2024)11-4950-18
收稿日期:2024-05-13
基金項目:重慶市研究生科研創(chuàng)新項目(CYS21126);重慶市技術(shù)創(chuàng)新與應(yīng)用發(fā)展專項重點項目(CSTB2023TIAD-LUX0003);西南大學(xué)大學(xué)生創(chuàng)新創(chuàng)業(yè)訓(xùn)練計劃項目(202310635032)
作者簡介:蔣 婷(1995-),女,甘肅通渭人,碩士生,主要從事家禽遺傳育種研究,E-mail:jiangt019@163.com
*通信作者:劉凌斌,主要從事家禽遺傳育種研究,E-mail:liulb515@163.com
Screening Candidate Genes for Ovarian Development and Constructing Regulatory Network
in Nesting Chickens by Transcriptome and Proteome
JIANG" Ting1, LI" Wendong1, LI" Xingqi1, HUANG" Yu1, WANG" Qigui2, WANG" Haiwei2, YANG" Chaowu3, LIU" Lingbin1*
(1.College of Animal Science and Technology, Southwest University, Chongqing 400715," China;
2.Chongqing Academy of Animal Sciences, Chongqing 402460," China;
3.Sichuan Animal Science
Academy, Chengdu 610066," China)
Abstract:" The study aimed to investigate the potential regulatory mechanism of nesting ability of laying hens. The 12 300-day-old Chengkou mountain chickens with good body condition and the same body weight were used in this study. Among them, 3 individuals were in normal laying period, and the remaining 9 individuals were in nesting period for 10, 20 and 30 days, respectively.
The ovarian tissues of laying hens in each group were collected for anatomical observation.
Normal ovary (NO) and atrophic ovary (AO) of laying hens were analyzed by RNA-seq and iTRAQ sequencing technology. Differentially expressed genes (DEGs) and differentially expressed proteins (DEPs) were screened and functional enrichment analysis was performed to identify candidate genes related to nesting in poultry. The results showed that ovary atrophy and follicle atresia occurred in nesting hens. A total of 930 DEGs were identified in ovarian tissues, of which 430 genes were up-regulated and 500 genes were down-regulated. Meanwhile, 546 DEPs were identified, of which 178 proteins were up-regulated and 368 proteins were down-regulated. Through functional annotation and enrichment analysis, it was found that these DEGs and DEPs were significantly enriched in ECM receptor interaction, adhesion and PI3K-Akt signaling pathways. Finally, 7 candidate genes for nesting ability COMP, FN1, ITGA8, THBS1, COL4A2, COL4A1 and COL1A1 were selected by combined transcriptome and proteome analysis. In summary, key candidate genes for nesting ability were identified by transcriptome and proteome analysis, and a regulatory network for ovarian development in nesting poultry was constructed. This study provides a theoretical reference for further understanding the molecular regulation mechanism of ovarian atrophy at nesting stage, enriches the candidate genes regulating poultry nesting traits, and provides theoretical basis for genetic improvement and molecular breeding research of laying hens.
Key words: nesting; ovary; transcriptome; proteome;Chengkou mountain chicken
*Corresponding author:LIU Lingbin, E-mail:liulb515@163.com
就巢是母禽具有的一種天然母性行為,主要表現(xiàn)為頻繁窩巢、被毛蓬松、食欲衰退、停止產(chǎn)蛋并進(jìn)行孵化等行為[1]。同時,就巢的母禽會伴隨有體溫升高、體重顯著下降、免疫力降低、反應(yīng)遲緩、活動減少、攻擊性變強等一系列生理狀況的變化。母禽在就巢狀態(tài)下,會出現(xiàn)卵泡閉鎖、卵巢萎縮、排卵停止、產(chǎn)蛋量下降甚至停止產(chǎn)蛋的現(xiàn)象。除了白來航雞及相關(guān)商品化品系雞不表現(xiàn)出就巢行為,大部分家禽都會表現(xiàn)出不同程度的就巢行為[2]。頑固的就巢性是中國大多數(shù)地方家禽品種產(chǎn)蛋性能低下的主要原因之一。開展就巢性分子調(diào)控機制的研究有助于找到控制就巢性狀的主效基因或關(guān)鍵性靶點分子,最終通過遺傳改良的方法培育出低就巢性或無就巢性的品種(系),進(jìn)而提高蛋雞的產(chǎn)蛋性能。
家禽的就巢性是一種由多基因控制的復(fù)雜性狀,通常受到遺傳、內(nèi)分泌、環(huán)境等因素的調(diào)控[3-4]。就巢性的遺傳力很低,常規(guī)育種方法很難徹底消除就巢性。下丘腦-垂體-性腺(HPG)軸是調(diào)節(jié)家禽繁殖性能的關(guān)鍵調(diào)節(jié)軸,卵巢在其中發(fā)揮著重要作用[5]。卵巢分泌多種激素,如孕酮(progesterone,PROG)、雌二醇(estradiol,E2)、促卵泡激素(follicle-stimulating hormone,F(xiàn)SH)、促黃體素(luteinizing hormone,LH)和催乳素(prolactin,PRL)等,這些激素被認(rèn)為是與就巢相關(guān)的關(guān)鍵激素,其中大部分是類固醇激素[6-7]。這些激素影響卵母細(xì)胞和卵泡的發(fā)育和成熟,最終促進(jìn)或抑制排卵。先前的研究表明,雞的卵泡閉鎖受到顆粒細(xì)胞凋亡的調(diào)控[8]。顆粒細(xì)胞凋亡會導(dǎo)致哺乳動物和家禽卵泡閉鎖[9]。通常,當(dāng)發(fā)育卵泡中凋亡的顆粒細(xì)胞比例超過10%時卵泡就會發(fā)生閉鎖[10]。影響家禽就巢的內(nèi)分泌因子具有遺傳性,且受多種基因共同調(diào)控[11]。遺傳機制是導(dǎo)致家禽產(chǎn)生就巢性的根本原因,相關(guān)研究主要集中于PRL、LH、FSH等繁殖激素相關(guān)基因的表達(dá)和多態(tài)性的分析[12],結(jié)果表明這些基因位點與就巢發(fā)生和產(chǎn)蛋性能密切相關(guān)。
隨著RNA-seq和iTRAQ測序廣泛應(yīng)用于畜禽研究[13-15],許多新的生殖相關(guān)基因和蛋白質(zhì)被鑒定出來,RNA-seq和iTRAQ測序成為了探究分子機制的熱門工具。Huang等[16]對高產(chǎn)和低產(chǎn)蛋雞卵巢進(jìn)行了轉(zhuǎn)錄組測序和代謝組測序,聯(lián)合分析后篩選出4個候選基因,3條關(guān)鍵通路,為解析蛋雞產(chǎn)蛋機制提供了理論基礎(chǔ)。Wang等[17]對脂肪沉積相對較高和較低的南川黑豬背最長肌的轉(zhuǎn)錄組和蛋白組進(jìn)行分析,篩選了15個決定脂質(zhì)沉積遺傳分歧的候選基因,為地方豬品種的育種工作提供了理論指導(dǎo)。在就巢期和產(chǎn)蛋期的鵝卵巢中差異表達(dá)的基因及其蛋白質(zhì)主要富集于類固醇生物合成、類固醇激素生物合成、PI3K-Akt信號通路、孕激素介導(dǎo)的卵母細(xì)胞成熟和催產(chǎn)素信號通路,這些均是與生殖性能相關(guān)的關(guān)鍵信號通路[16-18]。同時,也有研究表明,蛋雞就巢期間一些基因富集到細(xì)胞周期、增殖、凋亡通路[7],這與就巢期家禽卵巢的形態(tài)學(xué)觀測結(jié)果相符。研究表明,抑制類固醇生物合成中的類固醇合酶會導(dǎo)致卵巢功能退化并觸發(fā)就巢行為[19]。此外,家禽繁殖期間大量的卵泡閉鎖和卵泡變性也與顆粒細(xì)胞的自噬和凋亡有關(guān)[20-21]。與細(xì)胞周期、增殖、凋亡通路和類固醇激素合成、代謝和轉(zhuǎn)運相關(guān)的基因、蛋白質(zhì)、代謝物、生長因子和信號通路仍是當(dāng)前研究的熱點。然而,這些因素如何影響家禽的卵巢結(jié)構(gòu)和功能,進(jìn)而導(dǎo)致其就巢行為的確切調(diào)控機制目前仍不清楚。本研究對蛋雞產(chǎn)蛋期正常卵巢和就巢期萎縮卵巢進(jìn)行了轉(zhuǎn)錄組和蛋白組測序,系統(tǒng)地分析了雞就巢期間轉(zhuǎn)錄水平和蛋白水平的變化,聯(lián)合分析轉(zhuǎn)錄組學(xué)和蛋白組學(xué)數(shù)據(jù),為探明家禽就巢期卵巢萎縮的關(guān)鍵分子機制提供了理論基礎(chǔ)。
1 材料與方法
1.1 動物試驗
試驗動物來源于重慶市城口山地雞遺傳研究所飼養(yǎng)的同一批次的1 700只城口山地雞。試驗雞飼養(yǎng)在采用機械通風(fēng)的封閉式雞舍,三層階梯式單體籠養(yǎng)。按照NRC(1994)蛋雞飼養(yǎng)標(biāo)準(zhǔn)飼養(yǎng),飼喂324蛋雞產(chǎn)蛋期配合飼料,雞舍設(shè)有乳頭飲水器,雞可自由采食,飲水。每天記錄雞群產(chǎn)蛋量和就巢情況,在300日齡時期,選擇同一飼養(yǎng)條件下12只體況良好、體重一致,具有典型品種外觀特征的城口山地雞作為研究材料。其中,挑選處于正常產(chǎn)蛋期的3只個體作為正常組(NO)試驗材料,挑選進(jìn)入就巢期10、20、30天的3個時間點各3只個體作為就巢10(AO-10)、20(AO-20)、30天(AO-30)的試驗材料。采集各組母雞的卵巢組織進(jìn)行觀察。NO組和AO-30組的卵巢部分置入4%多聚甲醛,用于制作石蠟切片,剩余部分液氮速凍后-80 ℃冰箱保存,用于轉(zhuǎn)錄組測序和蛋白質(zhì)提取。本研究所有試驗動物操作均嚴(yán)格符合國際動物福利合作委員會(ICCAW)和西南大學(xué)實驗室動物福利和倫理準(zhǔn)則。
1.2 蘇木精和伊紅(HE)染色
將卵巢組織在4%多聚甲醛固定液(#BL539A;Biosharp)中固定48 h,常規(guī)修剪成標(biāo)準(zhǔn)橫截面,并用石蠟處理。然后切成4 μm的切片,裝在涂有3-aminopropyltriethoxysilane的載玻片上,HE染色后觀察卵巢組織狀態(tài)。用Panoramic 250 Flash II幻燈片掃描儀掃描玻片,并用3DHISTECH全景玻片觀察儀以100倍和200倍的放大率拍照。
1.3 RNA提取、文庫構(gòu)建和測序
使用TRIzolTM LS Reagent提取卵巢組織總RNA。采用Agilent 2100生物分析儀對RNA質(zhì)量進(jìn)行評估,采用RNase-free瓊脂糖凝膠電泳進(jìn)行檢測。采用RNA完整性編號(RNA integrity number, RIN)評價總RNA的完整性。RIN值越小表示降解越嚴(yán)重,10表示完整性最高。當(dāng)RIN≥7且28S/18S≥0.7時,樣品質(zhì)量滿足試驗要求。用QIA quick RT-qPCR提取試劑盒(Qiagen)純化cDNA片段,末端修復(fù),poly(a)尾,連接Illumina測序適配器,由Gene Denovo生物技術(shù)公司使用Illumina Nova seq. 6000測序儀進(jìn)行測序。
1.4 蛋白質(zhì)提取和質(zhì)量控制
稱取適當(dāng)?shù)臉悠罚尤?×Cocktail,在冰上放置5 min后,加入10 mmol·L-1 DTT。然后用研磨機研磨組織,離心后獲得上清液。再加入10 mmol·L-1 DTT,在56 ℃水浴中放置1 h。然后加入55 mmol·L-1 IAM,在暗室中放置45 min。之后向蛋白溶液中加入預(yù)冷丙酮(1∶5),在-20 ℃下保存30 min,離心15 min(4 ℃,25 000 g)后棄上清液,將沉淀風(fēng)干后加入裂解緩沖液。第二次離心后得到蛋白質(zhì)溶液(上清液)。采用Bradford定量和SDS-PAGE進(jìn)行蛋白質(zhì)質(zhì)量控制。
1.5 數(shù)據(jù)處理和生物信息學(xué)分析
使用FASTP(version 0.18.0)軟件對測序獲得的reads進(jìn)行篩選。過濾標(biāo)準(zhǔn)如下:1)刪除包含適配器的reads數(shù)據(jù);2)去除包含超過10%未知核苷酸的序列;3)剔除含有50%以上低質(zhì)量(Q值≤20)堿基的低質(zhì)量reads。使用短讀長比對工具Bowtie225(版本2.2.8)將讀長映射到核糖體RNA(rRNA)數(shù)據(jù)庫,然后刪除映射的rRNA讀長。剩余的純序列進(jìn)一步用于組裝和基因豐度計算。使用HISAT2將干凈的配對端序列映射到參考基因組。使用StringTie v1.3.127基于參考方法組裝每個樣本的映射reads。采用StringTie軟件計算每個基因的FPKM(fragment per kilobase of transcript per million mapped reads)值,以量化其表達(dá)豐度和變異。FPKM公式如下:
FPKM=106CNL/103
設(shè)FPKM(A)為轉(zhuǎn)錄本A的表達(dá)量,C為轉(zhuǎn)錄本A映射的片段數(shù),N為映射到參考基因的片段總數(shù),L為轉(zhuǎn)錄本A上的堿基數(shù)。根據(jù)每個基因的FPKM值,通過表達(dá)分布圖顯示不同樣本基因或轉(zhuǎn)錄本的表達(dá)分布。此外,使用R程序(http://www.r-project.org/)進(jìn)行主成分分析(PCA),并計算樣本之間的Pearson相關(guān)系數(shù),了解樣本之間的可重復(fù)性,并幫助排除離群樣本。采用R軟件DESeq包以log2FCgt;1,P≤0.05為篩選標(biāo)準(zhǔn),篩選就巢期萎縮卵巢與正常產(chǎn)蛋期卵巢之間的差異表達(dá)基因(DEGs)。使用熱圖和火山圖來顯示兩組之間的差異。
1.6 基因和蛋白功能富集分析
在GO(http://www.geneontology.org/)和KEGG(https://www.genome.jp/kegg/)數(shù)據(jù)庫中檢索所有的DEGs和DEPs,計算每個詞條注釋的基因數(shù)量。GSEA(gene set enrichment analysis)分析能夠有效彌補微效基因的有效信息挖據(jù)不足等問題,更為全面地對某一功能單位(KEGG pathway,GO term)的調(diào)節(jié)作用進(jìn)行解釋。對GSEA富集分析的原理闡述見Powers等[22]的報道。
1.7 實時熒光定量PCR(qRT-PCR)驗證
采用Trizol法提取NO組和AO組卵巢組織總RNA。然后將提取的RNA按照PrimeScriptTM RT reagent Kit with gDNA Eraser說明進(jìn)行逆轉(zhuǎn)錄得到cDNA。根據(jù)測序分析結(jié)果,隨機選取FN1、COL1A1、SGK2、CASTOR1、ACTA1和MMP10,共6個差異表達(dá)基因,利用NCBI(https://www.ncbi.nlm.nih.gov/)和測序數(shù)據(jù)尋找目標(biāo)基因序列,利用生工生物、NCBI等網(wǎng)站在線設(shè)計引物(表1),引物設(shè)計完成后,利用NCBI primer blast功能進(jìn)行引物特異性比對,比對合格的引物交由華大基因合成。以反轉(zhuǎn)錄合成的定量cDNA為模板,以β-actin為內(nèi)參基因,按照TB Green premix Ex TaqTM Ⅱ定量試劑盒說明進(jìn)行反應(yīng)液配制。反應(yīng)體系共20 μL:(2×)TB Green premix Ex Taq Ⅱ (Tli RNase H Plus) 10 μL,上、下游引物各0.8 μL,cDNA 1.6 μL,ddH2O 6.8 μL。PCR擴(kuò)增程序:預(yù)變性階段為95 ℃ 30 s;循環(huán)變性階段為95 ℃ 5 s,60 ℃ 40 s(40個循環(huán));熔解曲線為95 ℃ 15 s,60 ℃ 1 min,95 ℃ 15 s。每個樣品進(jìn)行3次生物學(xué)重復(fù)。
1.8 數(shù)據(jù)統(tǒng)計分析
采用2-△△CT法計算基因相對表達(dá)量,以β-actin作為內(nèi)參基因,所有試驗數(shù)據(jù)均以“Mean±SEM”表示,使用SPSS 27.0單因素方差分析(One-way ANOVA)和獨立樣本t檢驗分析數(shù)據(jù)的顯著性,Plt;0.05(在圖中用*表示)為顯著相關(guān),Plt;0.01(在圖中用**表示)為極顯著相關(guān)。采用GraphPad Prism 8(version8.2.1)、R語言(version4.2.0)、Cytoscape(Version3.8.2)和AI(Adobe illustrator 2021)進(jìn)行可視化統(tǒng)計學(xué)分析。
2 結(jié) 果
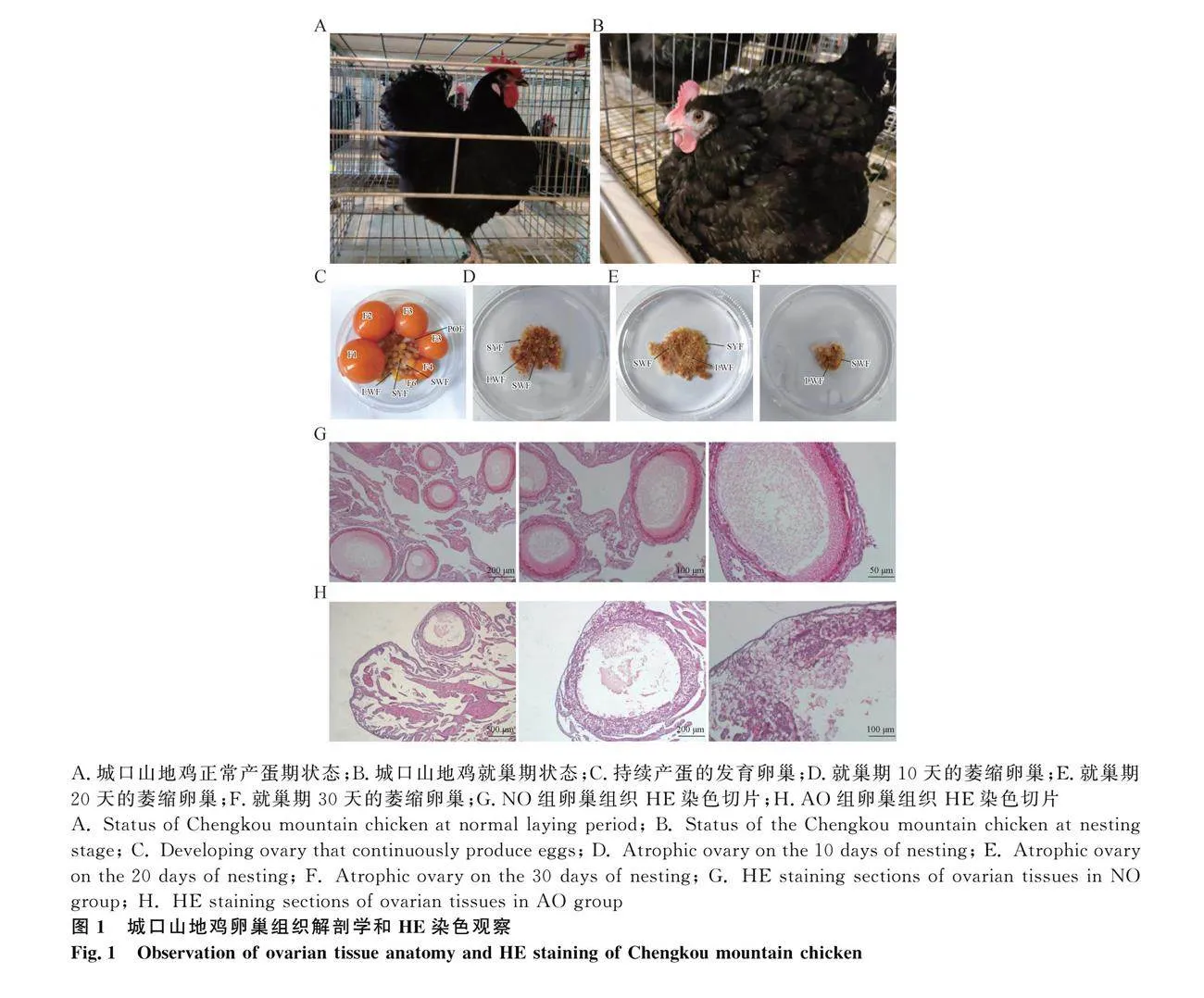
2.1 城口山地雞持續(xù)產(chǎn)蛋期與就巢期狀態(tài)和卵巢組織解剖學(xué)形態(tài)觀察
在養(yǎng)殖過程中發(fā)現(xiàn),持續(xù)產(chǎn)蛋期的城口山地雞表現(xiàn)出站立有神、翅收縮有力且緊貼體軀、動作敏捷、聽覺敏銳、食欲旺盛等特征(圖1A)。而處于就巢期的城口山地雞會發(fā)生窩巢、被毛蓬松、停止產(chǎn)蛋、食欲不振、攻擊性變強、活動減少、反應(yīng)遲緩等一系列生理狀況的變化(圖1B)。為了進(jìn)一步探究就巢期城口山地雞卵巢的變化,解剖并觀察了持續(xù)產(chǎn)蛋期和進(jìn)入就巢期10、20、30天的城口山地雞的卵巢組織。從產(chǎn)蛋期到就巢期并伴隨著就巢時間的延長,卵巢體積逐漸變小。在產(chǎn)蛋期的卵巢中可以看見等級卵泡生長發(fā)育活躍、層次分明,而進(jìn)入就巢期后等級卵泡停止發(fā)育,甚至發(fā)生閉鎖。產(chǎn)蛋期的卵巢包含了從原始卵泡到排卵前卵泡的各個發(fā)育階段的卵泡結(jié)構(gòu)。產(chǎn)蛋期的卵巢中有6個大的排卵前卵泡(F1-F6),大量的包括原始卵泡、初級卵泡、白色卵泡(LWF、SWF)和黃色卵泡(SYF)的等級前卵泡,以及處于退化階段的排卵后卵泡(POF)。而就巢期卵巢體積較小,呈現(xiàn)萎縮狀,出現(xiàn)大量閉鎖卵泡,卵泡的數(shù)量也越來越少,僅含有一些小卵泡,無突出大卵泡(圖1C-F)。在光學(xué)顯微鏡下觀察卵巢石蠟切片,在NO組的卵巢中,可以看到在正常的卵巢中顆粒層致密緊湊,卵泡內(nèi)膜層與外膜層結(jié)構(gòu)清晰。卵巢中有許多被單層扁平或立方狀的顆粒細(xì)胞包圍的原始卵泡,以及多個被一層立方體顆粒細(xì)胞包圍的初級卵泡(圖1G)。與NO組相比,AO組卵巢中也有許多被單層扁平或立方狀的顆粒細(xì)胞包圍的原始卵泡,但是卵泡數(shù)目較少,初級卵泡顆粒層和膜層之間界限不清,與周圍細(xì)胞分離脫落,胞漿濃縮,顆粒細(xì)胞增生,顆粒層變厚,細(xì)胞密度變稀疏。膜層中,內(nèi)膜層和外膜層從形態(tài)上無法辨認(rèn)(圖1H)。上述研究結(jié)果表明,就巢導(dǎo)致蛋雞卵巢萎縮,卵泡大量閉鎖。
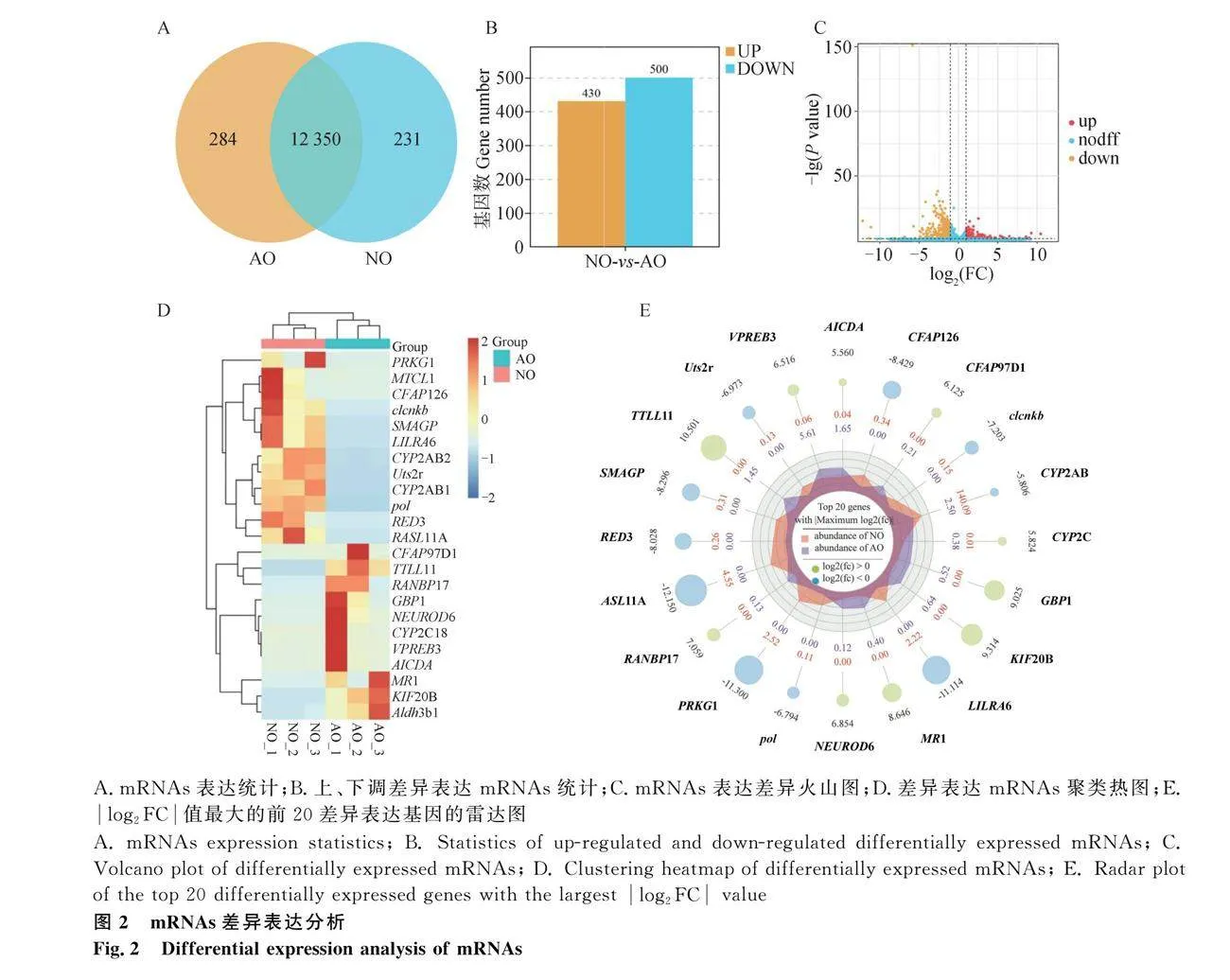
2.2 城口山地雞持續(xù)產(chǎn)蛋期與就巢期卵巢的轉(zhuǎn)錄組分析
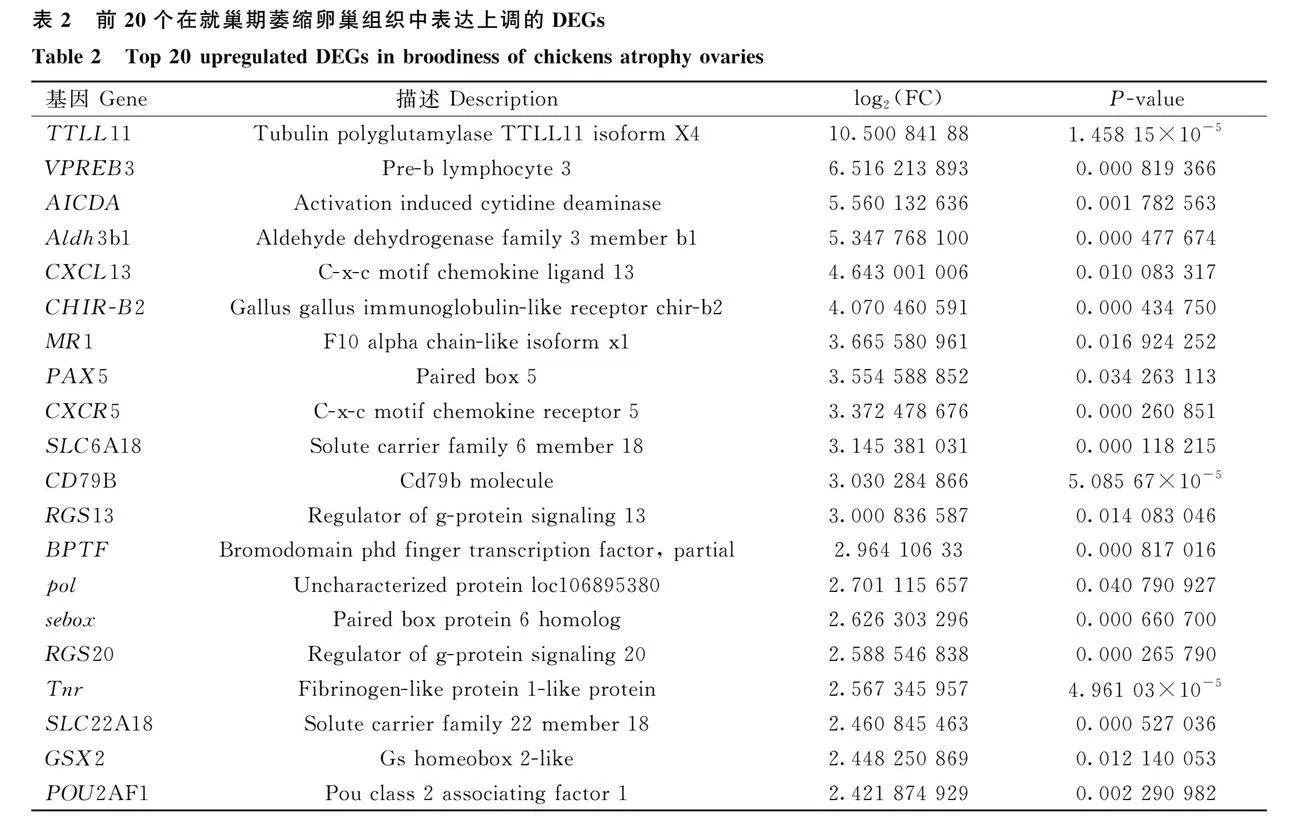
為了進(jìn)一步探究就巢導(dǎo)致卵巢萎縮的關(guān)鍵分子和分子機制,收集了正常產(chǎn)蛋期的卵巢和就巢30天的萎縮卵巢作為樣本,進(jìn)行了基于RNA-seq的轉(zhuǎn)錄組學(xué)分析和基于iTRAQ的蛋白質(zhì)組學(xué)分析。通過轉(zhuǎn)錄組測序產(chǎn)生了6個cDNA文庫,每個樣本平均獲得93 064 205(超過99%)個clean reads,Q30堿基百分比都在92%以上。Pearson相關(guān)性分析表明,就巢組與正常組樣本之間的相關(guān)系數(shù)平方(R2)gt;0.9。以上結(jié)果符合后續(xù)生物信息學(xué)分析的要求。共鑒定出了16 183個mRNAs,包括已知的15 817個mRNAs和預(yù)測的新的366個mRNAs。AO組中共鑒定出了12 634個基因,其中有284個在AO組中特異表達(dá)(圖2A)。AO組與NO組相比,共有930個顯著差異表達(dá)的mRNAs(log2FCgt;1,P≤0.05),其中430個表達(dá)上調(diào)和500個表達(dá)下調(diào)(圖2B-C)。表2和表3分別展示了log2FC最高的20個上調(diào)和下調(diào)差異表達(dá)基因。雷達(dá)圖展示了log2FC值最大的前20個DGEs,以log2FCgt;5,P≤0.05為閾值選擇了23個差異表達(dá)的mRNAs進(jìn)行聚類分析,熱圖進(jìn)一步揭示了不同組間差異表達(dá)模式(圖2D-E)。
2.3 差異表達(dá)基因功能富集分析
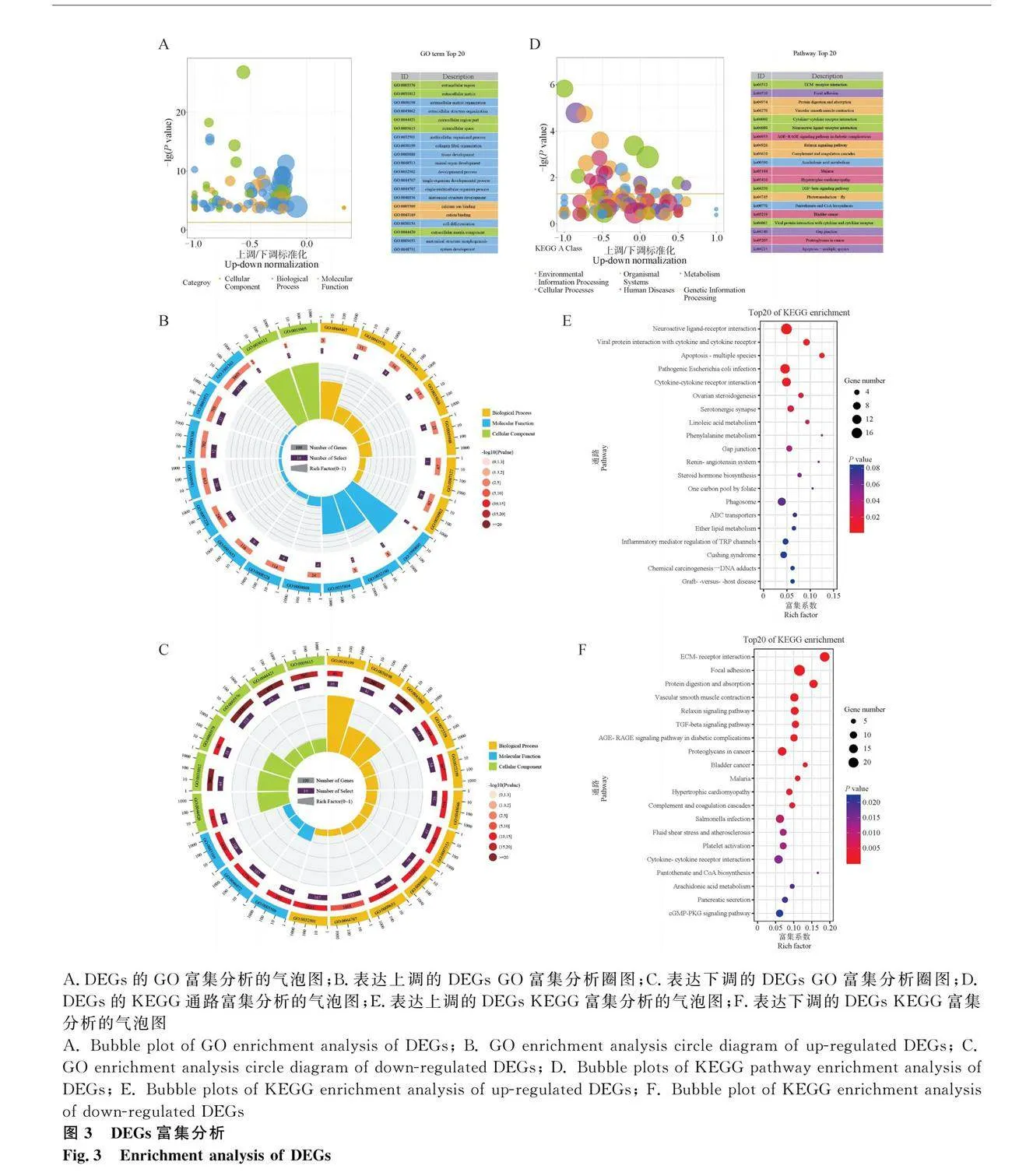
為了進(jìn)一步探究DEGs的功能和相關(guān)通路,對DEGs進(jìn)行了GO注釋和KEGG功能富集分析,以了解發(fā)育卵巢和萎縮卵巢的整體轉(zhuǎn)錄效應(yīng)。GO功能富集分析結(jié)果顯示,DEGs在GO功能富集分析中包含了3個主題,生物過程(BP)、分子功能(MF)和細(xì)胞成分(CC)。22.8%的基因與多細(xì)胞有機體過程、動物器官發(fā)育、離子結(jié)合以及發(fā)育過程等相關(guān)(圖3A)。而且,參與這些功能調(diào)節(jié)的基因61.8%是表達(dá)下調(diào)的。
在表達(dá)上調(diào)的DEGs中,與就巢相關(guān)的高度活化的生物過程包括細(xì)胞-細(xì)胞識別、減數(shù)分裂I、體液免疫反應(yīng)的正調(diào)節(jié)、動物器官發(fā)育、調(diào)節(jié)食欲和卵母細(xì)胞發(fā)育;在分子功能方面,RNA聚合酶II轉(zhuǎn)錄因子活性、序列特異性DNA結(jié)合、核酸結(jié)合轉(zhuǎn)錄因子活性、Netrin受體結(jié)合、有機環(huán)狀化合物結(jié)合與卵巢萎縮的關(guān)聯(lián)度最高;在細(xì)胞組分中,上調(diào)的DEGs主要富集在外部封裝結(jié)構(gòu)、質(zhì)膜的內(nèi)在成分等方面(圖3B)。在表達(dá)下調(diào)的DEGs中,與就巢相關(guān)的高度活化的生物過程主要富集在細(xì)胞外基質(zhì)組織、組織發(fā)育、細(xì)胞黏附、動物器官發(fā)育和細(xì)胞分化;在分子功能方面,就巢產(chǎn)生的變化與金屬離子結(jié)合、生長因子結(jié)合、胰島素樣生長因子結(jié)合最相關(guān);在細(xì)胞組分中,下調(diào)的DEGs主要富集在細(xì)胞外區(qū)域、質(zhì)膜、抑制素復(fù)合物等方面(圖3C)。
KEGG功能富集分析結(jié)果顯示,與就巢相關(guān)的通路包括ECM受體反應(yīng)、黏著斑、蛋白質(zhì)消化吸收、血管平滑肌收縮等(圖3D)。其中,表達(dá)上調(diào)的DEGs顯著富集到了神經(jīng)活性配體-受體相互作用、細(xì)胞凋亡-多個物種、細(xì)胞因子-細(xì)胞因子受體相互作用和卵巢類固醇生成等通路(圖3E)。表達(dá)下調(diào)的DEGs顯著富集到了ECM受體相互作用、黏著、蛋白質(zhì)消化和吸收、松弛素信號通路、TGF-β 信號通路和雌激素信號通路等信號通路(圖3F)。GSEA分析顯示,細(xì)胞周期、卵母細(xì)胞減數(shù)分裂、黃體酮介導(dǎo)的卵母細(xì)胞成熟、雌激素信號通路、Notch信號通路、胰島素分泌、催產(chǎn)素信號通路在卵巢組織中顯著上調(diào)。值得注意的是,PI3K-Akt信號通路、Notch信號通路、p53信號通路等與細(xì)胞增殖、分化和轉(zhuǎn)移,細(xì)胞生存與死亡,調(diào)控細(xì)胞命運,參與炎癥反應(yīng)的重要信號轉(zhuǎn)導(dǎo)途徑相關(guān)的類別也顯著富集。上述結(jié)果為從mRNA水平探討就巢導(dǎo)致卵巢萎縮的關(guān)鍵調(diào)控機制提供了重要的背景信息。
圖4A簡要展示了篩選就巢期卵巢萎縮相關(guān)的23個關(guān)鍵PI3K-Akt信號通路調(diào)節(jié)因子的試驗方案。GSEA結(jié)果顯示,在AO組的卵巢組織中,細(xì)胞生長和死亡相關(guān)通路都被激活(圖4B)。進(jìn)一步,重點評估了參與PI3K-Akt信號通路(圖4C)的基因,其中包括了6個在AO組中上調(diào)的DEGs:FGFR4、LPAR5、CCNE1、NGFR、SGK2、EIF4E1B和17個在AO組下調(diào)的DEGs:OSMR、COMP、ITGA8、VTN、COL6A2、COL6A1、PDGFD、THBS2、ITGA10、CHAD、TNC、COL1A2、FN1、COL1A1、THBS1、COL4A2、COL4A1。在熱圖中顯示了這23個基因的FPKM水平(圖4D)。蛋白質(zhì)-蛋白質(zhì)相互作用網(wǎng)絡(luò)(PPI)顯示,這23個PI3K-Akt信號通路中的調(diào)節(jié)因子可以通過7種方式相互作用,包括來自精選數(shù)據(jù)庫和試驗確定的已知相互作用,基因鄰里、基因融合和基因共生的預(yù)測的交互,以及文本挖掘、共表達(dá)和蛋白質(zhì)同源性(圖4E)。繼續(xù)分析了參與的BPs和富集通路。結(jié)果顯示細(xì)胞通訊、細(xì)胞黏附、細(xì)胞表面受體信號通路、細(xì)胞增殖、細(xì)胞遷移和細(xì)胞過程的正向調(diào)節(jié)是富集最多的BPs(圖4F)。并且,這23個基因顯著富集于肌動蛋白細(xì)胞骨架的調(diào)節(jié)、Rap1信號通路和Ras信號通路等與細(xì)胞黏附和細(xì)胞連接的信息與調(diào)控,細(xì)胞遷移、極化,細(xì)胞增殖和存活相關(guān)顯著的通路中(圖4G)。
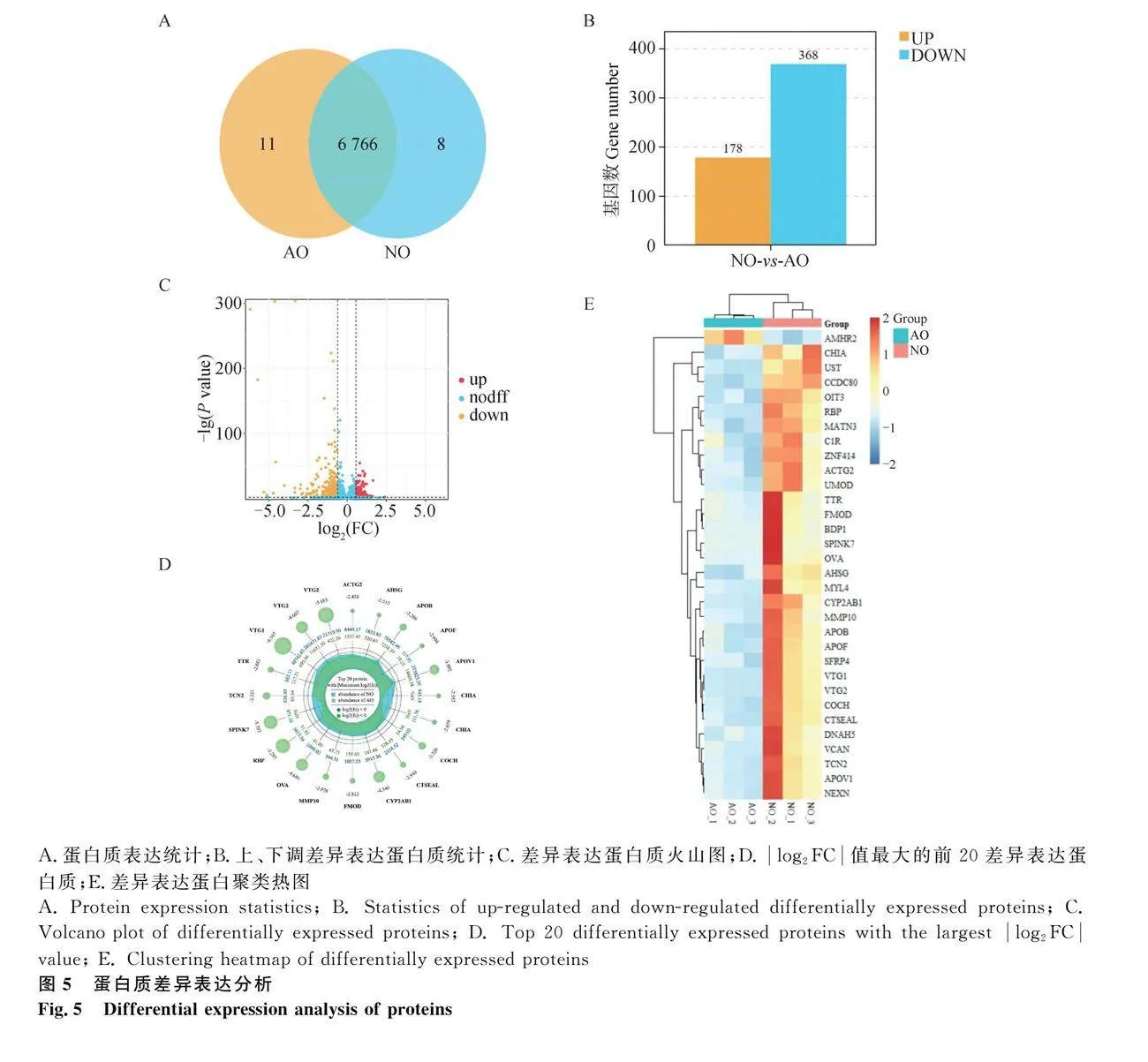
2.4 城口山地雞持續(xù)產(chǎn)蛋期卵巢與就巢期卵巢的蛋白組分析
以NO組為對照組,使用iTRAO對AO組進(jìn)行蛋白質(zhì)測序,然后進(jìn)行LC-MS/MS分析。總共鑒定出91 569個光譜,80 267個肽段、6 703個蛋白質(zhì)組和6 785個蛋白質(zhì)。鑒定的蛋白質(zhì)大部分含有11個以上的肽段。Pearson相關(guān)性分析顯示,AO組與NO組樣本之間的相關(guān)系數(shù)平方(R2)gt;0.9。以上結(jié)果符合后續(xù)生物信息學(xué)分析的要求。與NO組相比,AO組共鑒定出了6 777個蛋白質(zhì),其中有11個在就巢組中特異性表達(dá)(圖5A)。AO組與NO組相比,共鑒定出546個顯著差異表達(dá)的蛋白質(zhì)(Fold Changegt;1.5,P≤0.05),其中表達(dá)上調(diào)的有178個,表達(dá)下調(diào)的有368個(圖5B)。繪制了火山圖來展示DEPs的詳細(xì)情況(圖5C)。雷達(dá)圖展示了log2FC值最大的前20個DEPs(圖5D),熱圖選擇了30個DEPs進(jìn)行聚類分析,進(jìn)一步揭示了不同組間差異表達(dá)模式(圖5E)。
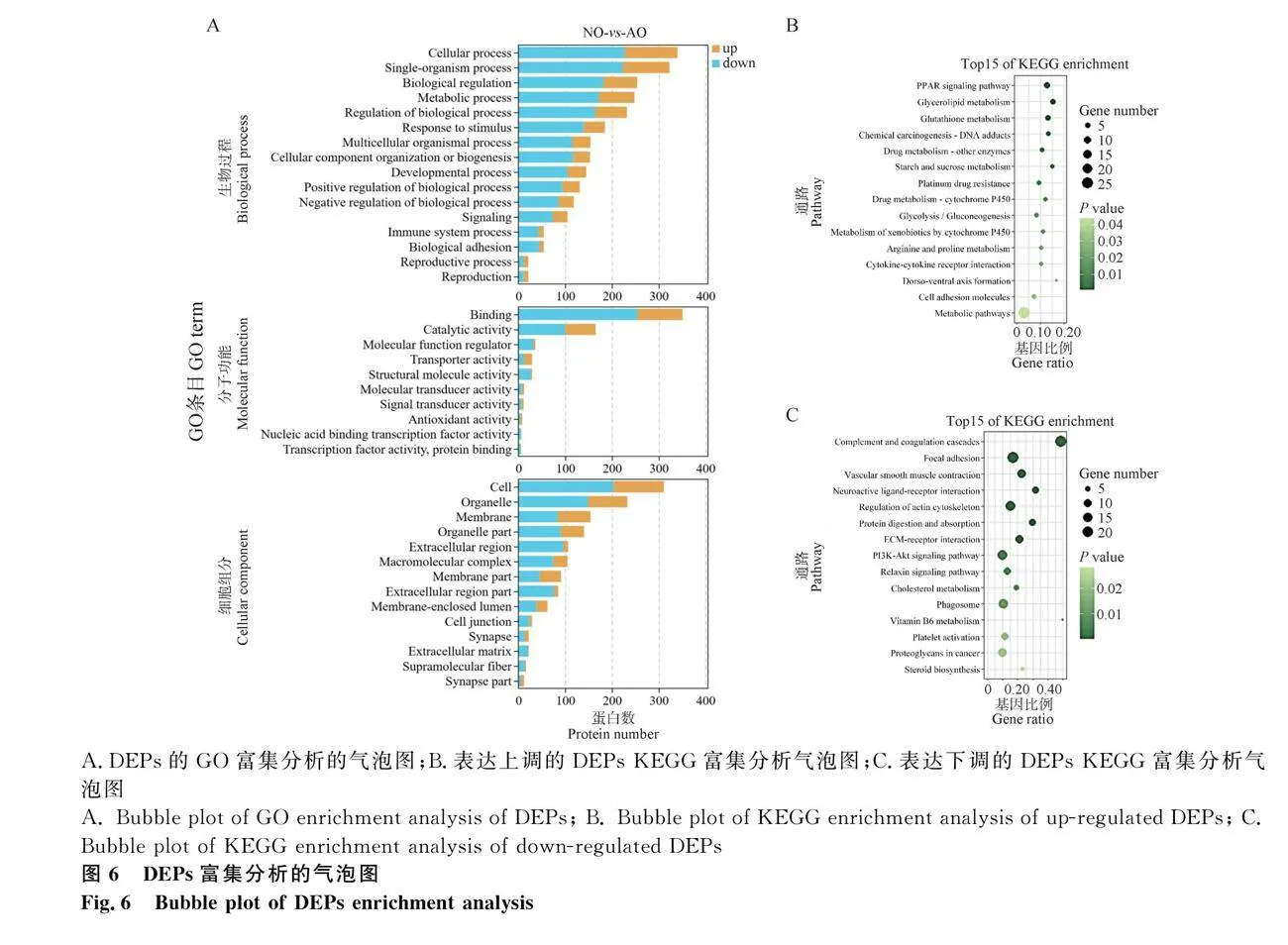
為了進(jìn)一步分析DEPs的生物學(xué)功能,對DEPs進(jìn)行了GO注釋和KEGG通路富集分析。GO富集分析結(jié)果顯示(圖6A),在GO二級分類條目中的生物過程類別中,22.6%的DEPs主要涉及細(xì)胞過程、代謝過程、生物過程調(diào)控、信號及生物調(diào)控功能,且這些DEPs中69.4%是表達(dá)下調(diào)的。值得注意的是,有幾個DEPs參與了免疫系統(tǒng)過程、生物黏附、繁殖過程、生長和細(xì)胞殺傷;在細(xì)胞成分類別中,大量的DEPs涉及細(xì)胞、細(xì)胞部分、細(xì)胞器、細(xì)胞外區(qū)域、大分子復(fù)合物和膜。值得注意的是,少數(shù)DEPs涉及細(xì)胞外基質(zhì)、細(xì)胞連接和超分子纖維;在分子功能方面,DEPs主要涉及到結(jié)合和催化活性。此外還觀察到,下調(diào)表達(dá)的DEPs數(shù)量顯著多于上調(diào)表達(dá)的DEPs,尤其是富集在與蛋白質(zhì)結(jié)合、催化活性、分子功能調(diào)節(jié)劑、結(jié)構(gòu)分子和轉(zhuǎn)錄因子活性相關(guān)的GO條目。這說明通過雞的就巢行為可能通過抑制細(xì)胞與大分子物質(zhì)結(jié)合來降低卵巢的代謝過程,從而導(dǎo)致了卵巢的萎縮。然而,在轉(zhuǎn)運蛋白條目中,上調(diào)的DEPs的數(shù)量顯著高于下調(diào)DEPs的數(shù)量,這表明依靠特異轉(zhuǎn)運蛋白的活性在生物膜內(nèi)外的化學(xué)物質(zhì)以及信號交換過程中,抑制細(xì)胞代謝、信號轉(zhuǎn)導(dǎo)等廣泛的細(xì)胞活動。上述結(jié)果與轉(zhuǎn)錄組分析的結(jié)果一致。
KEGG富集分析結(jié)果顯示(圖6B),上調(diào)的DEPs主要與代謝通路密切相關(guān),如代謝途徑、甘油代謝、糖酵解/糖異生和PPAR信號通路。其中,一些信號通路與機體的能量來源密切相關(guān),特別是甘油代謝和糖酵解/糖異生信號通路。此外,幾個DEPs與谷胱甘肽代謝和PPAR信號通路有關(guān),這些通路主要參與細(xì)胞保護(hù)、細(xì)胞增殖和調(diào)節(jié)脂質(zhì)氧化,促進(jìn)脂肪細(xì)胞分化以增強血糖攝取,維持代謝穩(wěn)態(tài)和炎癥基因的表達(dá)(圖6B)。下調(diào)的DEPs主要與黏著、肌動蛋白細(xì)胞骨架的調(diào)節(jié)、PI3K-Akt信號通路、吞噬體和松弛素信號通路有關(guān)。其中,黏著和肌動蛋白細(xì)胞骨架的調(diào)節(jié)信號通路在細(xì)胞與外部基質(zhì)之間對于細(xì)胞凋亡、增殖、運動(分裂、遷移、收縮和黏附)、分化等過程都具有重要作用,特別是PI3K-Akt信號通路對許多細(xì)胞基本進(jìn)程的調(diào)控,包括細(xì)胞生長、轉(zhuǎn)錄、翻譯、細(xì)胞增殖、細(xì)胞運動及糖原代謝中調(diào)控關(guān)鍵細(xì)胞反應(yīng)。此外,一些DEPs與血管平滑肌收縮、ECM與受體的相互作用和核糖體有關(guān),同樣在細(xì)胞遷移、增殖、分化和信號轉(zhuǎn)導(dǎo)等細(xì)胞活動中發(fā)揮重要作用(圖6C)。這些數(shù)據(jù)為在蛋白質(zhì)水平上闡明就巢與卵巢萎縮之間的調(diào)控機制提供了重要的背景信息。
2.5 轉(zhuǎn)錄組學(xué)和蛋白質(zhì)組學(xué)聯(lián)合分析
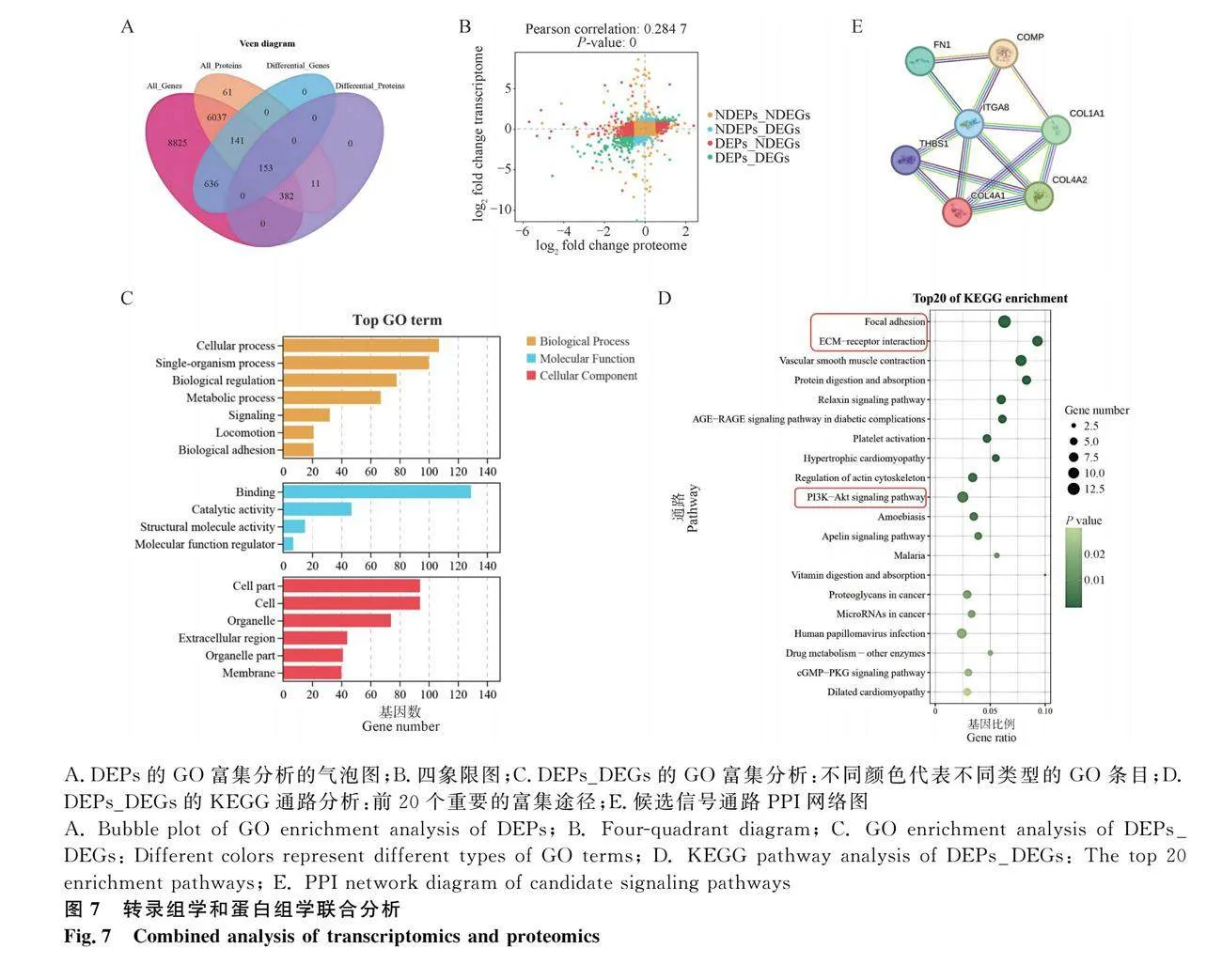
為了挖掘轉(zhuǎn)錄組和蛋白組的互補性和整合性,分析了DEGs和DEPs之間的相關(guān)性。大量轉(zhuǎn)錄本編碼的蛋白質(zhì)覆蓋率很高,其中,顯示了930個DEGs和546個DEPs,共包括153個在mRNAs和蛋白質(zhì)水平均有差異表達(dá)的基因(圖7A)。此外,為了研究蛋白質(zhì)水平的變化是否與相應(yīng)轉(zhuǎn)錄本的變化相關(guān),繪制了四象限圖分析了相應(yīng)mRNAs與蛋白質(zhì)比率的分布。大多數(shù)mRNAs與蛋白質(zhì)的比值集中在圖的中心位置,只有部分基因和蛋白質(zhì)的表達(dá)水平不一致,大多數(shù)表達(dá)差異顯著基因的mRNAs與蛋白質(zhì)的比值集中在圖的第二、三象限,表達(dá)水平是一致的(圖7B)。對表現(xiàn)出相同表達(dá)趨勢的mRNAs和蛋白質(zhì)進(jìn)行GO注釋和KEGG通路富集分析,GO分析表明,這些基因主要富集于細(xì)胞、細(xì)胞器、信號、代謝、結(jié)合、生物調(diào)控和催化活性(圖7C)。通過KEGG通路富集分析,它們主要參與黏著斑、ECM受體反應(yīng)、PI3K-Akt信號通路(圖7D)。值得注意的是,這3個通路都是典型的與細(xì)胞遷移、增殖、分化和信號轉(zhuǎn)導(dǎo)等廣泛的細(xì)胞活動顯著相關(guān)的通路。進(jìn)一步分析了這3個通路調(diào)控網(wǎng)絡(luò)中富集到的基因,表4中顯示了富集到3個候選通路中的基因和蛋白質(zhì),其中COMP、FN1、ITGA8、THBS1、COL4A2、COL4A1、COL1A1為3個通路中共有的基因,推測它們可能在蛋雞就巢期間卵巢結(jié)構(gòu)和功能的變化中起著重要的作用,可以作為家禽就巢性的候選基因。利用STRING數(shù)據(jù)庫(https://cn.string-db.org/)進(jìn)一步分析,闡明蛋白-蛋白相互作用(PPI)結(jié)果(圖7E)。
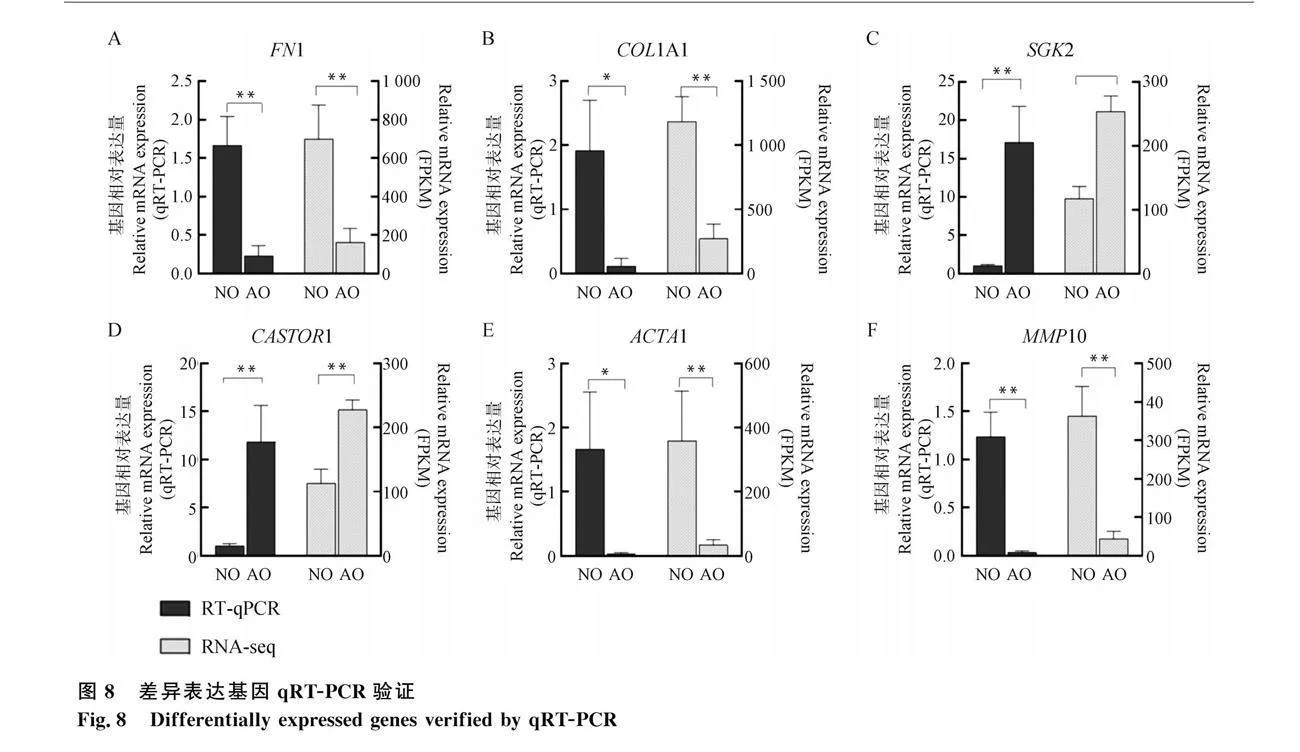
2.6 測序結(jié)果驗證
隨機選擇在AO組上調(diào)或下調(diào)的6個基因,F(xiàn)N1、COL1A1、SGK2、CASTOR1、ACTA1和MMP10在NO組和AO組進(jìn)行qRT-PCR驗證。結(jié)果顯示RT-qPCR和RNA-seq結(jié)果一致,6個差異表達(dá)基因的相對表達(dá)量均有顯著差異(Plt;0.05或Plt;0.01)(圖8),結(jié)果驗證了測序數(shù)據(jù)的可靠性。
3 討 論
本研究探討了就巢導(dǎo)致卵巢萎縮的關(guān)鍵調(diào)控機制,特別是就巢如何調(diào)控卵巢萎縮導(dǎo)致卵泡閉鎖。首先,觀察了就巢組萎縮卵巢的HE染色切片,與產(chǎn)蛋組正常卵巢相比,可以看出卵泡中顆粒層和膜細(xì)胞有明顯的減少,卵巢發(fā)生萎縮,卵泡大量閉鎖,這與He等[23]的研究結(jié)果是一致的。然后,收集了正常卵巢和萎縮卵巢樣本,進(jìn)行了基于RNA-seq的轉(zhuǎn)錄組學(xué)分析和基于iTRAO的蛋白質(zhì)組學(xué)分析,篩選出大量的DEGs和DEPs,并進(jìn)一步進(jìn)行了關(guān)聯(lián)分析,不僅代表了mRNAs表達(dá)水平,也有展現(xiàn)翻譯后的特征。轉(zhuǎn)錄組和蛋白組的聯(lián)合分析將為探究就巢與卵巢萎縮之間的內(nèi)在調(diào)控機制提供一個全面的視角。
禽類有超過95%的卵泡會發(fā)生內(nèi)源性的閉鎖,當(dāng)卵泡發(fā)育到排卵前卵泡階段時,一般不會發(fā)生內(nèi)源性的閉鎖,而其他階段的卵泡都會發(fā)生不同程度的閉鎖。Tilly等[24]發(fā)現(xiàn),雞卵泡閉鎖存在顆粒細(xì)胞凋亡現(xiàn)象。也有報道提出,家鵝就巢導(dǎo)致卵泡閉鎖,且與顆粒細(xì)胞凋亡相關(guān)[25-26]。細(xì)胞凋亡是維持生物體穩(wěn)定的基本生理機制。在家禽中,卵泡閉鎖發(fā)生在卵泡發(fā)育的各個階段。在正常生長的卵泡中,顆粒細(xì)胞的分裂、增殖和凋亡也是并存的[27],其中顆粒細(xì)胞的凋亡會直接導(dǎo)致卵泡發(fā)生閉鎖[28]。在本研究中觀察到與細(xì)胞凋亡通路相關(guān)的基因,抗凋亡基因MAPK10、NGFR、BIRC7和BIRC均表達(dá)上調(diào),而SEPTIN4表達(dá)下調(diào)。SEPTIN4是一種定位于線粒體的蛋白,主要通過與X連鎖凋亡抑制因子(x-linked inhibitors of apoptosis,XIAP)結(jié)合促進(jìn)細(xì)胞凋亡[29]。分裂原激活蛋白激酶(mitogen-activated protein kinase,MAPK)通過轉(zhuǎn)錄和轉(zhuǎn)錄后機制調(diào)節(jié)細(xì)胞凋亡。MAPK可將細(xì)胞外信息傳遞至細(xì)胞核中,在生理及病理過程中發(fā)揮重要的作用。MAPK在細(xì)胞死亡(促凋亡和抗凋亡)中起哪種作用取決于不同刺激激活靶點的多樣性,或特定刺激作用的分子環(huán)境。然而,即使在受到特定刺激的某一細(xì)胞類型中,MAPK的激活也可能是促凋亡和/或抗凋亡[30]。因此,推測在就巢期間MAPK與促進(jìn)卵泡閉鎖的細(xì)胞凋亡和促進(jìn)卵泡發(fā)育的抗凋亡的不同膜受體結(jié)合的現(xiàn)象同時存在。不過,這還需要進(jìn)一步的試驗驗證。
然而,細(xì)胞凋亡并不是導(dǎo)致卵泡閉鎖的唯一因素。HPG軸的協(xié)調(diào)活動通過幾種生殖內(nèi)分泌激素和卵巢內(nèi)生長因子的啟動在維持卵泡生長和排卵中發(fā)揮著至關(guān)重要的作用。對雞的分級前、排卵前和排卵后卵泡進(jìn)行轉(zhuǎn)錄組學(xué)分析時,也篩選出了差異表達(dá)的凋亡和類固醇生物合成相關(guān)的基因[31]。同樣,在本研究中也篩選出了一些與卵巢類固醇生成信號通路相關(guān)的基因,如CYP1A1、CYP2J2、HSD17B7和BMP15表達(dá)顯著上調(diào),而HSD3B1表達(dá)下調(diào)。細(xì)胞色素P450(CYP)超家族包含的基因編碼的酶反應(yīng)可以在多種方面發(fā)揮作用,包括膽固醇代謝和膽汁酸生物合成、類固醇合成與代謝、維生素D3的合成與代謝等,也包括無數(shù)涉及酶反應(yīng)的重要生命過程。許多CYP酶對內(nèi)源性和外源性底物的代謝會導(dǎo)致活性氧代謝物(ROM)介導(dǎo)的氧化應(yīng)激,而氧化應(yīng)激是誘發(fā)細(xì)胞凋亡的一個主要信號[32]。而CYP2J2(CYP家族2亞家族J成員2)或其代謝物會通過增加MAPK和PI3K/Akt通路的活性,以及增強表皮生長因子受體(epithelial growth factor receptor,EGFR)的磷酸化,促進(jìn)癌細(xì)胞明顯增殖。此外,CYP2J2還通過上調(diào)抗凋亡蛋白Bcl-2和Bcl-xL以及下調(diào)促凋亡蛋白Bax來抑制癌細(xì)胞凋亡[33]。而且,許多CYP基因的突變導(dǎo)致先天性代謝缺陷,并導(dǎo)致許多臨床相關(guān)疾病。但在就巢過程中類固醇生成信號通路相關(guān)的基因促進(jìn)或抑制細(xì)胞凋亡并可能進(jìn)一步導(dǎo)致卵泡閉鎖的作用機制目前仍不清楚,這些都為今后的研究提供了方向。
此外,在本研究中觀察到有多個基因及其編碼的蛋白質(zhì)顯著富集到了ECM受體反應(yīng)信號通路,包括COMP、FN1、ITGA8、THBS1、SDC1、COL4A2、COL4A1、TNC和COL1A1,但這些基因均表達(dá)下調(diào)。ECM作為一種細(xì)胞分泌組裝的蛋白質(zhì)和多糖的復(fù)雜網(wǎng)絡(luò)結(jié)構(gòu),不僅能為細(xì)胞提供附著位點和結(jié)構(gòu)支持,還可以為調(diào)節(jié)細(xì)胞活性和器官、組織功能提供重要的生物信息[34]。禽類卵巢中卵泡的生長、發(fā)育、排卵和閉鎖不僅伴隨著細(xì)胞生理學(xué)的變化,而且還伴隨ECM的廣泛重組。研究表明,ECM成分提供了組織特異性的支架,影響基本的細(xì)胞過程,如增殖、分化和凋亡等。卵泡發(fā)育過程中ECM的全面變化需要一系列的蛋白水解過程,對于正常的卵泡發(fā)育、功能、閉鎖和退化至關(guān)重要[35]。有研究指出,卵泡發(fā)育或者閉鎖期間的ECM周轉(zhuǎn)在很大程度上由基質(zhì)蛋白控制,除了特異性降解ECM成分和激活其他基質(zhì)金屬蛋白酶外,它們還可以通過釋放大量信號蛋白影響多種信號通路,在正常的生理和病理過程中調(diào)節(jié)細(xì)胞生物學(xué),其中包括生長因子、細(xì)胞因子、鈣黏蛋白E、Fas配體和腫瘤壞死因子等[36]。肌腱蛋白C(tenascin C,TNC)是ECM分泌的具有中介和整合子功能的基質(zhì)細(xì)胞蛋白,可以正向或負(fù)向地影響信號通路[37]。TNC在發(fā)育過程中高度表達(dá)于ECM中,參與了發(fā)育和組織重塑過程中的形態(tài)變化,以及細(xì)胞遷移、增殖分化、凋亡與細(xì)胞間的黏附和信號傳遞[38]。血小板源生長因子(plateletderived growth factor,PDGF)可以通過PI3K-AKT信號通路調(diào)節(jié)TNC的表達(dá),導(dǎo)致轉(zhuǎn)錄因子SP、ETS1和ETS2與特異性啟動子結(jié)合[39],參與細(xì)胞增殖、分化和死亡的調(diào)控。FN1基因編碼纖維連接蛋白也是ECM的主要成分,在細(xì)胞表面和細(xì)胞外基質(zhì)中以二聚體或多聚體形式存在,參與了多個生物過程,主要作用是支持細(xì)胞黏附,也參與細(xì)胞骨架組織、細(xì)胞遷移和重要的生理過程[40]。綜上所述,推測就巢可能會導(dǎo)致多種轉(zhuǎn)錄因子活化降低或者存在細(xì)胞因子介導(dǎo)的蛋白質(zhì)水解,降低ECM蛋白的表達(dá),從而進(jìn)一步抑制了細(xì)胞內(nèi)信號通路在卵泡發(fā)育過程中調(diào)控細(xì)胞遷移、增殖分化的信號轉(zhuǎn)導(dǎo)。
由生長因子調(diào)控的PI3K-Akt信號網(wǎng)絡(luò)無處不在,它通過直接調(diào)控營養(yǎng)物質(zhì)轉(zhuǎn)運體和代謝酶,或調(diào)控轉(zhuǎn)錄因子來調(diào)控代謝途徑關(guān)鍵組分的表達(dá),從而對細(xì)胞代謝產(chǎn)生多種下游影響。如Cui等[41]的研究表明,蛋雞就巢期萎縮卵巢高表達(dá)miR-204通過PI3K/Akt信號通路促進(jìn)顆粒細(xì)胞凋亡。在本研究中,許多富集于PI3K-Akt信號通路的基因及其對應(yīng)編碼的蛋白質(zhì)被上調(diào),包括FGFR4、LPAR5、CCNE1、NGFR、SGK2和EIF4E1B。其中,成纖維細(xì)胞生長因子受體4(fibroblast growth factor receptor 4,F(xiàn)GFR4)是細(xì)胞膜上高度保守的酪氨酸激酶受體(receptor tyrosine kinases,RTKs)家族的成員,通過與配體結(jié)合并激活下游信號通路參與調(diào)節(jié)細(xì)胞增殖、分化和生存等多種生物學(xué)過程[42]。FGFR4在妊娠小鼠的卵巢中有表達(dá),并且在卵泡發(fā)育不同階段的卵巢卵泡中均有表達(dá)[43],可能介導(dǎo)從卵泡發(fā)生的最早階段開始的卵母細(xì)胞和顆粒細(xì)胞之間的旁分泌相互作用。溶血磷脂酸受體5(lysophosphatidic acid receptor 5,LPAR5)是介導(dǎo)細(xì)胞內(nèi)對溶血磷脂酸(lysophosphatidic acid,LPA)反應(yīng)的G蛋白偶聯(lián)跨膜溶血磷脂酸(LPA)受體之一,具有多種生物學(xué)功能,與細(xì)胞增殖、形態(tài)發(fā)生、分化和防止細(xì)胞凋亡有關(guān),并參與蛋白質(zhì)水解和介導(dǎo)細(xì)胞內(nèi)對LPA的反應(yīng)[44]。神經(jīng)生長因子受體(nerve growth factor receptor,NGFR)不具有內(nèi)在的酶活性,它的信號傳遞依賴于細(xì)胞內(nèi)結(jié)合蛋白的募集,從而激活不同的信號通路,通過MAPK、PI3K/AKT典型信號途徑傳遞細(xì)胞內(nèi)信號,介導(dǎo)細(xì)胞存活[45]。綜上所述,蛋雞就巢可激活多種細(xì)胞因子受體,與配體結(jié)合激活PI3K-Akt信號轉(zhuǎn)導(dǎo)通路,維持卵泡顆粒細(xì)胞的生存或者死亡狀態(tài),然而,具體的機制還需要進(jìn)一步的試驗驗證。
4 結(jié) 論
本研究獲得了就巢導(dǎo)致卵巢萎縮的基于RNA-seq的轉(zhuǎn)錄組學(xué)圖譜和基于iTRAQ的蛋白組學(xué)圖譜,篩選出了重要的DEGs和DEPs。通過對這些DEGs和DEPs進(jìn)行功能富集分析,發(fā)現(xiàn)就巢期卵巢組織中多種細(xì)胞因子受體與配體介導(dǎo)部分功能蛋白的水解,從而影響信號轉(zhuǎn)導(dǎo)的細(xì)胞環(huán)境,進(jìn)一步影響下游細(xì)胞活動。同時,篩選出了7個蛋雞就巢性的候選基因COMP、FN1、ITGA8、THBS1、COL4A2、COL4A1、COL1A1。本研究為深入了解蛋雞就巢期卵巢萎縮的調(diào)控機制提供了理論基礎(chǔ),為家禽就巢性遺傳改良和分子育種提供有效參考。
參考文獻(xiàn)(References):
[1] 李 俊,李建偉,楊 蓉,等.家禽就巢性影響因素的研究進(jìn)展[J].中國畜禽種業(yè),2023,19(8):91-95.
LI J,LI J W,YANG R,et al.Research progress on factors affecting broodiness in poultry[J].The Chinese Livestock and Poultry Breeding,2023,19(8):91-95.(in Chinese)
[2] ROMANOV M N,TALBOT R T,WILSON P W,et al.Genetic control of incubation behavior in the domestic hen[J].Poult Sci,2002,81(7):928-931.
[3] 劉安懷,徐茂森,張 旭,等.產(chǎn)蛋期和就巢期皖西白鵝卵巢組織結(jié)構(gòu)、血清生化指標(biāo)和激素水平比較研究[J].安徽科技學(xué)院學(xué)報,2023,37(2):1-5.
LIU A H,XU M S,ZHANG X,et al.Effects of brooding behavior on ovarian tissue structure,blood biochemical indexes and hormone levels of Wanxi white geese[J].Journal of Anhui Science and Technology University,2023,37(2):1-5.(in Chinese)
[4] 王思博,楊 季,馬渭青,等.鵝就巢期卵巢特征及其內(nèi)分泌機制的研究進(jìn)展[J].中國畜牧雜志,2020,56(3):33-37.
WANG S B,YANG J,MA W Q,et al.Characteristics of ovary and its endocrine mechanism in geese during nesting[J].Chinese Journal of Animal Science,2020,56(3):33-37.(in Chinese)
[5] ZHAO J B,PAN H B,LIU Y,et al.Interacting networks of the hypothalamic-pituitary-ovarian axis regulate layer hens performance[J].Genes (Basel),2023,14(1):141.
[6] YU J,LOU Y P,HE K,et al.Goose broodiness is involved in granulosa cell autophagy and homeostatic imbalance of follicular hormones[J].Poult Sci,2016,95(5):1156-1164.
[7] LIU L B,XIAO Q H,GILBERT E R,et al.Whole-transcriptome analysis of atrophic ovaries in broody chickens reveals regulatory pathways associated with proliferation and apoptosis[J].Sci Rep,2018,8(1):7231.
[8] JOHNSON A L,BRIDGHAM J T,WITTY J P,et al.Expression of bcl-2 and nr-13 in hen ovarian follicles during development[J].Biol Reprod,1997,57(5):1096-1103.
[9] XU G Q,DONG Y Y Y,WANG Z,et al.Melatonin Attenuates Oxidative Stress-induced apoptosis of bovine ovarian granulosa cells by promoting mitophagy via SIRT1/FoxO1 signaling pathway[J].Int J Mol Sci,2023,24(16):12854.
[10] GARRETT W M,GUTHRIE H D.Steroidogenic enzyme expression during preovulatory follicle maturation in pigs[J].Biol Reprod,1997,56(6):1424-1431.
[11] SHEN X,BAI X,LUO C L,et al.Quantitative proteomic analysis of chicken serum reveals key proteins affecting follicle development during reproductive phase transitions[J].Poult Sci,2021,100(1):325-333.
[12] JIANG R S,XU G Y,ZHANG X Q,et al.Association of polymorphisms for prolactin and prolactin receptor genes with broody traits in chickens[J].Poult Sci,2005,84(6):839-845.
[13] PONSUKSILI S,TRAKOOLJUL N,HADLICH F,et al.Genetic regulation of liver metabolites and transcripts linking to biochemical-clinical parameters[J].Front Genet,2019,10:348.
[14] JIANG S S,ZHANG G H,MIAO J,et al.Transcriptome and metabolome analyses provide insight into the glucose-induced adipogenesis in porcine adipocytes[J].Curr Issues Mol Biol,2024,46(3):2027-2042.
[15] 王亞鑫,王 璟,田學(xué)凱,等.多組學(xué)技術(shù)在畜禽重要經(jīng)濟(jì)性狀研究中的應(yīng)用[J].畜牧獸醫(yī)學(xué)報,2024,55(5):1842-1853.
WANG Y X,WANG J,TIAN X K,et al.Application of multi-omics technology in the study of important economic traits of livestock and poultry[J].Acta Veterinaria et Zootechnica Sinica,2024,55(5):1842-1853.(in Chinese)
[16] HUANG X,ZHANG H Y,CAO H Y,et al.Transcriptomics and metabolomics analysis of the ovaries of high and low egg production chickens[J].Animals (Basel),2022,12(16):2010.
[17] WANG L Y,ZHANG Y W,ZHANG B,et al.Candidate gene screening for lipid deposition using combined transcriptomic and proteomic data from Nanyang black pigs[J].BMC Genomics,2021,22(1):441.
[18] TAO Z,SONG W,ZHU C,et al.Comparative transcriptomic analysis of high and low egg-producing duck ovaries[J].Poult Sci,2017,96(12):4378-4388.
[19] SHEN X,BAI X,XU J,et al.Transcriptome sequencing reveals genetic mechanisms underlying the transition between the laying and brooding phases and gene expression changes associated with divergent reproductive phenotypes in chickens[J].Mol Biol Rep,2016,43(9):977-989.
[20] KOVCS J,F(xiàn)ORG V,PCZELY P.The fine structure of the follicular cells in growing and atretic ovarian follicles of the domestic goose[J].Cell Tissue Res,1992,267(3):561-569.
[21] YU J,LOU Y P,ZHAO A Y.Transcriptome analysis of follicles reveals the importance of autophagy and hormones in regulating broodiness of Zhedong white goose[J].Sci Rep,2016,6(1):36877.
[22] POWERS R K,GOODSPEED A,PIELKE-LOMBARDO H,et al.GSEA-InContext:identifying novel and common patterns in expression experiments[J].Bioinformatics,2018,34(13):i555-i564.
[23] HE H R,LI D M,TIAN Y T,et al.miRNA sequencing analysis of healthy and atretic follicles of chickens revealed that miR-30a-5p inhibits granulosa cell death via targeting Beclin1[J].J Anim Sci Biotechnol,2022,13(1):55.
[24] TILLY J L,KOWALSKI K I,JOHNSON A L,et al.Involvement of apoptosis in ovarian follicular atresia and postovulatory regression[J].Endocrinology,1991,129(5):2799-2801.
[25] CHOI J,JO M,LEE E,et al.AKT is involved in granulosa cell autophagy regulation via mTOR signaling during rat follicular development and atresia[J].Reproduction,2014,147(1):73-80.
[26] KAIPIA A,HSUEH A J W.Regulation of ovarian follicle atresia[J].Annu Rev Physiol,1997,59:349-363.
[27] 茹 盟,曾文惠,彭劍玲,等.蛋雞卵泡發(fā)育及其表觀遺傳調(diào)控機制研究進(jìn)展[J].畜牧獸醫(yī)學(xué)報,2023,54(9):3613-3622.
RU M,ZENG W H,PENG J L,et al.Research progress on follicles development of hens and its epigenetic regulatory mechanism[J].Acta Veterinaria et Zootechnica Sinica,2023,54(9):3613-3622.(in Chinese)
[28] 鄭 言,曹中贊,馬 賁,等.氧化應(yīng)激對家禽卵泡顆粒細(xì)胞凋亡和自噬的影響及機制研究進(jìn)展[J].中國畜牧雜志,2023,59(9):110-115.
ZHENG Y,CAO Z Z,MA B,et al.Research progress on the effect of oxidative stress on apoptosis and autophagy of poultry follicular granulosa cells and its mechanism[J].Chinese Journal of Animal Science,2023,59(9):110-115.(in Chinese)
[29] ZHANG Y,ZHANG N J,ZOU Y M,et al.Deacetylation of Septin4 by SIRT2 (silent mating type information regulation 2 Homolog-2) mitigates damaging of hypertensive nephropathy[J].Circ Res,2023,132(5):601-624.
[30] YUE J C,LPEZ J M.Understanding MAPK signaling pathways in apoptosis[J].Int J Mol Sci,2020,21(7):2346.
[31] ZHU G Y,MAO Y,ZHOU W D,et al.Dynamic changes in the follicular transcriptome and promoter DNA methylation pattern of steroidogenic genes in chicken follicles throughout the ovulation cycle[J].PLoS One,2015,10(12):e0146028.
[32] BAO T T,YAO J W,ZHOU S,et al.Naringin prevents follicular atresia by inhibiting oxidative stress in the aging chicken[J].Poult Sci,2022,101(7):101891.
[33] JIANG J G,CHEN C L,CARD J W,et al.Cytochrome P450 2J2 promotes the neoplastic phenotype of carcinoma cells and is up-regulated in human tumors[J].Cancer Res,2005,65(11):4707-4715.
[34] GRESHAM R C H,BAHNEY C S,LEACH J K.Growth factor delivery using extracellular matrix-mimicking substrates for musculoskeletal tissue engineering and repair[J].Bioact Mater,2021,6(7):1945-1956.
[35] NY T,WAHLBERG P,BRNDSTRM I J M.Matrix remodeling in the ovary:regulation and functional role of the plasminogen activator and matrix metalloproteinase systems[J].Mol Cell Endocrinol,2002,187(1-2):29-38.
[36] HRABIA A,SOCHA J K,SECHMAN A.Involvement of matrix metalloproteinases (MMP-2,-7,-9) and their tissue inhibitors (TIMP-2,-3) in the regression of chicken postovulatory follicles[J].Gen Comp Endocrinol,2018,260:32-40.
[37] NIKOLOUDAKI G.Functions of matricellular proteins in dental tissues and their emerging roles in orofacial tissue development,maintenance,and disease[J].Int J Mol Sci,2021,22(12):6626.
[38] GIBLIN S P,MIDWOOD K S.Tenascin-C:form versus function[J].Cell Adh Migr,2015,9(1-2):48-82.
[39] CHIOVARO F,CHIQUET-EHRISMANN R,CHIQUET M.Transcriptional regulation of tenascin genes[J].Cell Adh Migr,2015,9(1-2):34-47.
[40] GOOSSENS K,VAN SOOM A,VAN ZEVEREN A,et al.Quantification of fibronectin 1 (FN1) splice variants,including two novel ones,and analysis of integrins as candidate FN1 receptors in bovine preimplantation embryos[J].BMC Dev Biol,2009,9:1.
[41] CUI Z F,LIU L B,KWAME AMEVOR F,et al.High expression of miR-204 in chicken atrophic ovaries promotes granulosa cell apoptosis and inhibits autophagy[J].Front Cell Dev Biol,2020,8:580072.
[42] LI X K.The FGF metabolic axis[J].Front Med,2019,13(5):511-530.
[43] PUSCHECK E E,PATEL Y,RAPPOLEE D A.Fibroblast growth factor receptor (FGFR)-4,but not FGFR-3 is expressed in the pregnant ovary[J].Mol Cell Endocrinol,1997,132(1-2):169-176.
[44] ZHAO W J,ZHU L L,YANG W Q,et al.LPAR5 promotes thyroid carcinoma cell proliferation and migration by activating class IA PI3K catalytic subunit p110β[J].Cancer Sci,2021,112(4):1624-1632.
[45] HERNNDEZ-BARRANCO A,SANTOS V,MAZARIEGOS M S,et al.NGFR regulates stromal cell activation in germinal centers[J].Cell Rep,2024,43(2):113705.
(編輯 郭云雁)
