腸炎沙門菌菌毛重組蛋白對其細胞黏附的競爭性阻斷效應
2024-12-18 00:00:00彭娜娜寧慧敏陳玉豪李欣穎祝福強于國濱董偉
畜牧獸醫學報 2024年11期

摘 要: 雞沙門菌通過其菌毛、鞭毛黏附宿主小腸上皮細胞是其致病過程不可或缺的重要環節。本研究旨在分析原核表達的菌毛重組蛋白FimA-sefA對腸炎沙門菌黏附小腸上皮細胞(IPEC-J2)競爭性阻斷效應。將FimA、sefA基因片段克隆至pET28a(+),構建重組質粒pET28a-FimA-sefA,轉化至菌株BL21(DE3)經鎳柱親和層析法(Ni-NTA)純化獲得重組蛋白FimA-sefA;利用間接免疫熒光法(IFA)、平板計數法分析腸炎沙門菌對IPEC-J2細胞的黏附。結果顯示:表達的重組蛋白FimA-sefA分子量為36 ku;重組蛋白FimA-sefA、重組菌及腸炎沙門菌均能黏附IPEC-J2細胞;重組蛋白FimA-sefA及重組菌均能競爭性阻斷腸炎沙門菌對IPEC-J2細胞的黏附,重組菌的阻斷效應強于重組蛋白。因此,我們認為重組蛋白FimA-sefA能夠在一定程度上競爭性阻斷腸炎沙門菌對IPEC-J2細胞的黏附,為預防及減輕腸炎沙門菌感染策略提供了新的思路。
關鍵詞: 腸炎沙門菌;菌毛蛋白;FimA-sefA;IPEC-J2;細胞黏附
中圖分類號:S852.612
文獻標志碼:A
文章編號:0366-6964(2024)11-5183-08
收稿日期:2023-12-22
基金項目:云南省重大科技專項計劃(202202AE090032);橫向課題(2022xczx-416;2021kjc-js178)
作者簡介:彭娜娜(1998-),女,湖南岳陽人,碩士生,主要從事動物疫病診斷與防控研究,E-mail:1625989715@qq.com
*通信作者:董 偉,主要從事獸藥新制劑研發及動物傳染病防控研究,E-mail: dongwei@hunau.edu.cn
Competitive Blocking Effect of Salmonella Enteritis Fimbriae Recombinant Protein
on Its Cell Adhesion
PENG" Nana, NING" Huimin, CHEN" Yuhao, LI" Xinying, ZHU" Fuqiang, YU" Guobin, DONG" Wei*
(College of Veterinary Medicine, Hunan Agricultural University, Changsha 410128, China)
Abstract:" Adhesion of host small intestinal epithelial cells through fungal hairs and flagella is an indispensable and important link in pathogenic processes of Salmonella Enterison. This study aims to analyze the competitive blocking effect of the hair recombinant protein FimA-sefA expressed by prokaryotes on the adhesion of Salmonella Enteritis to intestinal epithelial cells (IPEC-J2). FimA and sefA gene fragments were cloned into pET28a (+), resulting in the construction of a recombinant plasmid pET28a-FimA-sefA, which was then transformed into the strain BL21 (DE3). The recombinant protein FimA-sefA was purified using nickel-column affinity chromatography (Ni-NTA). The adhesion of enteritis to IPEC-J2 cells was analyzed using indirect immunofluorescence (IFA) and plate counting. The molecular weight of the expressed recombinant protein FimA-sefA was 36 ku; the recombinant protein FimA-sefA, recombinant bacteria, and Salmonella Enteritis can adhere to IPEC-J2 cells. Recombinant protein FimA-sefA and recombinant bacteria can competitively block the adhesion of Salmonella Enteritis to IPEC-J2 cells, and the blocking effect of recombinant bacteria was stronger than that of the recombinant protein. We believe that the recombinant protein FimA-sefA can competitively block the adhesion of Salmonella Enteritis to IPEC-J2 cells to a certain extent, providing a new strategy for preventing and reducing enteritis infection.
Key words: Salmonella Enteritidis; fimbria protein; FimA-sefA; IPEC-J2; cell adhesion enteritidis
*Corresponding author:" DONG Wei, E-mail: dongwei@hunau.edu.cn
沙門菌目前已有超過3 000多種血清型,在家禽產業鏈中主要以腸炎沙門菌(Salmonella Enteritidis)和雞白痢沙門菌為主,其中腸炎沙門菌作為優勢血清型常引起雞群腹污、發熱、甚至死亡,給養殖戶造成巨大的經濟損失[1]。大多數沙門菌宿主特異性較弱,可污染整個家禽生產鏈,通過肉、蛋等轉移給人類并引起腹污、胃腸炎及敗血癥等疾病[2-3]。沙門菌分布廣泛,血清型繁多,宿主多樣,傳播途徑眾多,耐藥性普遍,凈化困難等因素,導致雞沙門菌病防控比較困難[4]。應用抗生素控制雞群中沙門菌感染會增加抗生素殘留或多重抗生素耐藥的風險[5]。因此,迫切需要新的生物防治手段來控制雞群沙門菌感染。
沙門菌進入動物消化道后,憑借腸道蠕動功能進入小腸并通過其菌毛、鞭毛等黏附至宿主小腸上皮細胞。菌毛作為腸炎沙門菌的重要毒力因子,在黏附、定殖機體上皮細胞黏膜上發揮了重要作用[6-10]。腸炎沙門菌SEF21(Ⅰ型)菌毛廣泛分布于沙門菌屬,而SEF14菌毛特異性表達于腸炎沙門菌和都伯林沙門菌[11]。Ⅰ型菌毛介導細菌與黏膜上皮細胞表面的D-甘露糖受體結合,促使細菌定殖黏膜,激發巨噬細胞、肥大細胞的免疫反應[12]。在SEF14菌毛的作用下,腸炎沙門菌能夠更好地黏附于腸上皮細胞[13-14]。SEF14菌毛是可以用于區分菌株血清型的毒力因子,有助于腸炎沙門菌黏附至鼠上皮細胞,而不能與人 HEp-2、Caco-2、INT-407或 HeLa 細胞黏附[15]。免疫SEF14菌毛蛋白可以產生強烈的保護性免疫應答,介導細菌與宿主吞噬細胞之間的相互作用[16]。
FimA是作為Ⅰ型菌毛亞基的主要蛋白,在Ⅰ型菌毛生物合成中起關鍵作用[17-18]。sefA編碼SEF14菌毛的主要亞單位蛋白,是特異性檢測腸炎沙門菌的一種靶蛋白[19]。該蛋白能增強腸炎沙門菌對腸上皮細胞的黏附以及其在巨噬細胞中的存活能力,并能強化小鼠腸道中腸炎沙門菌的毒力,這些都證明SEF14菌毛在腸炎沙門菌感染的過程中發揮了重要毒力作用[20-21]。
本研究根據腸炎沙門菌菌毛在其致病過程中的作用機制,表達編碼SEF21、SEF14菌毛的主要亞單位蛋白FimA、sefA,并研究菌毛蛋白能否競爭性阻斷腸炎沙門菌黏附腸上皮細胞,從而阻止腸炎沙門菌的感染致病過程。本研究旨在探究通過阻斷沙門菌侵染腸上皮細胞來防控雞沙門菌病的潛能,為雞沙門菌病的凈化提供新策略。
1 材料與方法
1.1 細菌菌株、質粒及細胞
腸炎沙門菌菌株CICC 21513購自中國工業微生物菌種保藏中心;pET28a(+)載體、豬小腸上皮細胞(IPEC-J2)由本實驗室保存;E. coli DH5a、E. coli BL21(DE3)感受態購自北京擎科生物科技有限公司。
1.2 主要試劑
限制性內切酶(Sal Ⅰ/Not Ⅰ)、T4 DNA Ligase購自美國紐英倫生物技術(NEB)公司;HRP標記羊抗兔IgG抗體、HRP標記羊抗鼠IgG抗體、FITC標記的羊抗兔IgG購自美國Abbkine公司;質粒小提試劑盒、瓊脂糖凝膠DNA回收試劑盒、BCA蛋白濃度測定試劑盒購自南京諾唯贊生物科技股份有限公司;Ni-NTA親和純化試劑盒購自德國QIAGEN公司;腸炎沙門菌LPS單克隆抗體購自長沙博儀生物科技有限公司;DMEM/F12、胰酶、PBS購自Gibco;胎牛血清購自南京森貝伽生物科技有限公司。
1.3 重組質粒的構建
根據GenBank中公布的FimA和sefA基因序列(S76043.1、L03833.1),用柔性連接肽(GGGGS)連接,并添加6×His標簽,全長共1 095 bp,提交至上海擎科生物科技有限公司合成。合成引物F:5′-GCGTCGACATGAAACATAAATTAATG-3′,R:5′-TTTAGCGGCCGCTTAGTGGTGGTGGT-3′擴增、膠回收目的片段,將產物與pET28a(+)空載體雙酶切并做產物的回收,連接、轉化及菌落的篩選獲得重組表達載體,并將測序正確的重組質粒命名為pET28a-FimA-sefA。
1.4 FimA-sefA重組蛋白的表達、鑒定與純化
將重組質粒pET28a-FimA-sefA轉化至大腸桿菌BL21(DE3)感受態細胞,37 ℃培養至OD600 nm為0.5左右。加入IPTG(終濃度為1 mmol·L-1)30℃誘導蛋白12 h,收獲細菌、超聲裂解,SDS-PAGE電泳分析及Western blot鑒定分析,鑒定重組蛋白的可溶性及與鼠His-tag單克隆抗體的特異性反應。Ni-NTA親和層析柱純化目的蛋白,并使用BCA蛋白濃度測定試劑盒測定蛋白的濃度。
1.5 間接免疫熒光法檢測FimA-sefA蛋白、重組菌及腸炎沙門菌黏附IPEC-J2細胞
FimA-sefA蛋白黏附細胞試驗:IPEC-J2細胞鋪板,待細胞長至90%左右,固定、封閉,加入40 μg·mL-1的FimA-sefA蛋白37 ℃作用60 min,PBS漂洗,加入兔抗FimA-sefA蛋白抗體(1∶1 000稀釋,由本實驗室制備),4 ℃孵育過夜,漂洗后加入FITC-羊抗兔IgG(1∶500稀釋),37 ℃孵育60 min,最后DAPI染色,置于熒光顯微鏡下觀察。
腸炎沙門菌黏附細胞試驗:將腸炎沙門菌接種至CFA培養基中,37 ℃靜置48 h,使其菌毛充分表達,并進行濃度測定。以MOI=100感染IPEC-J2細胞,孵育60 min,加入兔抗腸炎沙門菌抗體(1∶1 000稀釋,由實驗室制備),其余步驟與FimA-sefA蛋白黏附細胞試驗一致。
重組菌黏附細胞試驗:含有陽性質粒pET28a-FimA-sefA的E. coli BL21(DE3)菌株為重組菌,濃度測定后,以MOI=100感染IPEC-J2細胞,孵育60 min,設置對照組為同等培養條件的空載體菌株,一抗使用兔抗FimA-sefA蛋白抗體(1∶1 000稀釋),其余步驟與FimA-sefA蛋白黏附細胞試驗一致。
1.6 FimA-sefA蛋白、重組菌阻斷腸炎沙門菌黏附IPEC-J2細胞試驗
MTT試驗檢測FimA-sefA蛋白對細胞活力的影響。
平板計數法:IPEC-J2細胞鋪板,待細胞長至90%左右,加入蛋白(8、10、12、14、16、18 μg·mL-1)和重組菌(MOI=100、MOI=200、MOI=400)分別與細胞37 ℃孵育60 min,PBS漂洗后,加入腸炎沙門菌(MOI=100),與細胞37 ℃孵育60 min,陽性對照組僅加入腸炎沙門菌,漂洗后,加入100 μL胰酶,待細胞脫落后,加入細胞培養基終止反應,收集樣品,倍比稀釋后涂布至固體LB平板,37 ℃過夜培養,進行菌落計數,每個處理重復平行3次,試驗重復3次后,對數據進行統計并處理。
間接免疫熒光法:IPEC-J2細胞鋪板,待細胞長至90%左右,加入蛋白和重組菌分別與細胞37℃孵育60 min,PBS漂洗后,加入腸炎沙門菌(MOI=100),與細胞37℃孵育60 min,陽性對照組僅加入腸炎沙門菌,漂洗后,固定、封閉后,加入鼠抗腸炎沙門菌LPS單克隆抗體(1:50稀釋),4 ℃孵育過夜,漂洗后加入FITC-羊抗鼠IgG(1∶500稀釋),37 ℃孵育60 min,最后進行DAPI染色,置于熒光顯微鏡下觀察。
2 結 果
2.1 重組質粒的構建與蛋白表達
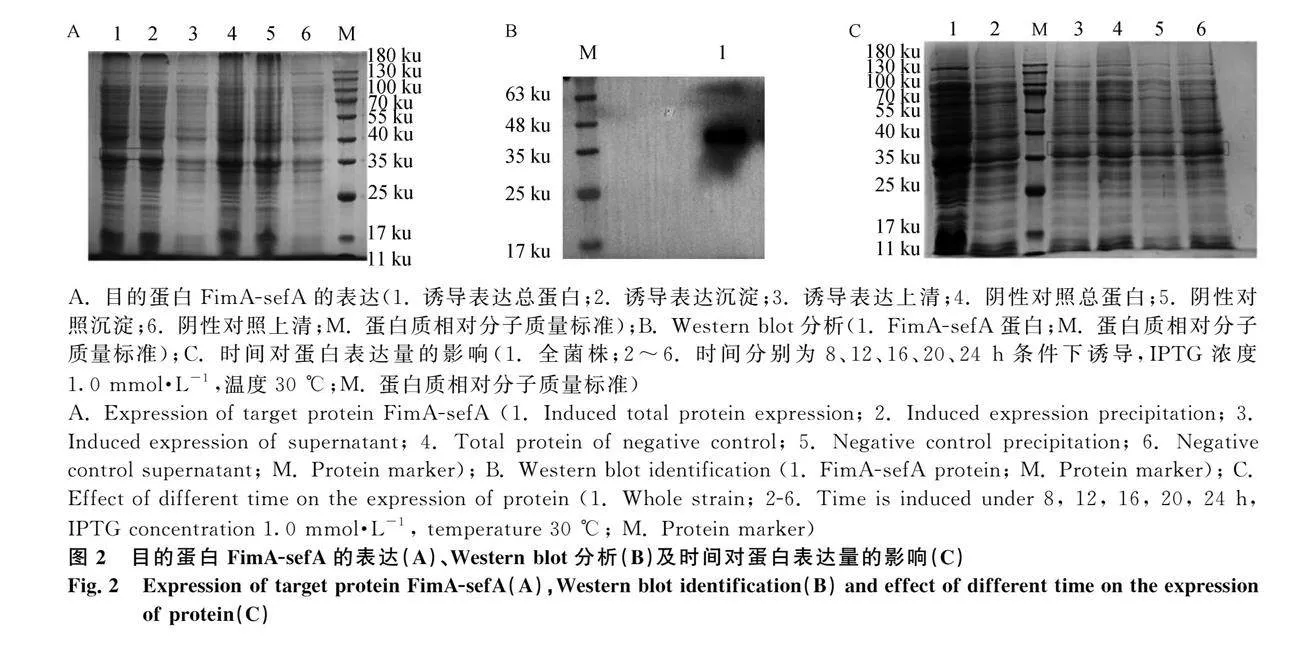
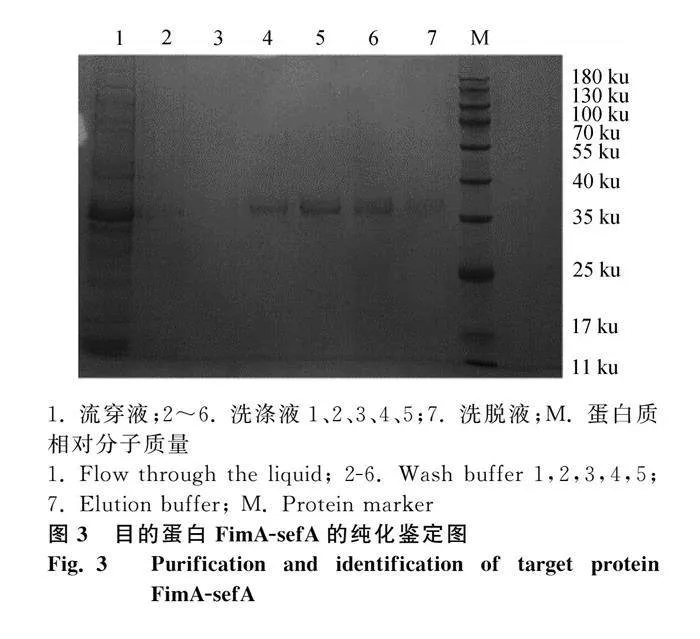
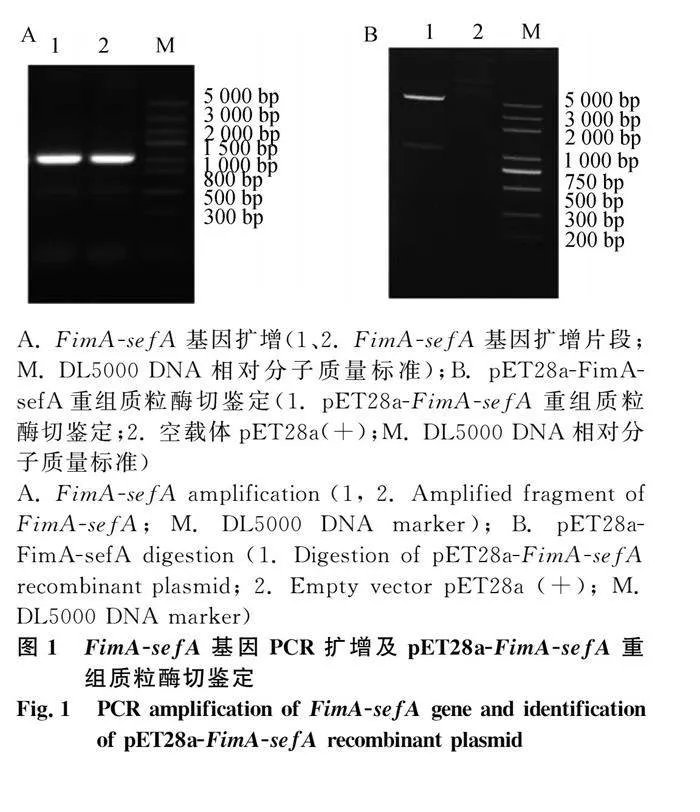
本研究成功表達FimA-sefA蛋白:克隆FimA-sefA基因(圖1A);雙酶切驗證重組質粒pET28a-FimA-sefA(圖1B);獲得重組質粒BL21(DE3)表達菌,IPTG誘導,30℃,12 h表達量最高;重組蛋白主要以包涵體表達,大小為36 ku(圖2A、C);鼠抗His-tag單克隆抗體能夠識別重組蛋白(圖2B);通過Ni-NTA親和層析柱純化得到目的蛋白(圖3)。經檢測蛋白濃度為267 μg·mL-1,-80 ℃保存備用。
2.2 FimA-sefA蛋白、重組菌及腸炎沙門菌對IPEC-J2細胞的黏附
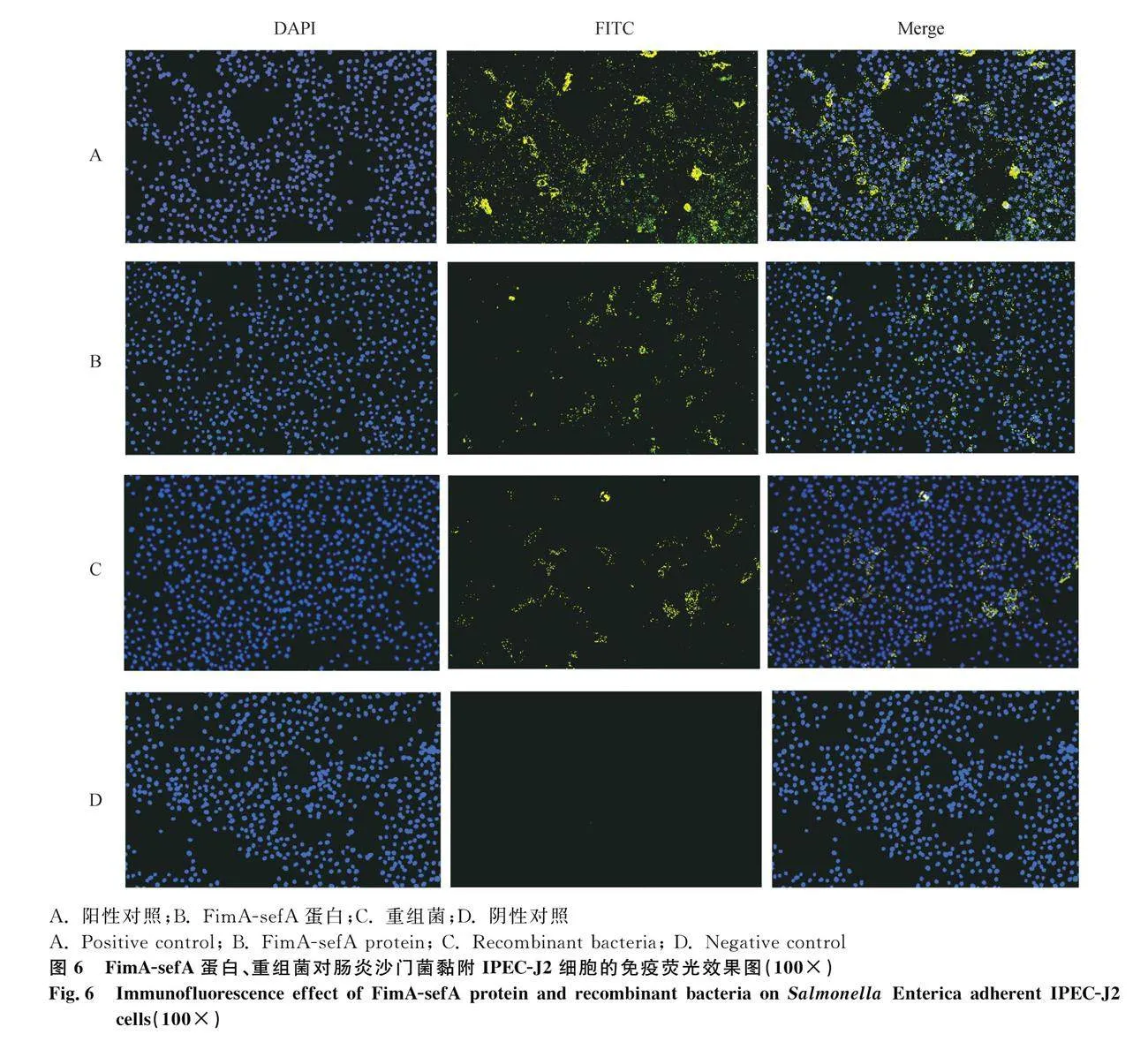
間接免疫熒光顯示FimA-sefA蛋白、重組菌、腸炎沙門菌均能黏附于IPEC-J2細胞(圖4)。
2.3 FimA-sefA蛋白、重組菌對腸炎沙門菌黏附阻斷
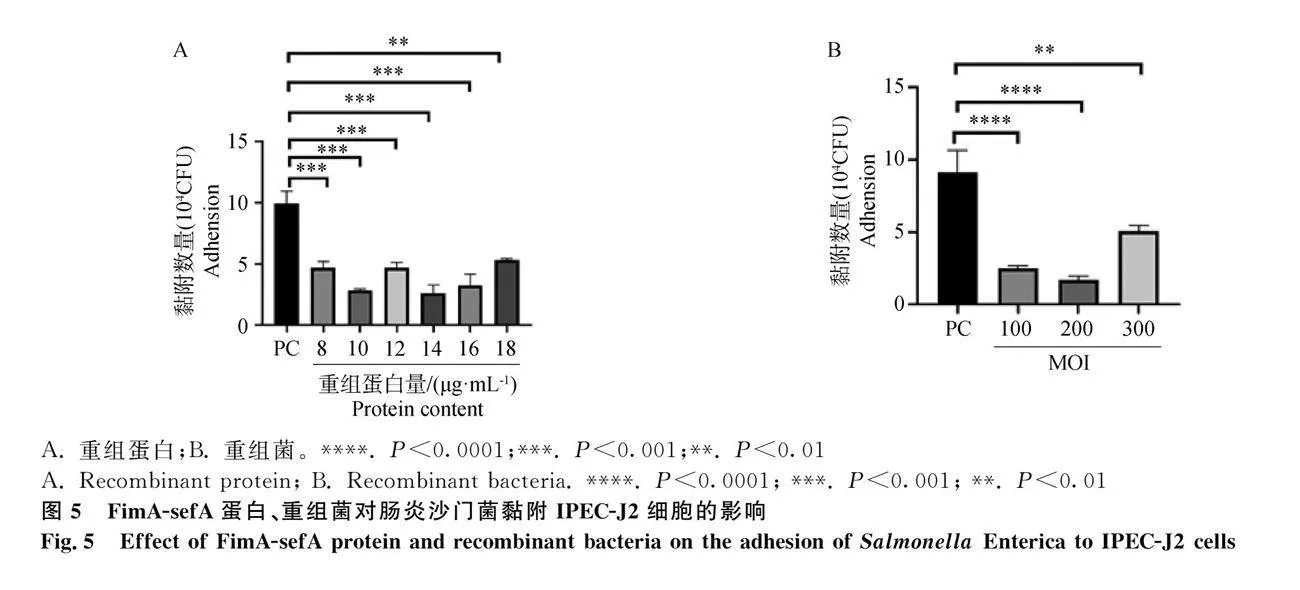
MTT證明低于18 μg·mL-1的FimA-sefA蛋白對細胞活力無影響;FimA-sefA蛋白與IPEC-J2細胞孵育后,與陽性對照組相比,腸炎沙門菌黏附數量會顯著降低(Plt;0.01),當蛋白為10、14 μg·mL-1時,阻斷效果顯著(Plt;0.001),但并非呈現依賴性效果(圖5A);重組菌與IPEC-J2細胞孵育后,同陽性對照組相比,腸炎沙門菌黏附數量會顯著降低(Plt;0.01,圖5B),當MOI=200時,阻斷效果最佳(Plt;0.000 1)。間接免疫熒光中,與陽性對照組相比,FimA-sefA蛋白組及重組菌組熒光強度較弱,陰性對照組成立,再次證明了重組蛋白能夠阻斷腸炎沙門菌對IPEC-J2細胞的黏附(圖6)。
3 討 論
腸炎沙門菌作為一種人畜共患病病原菌,對動物及人類健康造成威脅[22],因此防治腸炎沙門菌感染,建立公共衛生意識具有重要意義[23-24]。當前主要依賴抗菌藥物進行防治,但考慮到菌株的突變及抗菌藥物的耐藥性等問題[25-27],現行的沙門菌防控策略已經難以滿足生產實踐需求,亟需應用新策略預防腸炎沙門菌感染。腸炎沙門菌通過菌毛黏附小腸上皮細胞與M細胞,激活信號通路并進入吞噬泡內,擊毀細胞。本試驗室提出的“黏膜阻斷”防控沙門菌病新策略是應用沙門菌菌毛基因工程亞單位蛋白競爭性阻斷腸炎沙門菌黏附雞腸道黏膜上的小腸上皮細胞與M細胞,從而切斷腸炎沙門菌的感染過程。目前,沙門菌菌毛基因工程亞單位蛋白被廣泛應用于免疫和病原的檢測方面[28],能與腸上皮細胞及M細胞上的菌毛受體高親和度結合。
將FimA-sefA片段克隆至pET28a(+),并轉化至菌株BL21(DE3),經Ni-NTA親和層析柱純化獲得重組蛋白FimA-sefA;IFA檢測到重組蛋白、重組菌、腸炎沙門菌能與IPEC-J2細胞黏附;平板計數法與IFA證明了重組蛋白FimA-sefA、重組菌均能競爭性阻斷腸炎沙門菌對IPEC-J2細胞的黏附,重組菌阻斷效應強于重組蛋白。因此,本研究不僅再一次證明了SEF21、SEF14菌毛在腸炎沙門菌黏附細胞過程中的重要性,同時證明了應用重組FimA-sefA蛋白、重組菌阻斷腸炎沙門菌黏附IPEC-J2細胞,從而防控雞腸炎沙門菌病的可行性。
重組蛋白對IPEC-J2細胞吸附熒光強于重組菌對IPEC-J2細胞,而重組菌阻斷腸炎沙門菌對IPEC-J2細胞的黏附效應強于重組蛋白阻斷腸炎沙門菌對IPEC-J2細胞。我們推測重組菌釋放FimA-sefA蛋白濃度高,且重組菌菌體表面存在的FimA-sefA蛋白與IPEC-J2細胞上FimA-sefA蛋白受體結合后,致使重組菌黏附在IPEC-J2細胞表面,對后續的腸炎沙門菌的黏附形成了空間位阻,減少了貼近IPEC-J2細胞表面的腸炎沙門菌。
重組蛋白對IPEC-J2細胞吸附的熒光結果顯示其密集吸附于每一個細胞表面,但重組蛋白阻斷腸炎沙門菌黏附IPEC-J2細胞的熒光結果顯示仍有腸炎沙門菌黏附IPEC-J2細胞。這是由于IPEC-J2細胞表面FimA-sefA蛋白受體的表達是一個動態持續的過程,隨著細胞周圍游離的FimA-sefA蛋白逐漸減少,越來越多的細胞受體裸露出來供腸炎沙門菌菌體表面的FimA-sefA蛋白結合,從而使腸炎沙門菌成功黏附。這些結果提示應用單純的FimA-sefA重組蛋白阻斷腸炎沙門菌黏附,其阻斷效果并不能滿足生產實踐需求,需要構建表達FimA-sefA蛋白的乳酸菌,使得乳酸菌能在腸道黏膜定植,并能連續不斷分泌FimA-sefA蛋白,持續封閉腸上皮細胞不斷表達的FimA-sefA蛋白受體。這樣機體能夠通過口服表達FimA-sefA蛋白的乳酸菌防治腸炎沙門菌感染。
相較于疫苗,阻斷劑在腸炎沙門菌病防控實踐中更具有獨特的優勢。免疫接種的作用機制是反向阻斷,疫苗刺激機體產生的抗體或高免血清抗體占位封閉的是菌體抗原。由于腸道沙門菌存在不同血清型菌株,要保障占位封閉效果,刺激機體產生抗體的疫苗菌株必須要與流行菌株一致。腸炎沙門菌菌株雖然存在多種突變的情況,但腸炎沙門菌所有菌株的靶細胞受體是相同的,且不會發生改變。因此,阻斷劑能有效阻斷不同腸炎沙門菌菌株黏附靶細胞。
4 結 論
本研究利用原核表達系統成功表達腸炎沙門菌菌毛重組蛋白——FimA-sefA蛋白,同時利用間接免疫熒光法、平板計數法證明了重組蛋白、重組菌均能有效阻斷腸炎沙門菌黏附IPEC-J2細胞,為預防和減輕腸炎沙門菌感染提供了新策略。
參考文獻(References):
[1] KNODLER L A, ELFENBEIN J R. Salmonella enterica[J]. Trends Microbiol, 2019, 27(11):964-965.
[2] 鄭 琳, 魏炳棟, 滑 峰, 等. 裂解性噬菌體對肉仔雞感染腸炎沙門菌的治療效果[J]. 畜牧獸醫學報, 2024, 55(3):1314-1327.
ZHENG L, WEI B D, HUA F, et al. Therapeutic effect of lytic phage on Salmonella enteritidis infection in broilers[J]. Acta Veterinaria et Zootechnica Sinica, 2024, 55(3):1314-1327. (in Chinese)
[3] STILZ C R, CAVALLO S, GARMAN K, et al. Salmonella enteritidis outbreaks associated with egg-producing farms not regulated by food and drug administration’s egg safety rule[J]. Foodborne Pathog Dis, 2022, 19(8):529-534.
[4] BEN SALEM R, ABBASSI M S, GARCA V, et al. Antimicrobial drug resistance and genetic properties of Salmonella enterica serotype Enteritidis circulating in chicken farms in Tunisia[J]. J Infect Public Health, 2017, 10(6):855-860.
[5] OBE T, NANNAPANENI R, SCHILLING W, et al. Prevalence of Salmonella enterica on poultry processing equipment after completion of sanitization procedures[J]. Poult Sci, 2020, 99(9):4539-4548.
[6] 張旭梅, 魏玉榮, 許丞惠, 等. 基于網絡藥理學和試驗驗證分析小檗堿治療雞沙門菌感染的作用機制[J]. 畜牧獸醫學報, 2023, 54(8):3557-3570.
ZHANG X M, WEI Y R, XU C H, et al. To analyze the mechanism of berberine in the treatment of Salmonella gallinarum infection based on network pharmacology and experimental verification[J]. Acta Veterinaria et Zootechnica Sinica, 2023, 54(8):3557-3570. (in Chinese)
[7] TOWNSEND S M, KRAMER N E, EDWARDS R, et al. Salmonella enterica serovar Typhi possesses a unique repertoire of fimbrial gene sequences[J]. Infect Immun, 2001, 69(5):2894-2901.
[8] BODDICKER J D, LEDEBOER N A, JAGNOW J, et al. Differential binding to and biofilm formation on, HEp-2 cells by Salmonella enterica serovar Typhimurium is dependent upon allelic variation in the fimH gene of the fim gene cluster[J]. Mol Microbiol, 2002, 45(5):1255-1265.
[9] HUMPHRIES A D, RAFFATELLU M, WINTER S, et al. The use of flow cytometry to detect expression of subunits encoded by 11 Salmonella enterica serotype Typhimurium fimbrial operons[J]. Mol Microbiol, 2003, 48(5):1357-1376.
[10] QUAN G M, XIA P P, ZHAO J, et al. Fimbriae and related receptors for Salmonella Enteritidis[J]. Microb Pathog, 2019, 126:357-362.
[11] ZHU C H, WU J, CHEN W W, et al. Difference and variation of the sef14 operon gene clusters in S." pullorum[J]. J Basic Microbiol, 2010, 50 Suppl 1:S120-S123.
[12] LI W Z, WATARAI S, KODAMA H. Identification of glycosphingolipid binding sites for SEF21-fimbriated Salmonella enterica serovar Enteritidis in chicken oviductal mucosa[J]. Vet Microbiol, 2003, 93(1):73-78.
[13] CROWE S J, GREEN A, HERNANDEZ K, et al. Utility of combining whole genome sequencing with traditional investigational methods to solve foodborne outbreaks of salmonella infections associated with chicken:a new tool for tackling this challenging food vehicle[J]. J Food Prot, 2017, 80(4):654-660.
[14] ARISAWA T, NAKAMURA M, OTSUKA T, et al. Genetic polymorphisms of MAFK, encoding a small Maf protein, are associated with susceptibility to ulcerative colitis in Japan[J]. World J Gastroenterol, 2017, 23(29):5364-5370.
[15] CAI L L, XIE Y T, SHAO L T, et al. SaaS sRNA promotes Salmonella intestinal invasion via modulating MAPK inflammatory pathway[J]. Gut Microbes, 2023, 15(1):2211184.
[16] CAI L L, XIE Y T, HU H J, et al. A Small RNA, SaaS, promotes Salmonella pathogenicity by regulating invasion, intracellular growth, and virulence factors[J]. Microbiol Spectr, 2023, 11(1):e0293822.
[17] MENG X, MENG X C, WANG J Q, et al. Small non-coding RNA STnc640 regulates expression of fimA fimbrial gene and virulence of Salmonella enterica serovar Enteritidis[J]. BMC Vet Res, 2019, 15(1):319.
[18] 張成棟. Ⅰ型菌毛介導鼠傷寒沙門氏菌對小鼠的毒力及對抗原遞呈細胞侵襲力的研究[D]. 武漢:華中農業大學, 2009.
ZHANG C D. Type 1 fimbriae of Salmonella typhimurium mediate bacterial virulence and invasion to antigen presenting cells in mice[D]. Wuhan:Huazhong Agricultural University, 2009. (in Chinese)
[19] GONG J S, ZHUANG L L, ZHU C H, et al. Loop-mediated isothermal amplification of the sefA gene for rapid detection of Salmonella enteritidis and salmonella Gallinarum in chickens[J]. Foodborne Pathog Dis, 2016, 13(4):177-181.
[20] JONES B D, GHORI N, FALKOW S. Salmonella typhimurium initiates murine infection by penetrating and destroying the specialized epithelial M cells of the Peyer’s patches[J]. J Exp Med, 1994, 180(1):15-23.
[21] THORNS C J, TURCOTTE C, GEMMELL C G, et al. Studies into the role of the SEF14 fimbrial antigen in the pathogenesis of Salmonella enteritidis[J]. Microb Pathog, 1996, 20(4):235-246.
[22] BESSER J M. Salmonella epidemiology:a whirlwind of change[J]. Food Microbiol, 2018, 71:55-59.
[23] QIU Q H, DEWEY-MATTIA D, SUBRAMHANYA S, et al. Food recalls associated with foodborne disease outbreaks, United States, 2006-2016[J]. Epidemiol Infect, 2021, 149:e190.
[24] FADLALLAH S M, SHEHAB M, CHEAITO K, et al. PulseNet lebanon:an overview of its activities, outbreak investigations, and challenges[J]. Foodborne Pathog Dis, 2019, 16(7):498-503.
[25] EL-SAADONY M T, SALEM H M, EL-TAHAN A M, et al. The control of poultry salmonellosis using organic agents:an updated overview[J]. Poult Sci, 2022, 101(4):101716.
[26] XU D S, JI L, YAN W, et al. Characterization of clinical Salmonella entericas Trains in Huzhou, China[J]. Can J Infect Dis Med Microbiol, 2022, 2022:7280376.
[27] ZHOU X J, XU L, XU X B, et al. Antimicrobial resistance and molecular characterization of Salmonella enterica serovar enteritidis from retail chicken products in Shanghai, China[J]. Foodborne Pathog Dis, 2018, 15(6):346-352.
[28] 侯千禧, 顧宣強, 劉家奇, 等. 腸炎沙門氏菌SEF14菌毛的表達及其對該菌的特異性檢測研究[J]. 中國預防獸醫學報, 2022, 44(1):91-95.
HOU Q X, GU X Q, LIU J Q, et al. Expression of SEF14 fimbriae protein and research on specific diagnostic technology of Salmonella enteritidis[J]. Chinese Journal of Preventive Veterinary Medicine, 2022, 44(1):91-95. (in Chinese)
(編輯 白永平)
