間作模式對油橄欖土壤養分及土壤微生物的影響
2024-12-27 00:00:00黃楚亦胡青趙敏陸斌張光華王猛
經濟林研究 2024年4期
關鍵詞:群落結構


關鍵詞:油橄欖;間作;土壤養分;微生物多樣性;群落結構
油橄欖Oleaeuropaea,為木樨科Oleaceae木樨欖屬Olea常綠小喬木,是世界四大木本油料植物之一。油橄欖原產于地中海地區,在我國的主要適生區包括金沙江干熱河谷區、白龍江低山河谷區、長江三峽低山河谷區[1]。這些區域光熱資源充沛,為油橄欖的生長提供了良好的自然條件。然而,這些區域多位于水土流失和養分流失嚴重的半山干旱區[2],土壤質量較差,地表植被稀疏,生態環境脆弱。此外,油橄欖幼林期長,種植4~5a后才開始開花結果,7a后才能達到盛產期,這在一定程度上限制了油橄欖產業的生產效率和生態效益。因此,改善油橄欖園土壤管理方式是當前油橄欖產業發展亟待解決的問題之一。間作是一種高效的經濟林經營措施,是目前在國內外廣泛應用的一種現代化可持續發展的土壤管理方式[3]。將不同生態位作物復合種植,不僅能充分利用林地空間資源,提高土地資源的利用效率,還能發揮間作作物的生態功能,改善土壤環境,對提高油橄欖園的土地利用率及其生態和經濟效益具有深遠的意義。
土壤微生物是土壤生態系統的關鍵組成部分,是土壤生態系統中較為活躍的組分之一,也是養分轉化和物質循環等土壤生態過程中必不可少的生物因素[4],對維持土壤生態系統的穩定和健康起到重要作用。土壤養分是植物正常生長發育的基礎[5],土壤有機質和養分主要來源于土壤微生物對凋落物以及動植物殘體的分解,與土壤微生物多樣性和豐度密切相關。間作可以通過改變植物生理特征及根系分泌特性間接影響土壤微生物多樣性和群落結構,從而改善土壤微環境[6-7]。趙春梅等[8]發現,在橡膠林下間作王草、益智和咖啡作物有利于提高土壤微生物群落功能多樣性,改善土壤生態環境。不同的間作作物的生理特征和生物學特性不同,其對土壤養分及土壤微生物的影響也是不同的,如楊宏偉等[9]發現在油橄欖林間作百喜草增加了土壤有機質含量,并能夠有效改善根際土壤微環境,而嚴毅等[10]發現在油橄欖林下種植百合后林地土壤養分含量明顯降低。因此,研究不同間作模式下油橄欖土壤養分變化及微生物動態,對選擇適宜的油橄欖間作模式和促進我國油橄欖產業高效發展具有重要意義。
目前關于油橄欖間作經營方式的探索已取得一些成果,如油橄欖間作毛苕子、百喜草、紅三葉等綠肥作物[11-13],以及在油橄欖林下種植大球蓋菇、榆黃蘑、靈芝和小麥等作物[14-15]。這些研究內容集中于根系發育形態、光合特性、抗旱生理、經濟產量等方面。然而,關于不同間作模式下油橄欖林地土壤養分變化、微生物多樣性及其群落結構變化等的深入分析報道較為鮮見。本研究中以油橄欖單作為對照,以3種油橄欖間作模式為研究對象,分析了間作模式對油橄欖土壤養分含量、土壤微生物多樣性和群落結構的影響,旨在為探索油橄欖高效栽培模式和改善油橄欖林地土壤生態環境提供參考。
1材料與方法
1.1試驗區概況
研究地位于云南省麗江市永勝縣六德鄉玉水村油橄欖栽培示范園(100°86′E,26°57′N)。該園地處云南省北部金沙江中游亞熱帶干熱河谷地帶,平均海拔1991.7m,年平均氣溫16.2℃,年平均日照2403h,年降水量800.40mm。干、熱是該地區典型的氣候特征,降水主要集中在夏秋季節,冬春干旱,全年光照充足。示范園土壤以砂壤土為主,土層較厚。整個園區油橄欖種植面積超46.67hm2,在試驗基地內選擇坡向、海拔且水肥管理措施基本一致的油橄欖‘佛奧’品種種植區域進行試驗,株行距為5m×7m。
1.2試驗設計
油橄欖為2017年初種植的品種‘佛奧’。試驗設4個處理,以油橄欖單作為對照(CK),其余為間作模式,分別為油橄欖林下間作滇黃精(FD)、紫花苜蓿草(FZ)、3種混播草(F3)。2018年,將紫花苜蓿草與3種混播草種(紫花苜蓿草、黑麥草、金雞菊)撒播于油橄欖林地內,將滇黃精種植于油橄欖行間(株行距0.5m×1.0m)。試驗期間油橄欖樹的施肥、灌溉等管護措施保持一致,均按照常規方法統一進行管理。
1.3試驗方法
1.3.1土壤樣品采集
2023年5月,采用典型的“S”形布設土壤取樣法,在每個處理區域內設置3個土壤采樣點。在距離油橄欖樹干基部50cm處,除去地面的枯枝落葉、碎石和雜物,垂直采集深度0~10cm的土壤。每個樣地采取3份土樣,每處理重復3次,將其混合均勻為1份土樣。將每份土壤樣本取0.5kg,立即裝入無菌自封袋并編號,置于冷凍箱中保存備用,用于土壤微生物群落結構及多樣性的測定。將一部分土壤樣品自然風干,用于養分含量等指標的測定。
1.3.2土壤理化指標測定[16]
土壤pH值采用電位計法(土、水體積比1∶2.5)測定,有機碳含量采用重鉻酸鉀容量法測定,全氮含量采用凱氏定氮法測定,速效鉀和全鉀含量采用火焰原子分光光度法測定,有效磷和全磷含量采用鉬銻抗比色法測定,堿解氮含量采用堿解擴散法測定。
1.3.3土壤DNA的提取及微生物高通量測序
利用IlluminaMiseq高通量測序技術對細菌和真菌多樣性進行分析。采用OMEGASoilDNAKit試劑盒提取土壤基因組DNA,利用1%瓊脂糖凝膠電泳檢測DNA質量,利用NanodropNC2000測定DNA濃度,具體操作步驟以自帶說明書為標準。以提取的基因組DNA為模板,以細菌通用引物338F(5′-ACTCCTACGGGAGGCAGCA-3′)和806R(5′-GGACTACHVGGGTWTCTAAT-3′)對16S的V3~V4區進行PCR擴增,對土壤真菌ITS的V1區進行擴增,擴增引物為ITS5F(5′-GGAAGTAAAAGTCGTAACAAGG-3′)和ITS1R(5′-GCTGCGTTCTTCATCGATGC-3′)。利用IlluminaMiseq技術平臺進行文庫構建和Paired-end測序。
1.4數據處理與分析
使用Vsearch和Cutadapt軟件進行去引物、拼接、質量過濾、去重、去嵌合體、聚類等流程,獲取高質量序列,在97%相似度水平對高質量序列聚類,生成ASVs特征序列,并分別輸出代表序列和OTU表格,最后去除OTU表格中豐度為1的OTU及其代表序列。使用Silva數據庫和UNITE數據庫比對ASVs特征序列,得到每個ASV對應的分類學信息。使用R腳本的VennDiagram包制作韋恩圖,利用QIIME2分析平臺對物種分類組成情況和物種多樣性進行分析。采用SPSS22.0軟件分析數據差異的顯著性,采用Excel軟件制作表格。
2結果與分析
2.1不同間作模式下土壤養分含量和pH值
由表1可知,4個處理下土壤的pH值為8.12~8.36,均為堿性土壤,FD、F3間作處理下土壤pH值相對較低,但處理間差異均不顯著。相較于CK,FD間作模式下土壤有機碳、全氮、全磷、全鉀的含量均有所降低,但并無顯著差異,速效鉀、有效磷、堿解氮含量分別顯著降低了55.45%、45.55%和33.46%。相較于CK,FZ間作模式下土壤全氮、全磷含量無顯著差異,間作后有機碳、速效鉀和堿解氮含量分別顯著降低了60.73%、56.01%和77.48%,全鉀、有效磷含量顯著增加了29.40%、49.87%,且含量均高于其他3個處理。相較于CK,F3間作模式下土壤全鉀含量顯著增加了24.59%,堿解氮含量有所降低但不顯著,其他養分含量相較于CK均無顯著差異,但含量均有所增加。綜上可見,與對照組CK相比,3種油橄欖間作模式均能在一定程度上改變油橄欖林地土壤養分含量,FD和FZ間作處理下土壤部分速效養分含量顯著降低,F3間作處理對提高油橄欖林地土壤養分含量的效果最佳。
2.2不同間作模式下土壤微生物群落狀況
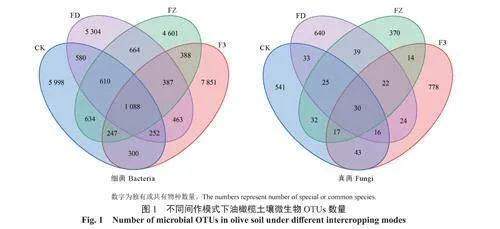
2.2.1微生物OTUs數量
由不同間作模式下油橄欖土壤微生物OTUs數量韋恩圖可知(圖1),4種間作模式下土壤細菌OTUs總數量為38652,真菌OTUs總數量為3059,共有細菌OTUs數量為1088,占OTUs總數量的2.81%,共有真菌OTUs數量為30,占OTUs總數量的0.98%。CK、FD、FZ、F3間作模式下土壤特有的細菌OTUs數量分別為5998、5304、4601、7851,分別占細菌特有OTUs總數量的25.25%、22.33%、19.37%、33.05%。CK、FD、FZ、F3間作模式下土壤特有的真菌OTUs數量分別為541、640、370、778,分別占真菌特有OTUs總數量的23.23%、27.48%、15.89%、33.40%。4種間作模式下土壤細菌和真菌所包含的OTUs數量和特有的OTUs數量均以F3間作模式最多,FZ間作模式最少。根據細菌的OTUs總數量和特有OTUs數量由大到小排列,各處理依次為F3、CK、FD、FZ;根據真菌的OTUs數量由大到小排列,各處理依次為F3、FD、CK、FZ。綜上,與油橄欖單作CK相比較,不同處理間細菌和真菌OTUs數量均有一定的差別。F3處理模式能在一定程度上增加土壤中細菌、真菌OTUs數量,FZ處理降低了土壤細菌、真菌微生物OTUs數量。
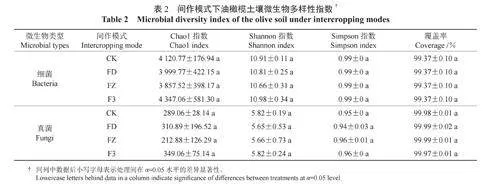
2.2.2微生物α多樣性指數
α多樣性指數是反映微生物群落豐富度、多樣性和均勻度的綜合指標。從表2中多個多樣性指數來看,4種間作模式下細菌和真菌多樣性指數間無顯著差異,細菌的Chao1、Shannon指數均大于真菌,各處理細菌和真菌多樣性指數變化與OTUs數量變化趨勢基本一致。根據細菌群落豐富度和多樣性指數由大到小排列,各處理依次為F3、CK、FD、FZ;根據真菌群落豐富度和多樣性指數由大到小排列,各處理依次為F3、FD、CK、FZ。
2.2.3微生物群落組成
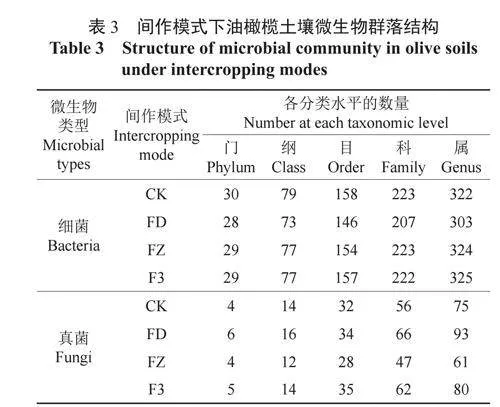
4種間作模式下油橄欖土壤細菌群落中共檢測到33門83綱165目232科348屬,真菌群落中共檢測出7門17綱42目93科154屬。由表3可知,細菌群落組成的數量在各分類水平上均明顯高于真菌群落組成數量。根據細菌群落組成在各分類水平上由多到少排列,各處理依次為CK、F3、FZ、FD;根據真菌群落組成在各分類水平上由多到少排列,各處理依次為FD、F3、CK、FZ。
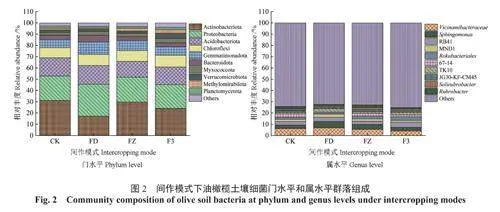
圖2~3分別展示了4種間作模式下油橄欖土壤細菌和真菌在門和屬分類學水平上相對豐度的變化規律,1種色塊代表1個物種,色塊的長度表示該物種的相對豐度。由圖2A可知,4種間作模式下油橄欖土壤細菌門類相對豐度在前10位的菌群基本相同,分別為放線菌門Actinobacteriota、變形菌門Proteobacteria、酸桿菌門Acidobacteriota、綠彎菌門Chloroflexi、芽單胞菌門Gemmatimonadota、擬桿菌門Bacteroidota、黏球菌門Myxococcota、疣微菌門Verrucomicrobiota、Methylomirabilota、浮游球菌門Planctomycetota。4種間作模式下油橄欖土壤中優勢菌門均為放線菌門(17.09%~31.03%)、變形菌門(21.27%~28.46%)、酸桿菌門(14.11%~16.60%)、綠彎菌門(8.95%~10.38%)、芽單胞菌門(7.06%~11.01%)。4種間作模式下放線菌門相對豐度由高到低分別為CK(31.03%)、FZ(29.72%)、F3(24.02%)、FD(17.09%),變形菌門相對豐度由高到低分別為FD(28.46%)、FZ(22.08%)、CK(21.84%)、F3(21.27%),酸桿菌門相對豐度由高到低分別為FD(16.60%)、CK(16.13%)、F3(15.58%)、FZ(14.11%)。4種間作模式下油橄欖土壤中細菌在屬分類水平上的相對豐度具有一定差異(圖2B),前3位優勢菌屬分別為Vicinamibacteraceae(4.03%~6.80%)、鞘氨醇單胞菌屬Sphingomonas(2.97%~5.64%)、RB41(2.75%~4.67%),4種間作模式下Vicinamibacteraceae相對豐度由高到低分別為FD(6.80%)、CK(6.30%)、FZ(5.45%)、F3(4.03%),鞘氨醇單胞菌屬相對豐度由高到低分別為FD(5.64%)、FZ(4.40%)、F3(3.20%)、CK(2.97%),RB41相對豐度由高到低分別為F3(4.67%)、FD(3.63%)、CK(3.16%)、FZ(2.75%)。綜上,間作后各模式下土壤細菌在門、屬水平群落物種組成上與CK相似,但間作后各菌群的相對豐度存在差異。
由圖3A可知,間作模式下土壤優勢真菌門類群主要為子囊菌門Ascomycota(88.42%~73.18%)、擔子菌門Basidiomycota(2.87%~13.45%)。同時發現:被孢霉門Mortierellomycota為FZ間作模式下的優勢菌門,相對豐度達6.03%,在其他模式中豐度較小。各間作模式按照土壤中子囊菌門的相對豐度由大到小排序,依次為FD(88.42%)、FZ(85.66%)、F3(82.20%)、CK(73.18%);按照擔子菌門的相對豐度由大到小排序,依次為CK(13.45%)、F3(10.07%)、FD(6.21%)、FZ(2.87%)。由圖3B可知,真菌屬水平類群主要為鐮刀菌屬Fusarium、青霉屬Penicillium、曲霉屬Aspergillus、古根菌屬Archaeorhizomyces、尖囊菌屬Subulicystidium、黑星菌屬Venturia、小囊菌屬Microascus、被孢霉屬Mortierella、阿澤拉霉屬Arxiella、枝孢屬Cladosporium。各間作模式下的優勢菌屬不同,CK模式下古根菌屬(5.46%)和尖囊菌屬(4.29%)是優勢菌屬,FD間作模式下鐮刀菌屬(11.32%)、曲霉屬(10.52%)和黑星菌屬(6.97%)為優勢菌屬,FZ間作模式下優勢菌屬為鐮刀菌屬(8.86%)和青霉屬(14.36%),F3間作模式下土壤中優勢菌屬為古根菌屬(9.26%)、青霉屬(7.83%)、小囊菌屬(6.13%)和尖囊菌屬(5.66%)。綜上,4種模式下土壤真菌在門水平上子囊菌門均為絕對優勢門,且間作后子囊菌門的相對豐度較CK均有所增加,在屬水平上各模式間優勢屬組成及相對豐度差異較大,3種間作模式均使部分土壤功能真菌微生物的相對豐度明顯增加,其中FD和FZ模式增加了常見致病菌屬鐮刀菌屬的相對豐度,病原菌屬黑星菌屬、枝孢屬在FD模式下的相對豐度也較高,說明該間作模式可能會增加土壤病害侵擾的概率。
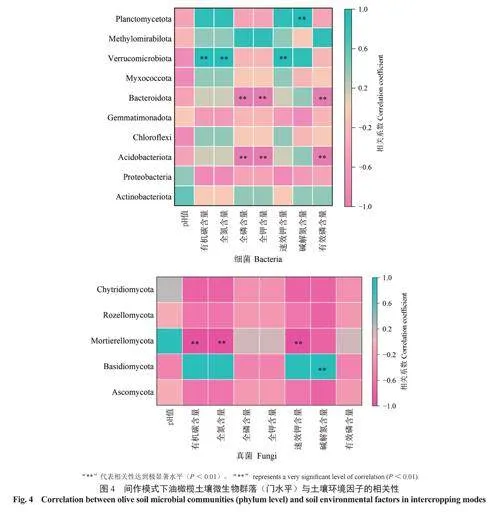
2.3不同間作模式下土壤微生物群落組成與土壤環境因子的相關性
為了進一步了解土壤環境因子與土壤細菌、真菌群落的關系,將土壤中主要優勢細菌和真菌菌群的相對豐度與土壤環境因子進行相關性分析。由土壤細菌相對豐度與土壤環境因子的Spearman相關性熱圖(圖4)可知,在門分類水平上,酸桿菌門和擬桿菌門的相對豐度與全磷含量、全鉀含量、有效磷含量極顯著負相關,疣微菌門的相對豐度與有機碳含量、全氮含量、速效鉀含量極顯著正相關,浮游球菌門的相對豐度與堿解氮含量極顯著正相關。由土壤真菌相對豐度與土壤環境因子的Spearman相關性熱圖(圖4)可知,擔子菌門Basidiomycota的相對豐度與堿解氮含量極顯著正相關,被孢霉門的相對豐度與有機碳含量、全氮含量、速效鉀含量極顯著負相關。同時,土壤pH值與細菌、真菌各類菌群的相對豐度均無顯著相關性。綜上,細菌和真菌群落結構與土壤環境因子有著密切的相關性,但影響其土壤微生物群落結構的主要環境因子存在差異。
3討論與結論
3.1間作模式對土壤養分元素含量的影響
大量研究結果證明,間作會對土壤環境因子產生影響[17-19]。本研究中,相較于CK,3種間作模式均對土壤養分含量產生了影響,FD間作模式下土壤速效鉀、有效磷、堿解氮含量顯著降低。滇黃精是多年生宿根草本植物,以地下根狀莖為主要收獲對象。在本研究中,自2018年起開始用滇黃精進行間作試驗,其生長階段對速效鉀、有效磷、堿解氮等養分有不同的需求,并且會將大量養分向根莖積累[20],導致土壤養分含量下降。這一現象與嚴毅等[10]的研究結果相吻合,他們發現在油橄欖林下種植百合后,土壤中營養成分含量均明顯降低,表明百合生長對有機質、氮、磷、鉀等養分有不同程度的吸收需求。因此,在間作滇黃精期間,應注意其與油橄欖之間的養分競爭,注意加強施肥管理。FZ間作模式顯著增加了土壤的全鉀、有效磷含量,但有機碳、堿解氮、速效鉀含量在間作后顯著降低。紫花苜蓿草為深根性多年生牧草,有研究結果表明豆科作物具有較強的質子釋放能力,其根系對土壤中難溶性磷的活化和其他作物吸收磷有促進作用[21-22],進而提高了作物的磷含量。大量生產實踐結果表明,紫花苜蓿草的根瘤有固氮的作用[23-24],可在一定程度上增加土壤中氮含量。本研究中可能由于紫花苜蓿草間作年限較長,其根系會分泌一些不易降解的酚酸類物質,這些物質累積在土壤中影響其生長[25],從而影響養分吸收與轉化,同時間作后紫花苜蓿草與油橄欖樹存在養分爭奪的現象,導致土壤中氮含量顯著降低。F3間作模式下土壤全鉀含量顯著增加,除堿解氮外其他養分含量相較于CK均有所增加,其原因是多種牧草根系在地下的分布存在生態位差異[26],可充分協調水、養、氣的協同[27],這與王雪等[28]、姜黎等[29]、張永亮等[30]的研究結果相似,同時混種間作作物占據了更多生態位,增加了群落的穩定性,其凋落物累積量也會增加,從而回歸土壤的養分增多[31]。
3.2間作模式對土壤微生物多樣性和群落組成的影響
間作對土壤微生物多樣性的改變通常與土壤環境、作物種類、間作年限等多重因素相關。OTU分析與α多樣性分析結果顯示,間作后細菌和真菌多樣性指數無顯著差異,但F3間作模式在一定程度上增加了土壤中細菌、真菌的OTUs數量,這也反映了多草種間作有利于提高土壤微生物多樣性,3種間作處理下細菌群落物種組成與CK相似,均以放線菌門、變形菌門、酸桿菌門為優勢細菌菌門,這些菌門在油橄欖土壤中的優勢地位已有報道[32],但間作后各菌群的相對豐度有一定變化,由于不同植物的根系分泌物和凋落物理化性質不同,其分解時釋放的養分具有差異,進而導致各處理中優勢菌門的相對豐度存在差異[33-34]。子囊菌門、擔子菌門、被孢霉門為真菌優勢菌門,這與許彥明等[35]的研究結果一致。子囊菌門作為真菌界種類最多的門,在土壤有機質的降解中具有重要作用[36]。真菌門水平上,3種間作處理相較于CK均增加了子囊菌門的相對豐度,這可能與間作作物的分泌物、凋落物為其提供營養與能量有關。被孢霉門具有溶磷性[37],被孢霉門為FZ間作模式下土壤的優勢菌門,這與FZ間作模式顯著增加了土壤中有效磷含量的分析結果一致。在真菌屬水平上,各模式間優勢屬組成及相對豐度的差異均較大,間作均增加了其他功能真菌群落的相對豐度。其中間作滇黃精(FD)和紫花苜蓿草(FZ)后鐮刀菌屬的相對豐度明顯增加,鐮刀菌屬為常見的根腐病病原菌,并在多種植物中被檢出[38-39],因此間作滇黃精與紫花苜蓿草后可能出現病害侵擾。相較于CK,F3模式下土壤中古根菌屬、青霉屬、小囊菌屬等有益微生物的相對豐度明顯增加,這種間作模式有利于提高土壤有益功能真菌的豐富度,多草種間作通常對土壤溫度、養分等土壤環境因子有明顯影響,從而對微生物群落種類和結構產生直接或間接影響[40],導致群落組成結構產生差異。
3.3土壤微生物群落組成與土壤環境因子的關系
土壤微生物對環境極為敏感,土壤類型和理化性質的改變會引起土壤微生物群落組成發生變化,主要菌群的豐度和結構也不同[41]。土壤理化性質對土壤微生物群落的影響較為復雜,目前尚無完全一致的結論。顏彩繽等[42]通過對3種菠蘿蜜間作模式進行研究,發現放線菌門與土壤全鉀、全氮、全磷含量均呈現極顯著或顯著負相關;何升然等[43]通過對紫花苜蓿與甜高粱間作進行研究,發現酸桿菌門和疣微菌門與土壤pH值呈現極顯著正相關,與土壤堿解氮含量呈現極顯著負相關。李敏等[44]發現氮和磷含量能夠顯著影響根際土壤真菌群落,氮元素的轉化和循環與土壤真菌有著密切的關系,磷元素在真菌的繁殖和生長中也發揮了重要的作用。相關性分析結果表明,土壤有機碳、全氮、全磷、全鉀、速效鉀、堿解氮、有效磷的含量與部分細菌門的相對豐度有極顯著相關性,土壤有機碳、全氮、速效鉀、堿解氮的含量與部分真菌門的相對豐度有極顯著相關性,表明養分元素對這些微生物的生長和代謝具有促進或抑制作用,間作通過改善土壤理化性質為土壤部分菌群提供了適宜的營養環境,進而使得土壤微生物數量、活性及群落組成發生變化[45-46],李承臻等[47]的研究結果表明,在荔枝園間作柱花草可以改變土壤微生物群落結構,微生物群落相對豐度的變化與間作柱花草后氮、磷、鉀等土壤養分含量的變化有關,這與本研究結果一致。同時部分養分含量與某些細菌、真菌存在抑制效應,土壤中某些養分過量可能抑制這些微生物的生長或導致其處于競爭劣勢[48]。
油橄欖×滇黃精(FD)、油橄欖×紫花苜蓿草(FZ)、油橄欖×紫花苜蓿草×黑麥草×金雞菊(F3)3種間作模式實施5a后,土壤化學性質均發生了改變。FD模式和FZ模式間作后土壤中部分養分含量顯著降低,間作作物與油橄欖存在養分爭奪現象,F3模式提高油橄欖林地土壤養分含量的效果最佳。3種間作模式對土壤微生物多樣性影響較小,但可誘導土壤細菌、真菌群落結構發生變化,以真菌群落結構的變化最為明顯。3種間作模式均明顯增加了部分土壤真菌有益微生物的相對豐度,但FD和FZ模式增加了常見部分土壤致病菌屬的相對豐度,F3間作模式對維持和促進土壤生態系統良性發展具有積極影響。土壤細菌與真菌群落結構變化受土壤化學性質驅動,土壤養分含量對部分細菌和真菌群落組成和豐度有重要影響。綜上,油橄欖×紫花苜蓿草×黑麥草×金雞菊(F3)多草種間作模式優于油橄欖×滇黃精(FD)和油橄欖×紫花苜蓿草(FZ)間作模式。本研究中僅測定分析了間作模式下油橄欖林地表層土壤養分含量指標和土壤微生物多樣性及群落結構變化的差異,未涉及不同深度土壤的養分含量和微生物群落變化,后續將進一步研究間作對油橄欖林地不同土層微生物群落和養分變化的影響,同時間作作物對油橄欖樹養分吸收及果實產量和品質的影響程度也有待進一步探究。
猜你喜歡
熱帶農業科學(2016年11期)2017-01-21 14:59:21
熱帶農業科學(2016年11期)2017-01-21 14:57:08
河北漁業(2015年12期)2015-12-21 14:37:31
河北漁業(2015年11期)2015-11-18 16:39:54
河北漁業(2015年11期)2015-11-18 16:37:49
河北漁業(2015年10期)2015-10-15 11:14:57
湖北農業科學(2015年18期)2015-10-13 21:34:01
河北漁業(2015年7期)2015-07-21 11:06:52
河北漁業(2015年6期)2015-06-24 11:04:37
安徽農學通報(2015年5期)2015-03-23 23:51:20
