苜蓿基因組密碼子偏好性分析
2024-12-31 00:00:00李琳徐明志劉燕蓉張萬軍
草地學報 2024年9期


摘要:物種基因的密碼子使用具有偏好性,能避免氨基酸的錯誤摻入。苜蓿(Medicago)密碼子的偏好性尚未見報道。為揭示苜蓿屬基因組密碼子偏好性特點,我們利用軟件分析四種苜蓿基因編碼區密碼子組成、相對同義密碼子使用頻率(RSCU)等參數探討密碼子偏好性特點,同時用多種方法分析造成苜蓿密碼子偏好性的影響因素,計算確定苜蓿的最優密碼子,最后通過聚類分析研究物種間密碼子使用偏好性進化關系。結果表明:四種苜蓿的密碼子偏好性相對較弱,其中‘中苜1號’最弱,苜蓿密碼子第三位偏愛使用A/T;四種苜蓿密碼子偏好性受自然選擇壓力的影響大,最優密碼子多以A/T結尾;四種苜蓿的偏好性相似,且與煙草(Nicatiana tabacum L.)、番茄(Solanum lycopersicum L.)偏好性接近。本研究為苜蓿密碼子優化應用提供了理論基礎,特別是通過優化可以提高Cas9蛋白表達量促進基因編輯。
關鍵詞:苜蓿;密碼子偏好性;最優密碼子;基因編輯;自然選擇
中圖分類號:Q756""" 文獻標識碼:A""""" 文章編號:1007-0435(2024)09-2695-12
doi:10.11733/j.issn.1007-0435.2024.09.003
引用格式:
李" 琳, 徐明志, 劉燕蓉,等.苜蓿基因組密碼子偏好性分析[J].草地學報,2024,32(9):2695-2706
LI Lin, XU Ming-zhi, LIU Yan-rong,et al.Codon Bias Analysis of Medicago Genome[J].Acta Agrestia Sinica,2024,32(9):2695-2706
Codon Bias Analysis of Medicago Genome
LI Lin, XU Ming-zhi, LIU Yan-rong, ZHANG Wan-jun*
(College of Grassland Science and Technology,China Agricultural University,Beijing 100193,China)
Abstract:The genes codon usage of species has a bias,which can avoid the wrong incorporation of amino acids. The bias of Medicago codon has not been reported. In order to reveal the codon bias characteristics of Medicago genome,we used software to analyze the codon composition of the coding region,the relative synonymous codon use frequency (RSCU),and other parameters of four Medicago species to explore the codon bias characteristics. At the same time,we used various methods to analyze the influencing factors causing the codon bias of Medicago,and calculated the optimal codon of Medicago. Finally,the evolutionary relationship of codon usage bias among species was studied by cluster analysis. The results showed that the codon bias of four Medicago species was relatively weak,of which ‘Zhongmu No.1’ was the weakest,and the third position of Medicago codon prefered A/T;The codon bias of four Medicago species was greatly affected by natural selection pressure,and most of the optimal codons ended in A/T. The bias of four Medicago species was similar and close to Nicatiana tabacum L.and Solanum lycopersicum L.. This study provides a theoretical basis for the application of Medicago codon optimization,especially through the optimization to improve the expression of Cas9 protein to promote gene editing.
Key words:Medicago;Codon usage bias;Optimal codon;Gene editing;Natural selection
收稿日期:2024-02-21;修回日期:2024-05-02
基金項目:科技創新2030苜蓿育種項目(2022ZD040120301)資助
作者簡介:
李琳(1997-),女,漢族,山西陽泉人,碩士研究生,主要從事苜蓿轉基因育種研究,E-mail:17824230559@163.com;*通信作者Author for correspondence,E-mail:wjzhang@cau.edu.cn
遺傳密碼子具有簡并性和偏好性。簡并性可以降低DNA突變引起的翻譯誤差,保持物種的穩定性;偏好性避免了氨基酸的錯誤摻入以減少遺傳信息的加工誤差[1]。生物體在進化和適應環境的過程中,如果沒有任何基因突變和選擇壓力,同義密碼子的選擇是隨機的。密碼子使用偏好性是生物體偏愛使用某些同義密碼子的現象,在植物、動物和微生物中普遍存在[2]。不同物種的基因在密碼子使用頻率上存在差異[3-5],甚至,同一物種的不同品種基因的密碼子使用頻率上也存在較大差異[6],物種密碼子使用的偏好性與同義突變引起的“沉默突變”不同[7-8]。密碼子的使用模式會影響mRNA折疊、翻譯延伸和蛋白折疊,進而影響基因表達[9],尤其在單子葉植物中GC含量與基因表達量密切相關[10]。基因密碼子使用的偏好性形成原因主要有基因變異、選擇、隨機漂移[11]。此外,其他因素如基因表達水平[12]、基因長度、重組率、組分偏性(GC含量)、環境壓力、種群大小等也會影響密碼子的使用。
為提高基因表達,在優化密碼子方面已取得了很大進展。其中,生物制藥領域同義密碼子突變可以用于增加蛋白表達產量。如陳徵婷等[13]研究發現按照大腸桿菌密碼子優化GBS表面蛋白Lrrc,在免疫原性保持的情況下,有效提高了其在大腸桿菌中的表達量,為制備和應用LrrG基因工程疫苗提供了指導。根據植物體內密碼子編碼偏好性,對SpCas9進行了密碼子優化,使其能在植物體內穩定表達[14]。基因編輯技術可以實現對重要基因的定向改造,但受限于基因編輯效率,應用上常難以達到預期效果。近年來,單子葉植物中所有獲得突變植株的報道中所采用的Cas9基因都是經過密碼子優化過的[15]。基于香蕉密碼子偏好性優化Cas9密碼子顯著提高了Cas9蛋白在香蕉中的表達,原生質體轉化后基因編輯效率提高了4倍[16]。針對鐮刀菌優化的Cas9已經成功實現了真菌的編輯[17]。
紫花苜蓿(Medicago sativa L.)應用廣泛,有“牧草皇后”的美稱[18],培育高產、抗逆性強的紫花苜蓿一直是其遺傳改良的目標[19]。然而,紫花苜蓿育種上因遺傳背景復雜,傳統育種存在周期長的問題[20]。分子設計育種一定程度上緩解了這些問題,但外源基因的表達水平常會受到密碼子偏好性的影響[21]。另外,CRISPR基因編輯技術應用前景廣大,而Cas9蛋白的表達量在其中發揮著重要作用,但其在苜蓿中密碼子偏好性尚未見報道。
本研究以4種不同苜蓿的基因組編碼序列為研究對象,探討密碼子組成、相對同義密碼子使用頻率等分析密碼子使用特點,并結合多種分析方法揭示了苜蓿密碼子偏好性的影響因素,還從進化關系上解析不同物種密碼子使用上的相似程度。研究結果為提高基因在紫花苜蓿中表達[22]、預測[23]及外源受體的選擇提供了參考[24],特別是紫花苜蓿偏好性對提高Cas9蛋白表達,提高編輯效率意義重大。
1" 材料與方法
1.1" 序列數據
從https://modms.lzu.edu.cn/中獲取蒺藜苜蓿(A17)、‘新疆大葉’‘中苜1號’以及‘中苜4號’基因組數據,獲取所有編碼蛋白的基因序列,使用Excel2019篩選以ATG為起始密碼子,以TAA、TAG或TGA為終止密碼子的基因。同時,剔除CDS長度小于300 bp的基因作為分析樣本。
1.2" 密碼子組成分析
密碼子組成分析由CodonW 1.4.2軟件對四種苜蓿的基因編碼序列分析完成,計算得到GC1,GC2,GC3值為密碼子三個位置上的GC含量,同時獲得同義密碼子使用頻率(RSCU)。顯著性通過雙向秩方或然性檢驗進行評估,檢驗標準為Plt;0.01[23]。ENc值大小反映密碼子偏好性強弱,按照慣例以35作為偏好性強弱的區分標準,高于此標準代表基因組密碼子偏好性較弱,反之,密碼子偏好性較強。
1.3" 相對同義密碼子使用頻率分析
相對同義密碼子使用頻率(RSCU)在實際使用中較為廣泛,表示密碼子的實際使用值與理論使用值之間的比值。當RSCU=1表示密碼子沒有偏好性,RSCUgt;1,RSCUlt;1分別代表該密碼子的使用頻率高于、低于同義密碼子。計算公式為:RSCUij=Xji1ni∑nij=1Xij,其中Xji是編碼第i個氨基酸的第j個密碼子的出現次數;ni是編碼第i個氨基酸的同義密碼子的數量[7]。
1.4"" ENc-plot分析
有效密碼子數(Effective number of codon,ENc)是反映同義密碼子非均衡使用偏好程度的重要指標。ENc-plot分析以ENc值為縱坐標,GC3為橫坐標繪制,ENc-plot主要分析堿基組成對密碼子偏好性的影響。標準曲線的計算公式如下:ENc=2+GC3+29(GC3)2+(1-GC3)2 [25],沿標準曲線分布或在其附近表示該基因密碼子偏好性僅受突變影響,基因落在標準曲線下方較遠的位置表示該基因密碼子偏好性受到選擇壓力的影響。
1.5" 中性繪圖分析
統計密碼子各位置GC含量,GC12表示GC1和GC2的平均值。以GC3、GC12分別為橫、縱坐標繪制散點圖,圖中每個散點表示一個基因。位于對角線上的基因代表僅受突變壓力的影響。
1.6 PR2-plot繪圖分析
以A3/(A3+T3)值為縱坐標,G3/(G3+C3)為橫坐標,繪制散點圖。如果DNA的兩條互補鏈之間的突變和選擇沒有偏差,則A=T和G=C,此時其他點和中心點之間的角度和距離記為偏移的程度和方向。
1.7" 最優密碼子分析
依據ENc值大小對各基因進行排序,從這一排列的兩端各取基因樣本總數的5%構建高表達基因庫和低表達基因庫,計算這兩組基因的相對密碼子使用度,并進行卡方檢驗,確定最優密碼子,并用*標出[26]。
1.8" 基于密碼子使用偏好性的聚類分析
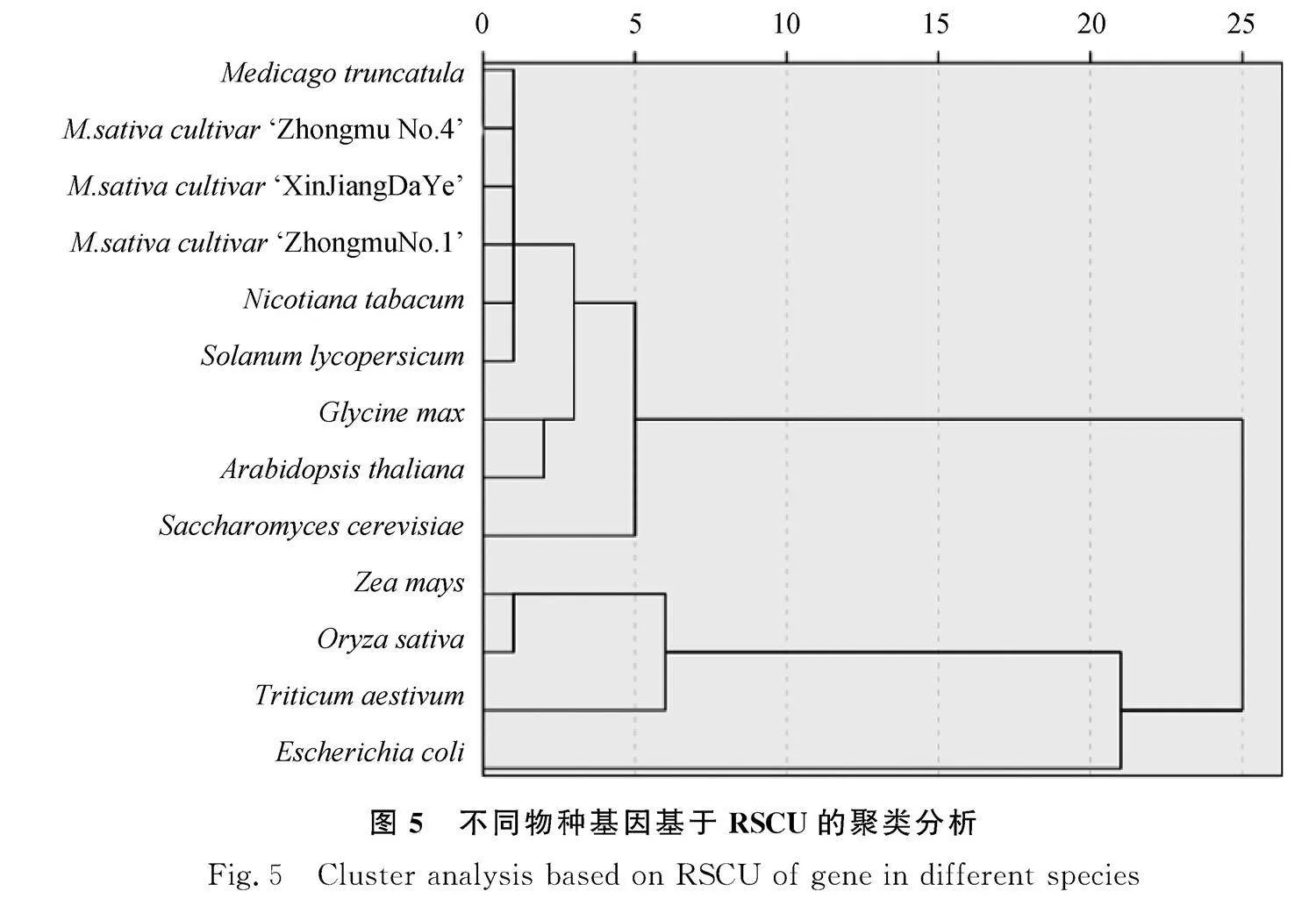
從Codon Usage Database (http://www.kazusa.or.jp/codon/) 中獲得酵母(Saccharomys cerevisiae)、大腸埃希菌(Escherichia coli)、擬南芥(Arabidopsis thaliana)、煙草(Nicotiana tabacum)、玉米(Zea mays)、番茄(Solanum lycopersicum)、水稻(Oryza sativa)、大豆(Glycine max)、小麥(Triticum aestivum)等植物的密碼子使用頻率進行比較。并通過CUSP計算蒺藜苜蓿(M. truncatula)、‘新疆大葉’(M. sativa cultivar ‘XinJiangDaYe’)、‘中苜1號’(M. sativa cultivar ‘Zhongmu No. 1’)、‘中苜4號’(M. sativa cultivar ‘Zhongmu No. 4’)的基因組密碼子使用頻率。利用SPSS 27.0對上述物種進行59種密碼子偏好性分析(其中除去起始密碼子ATG、色氨酸TGG以及終止密碼子TAA、TAG、TGA)。采用系統聚類法進行[27]。
2" 結果與分析
2.1" 密碼子組成分析
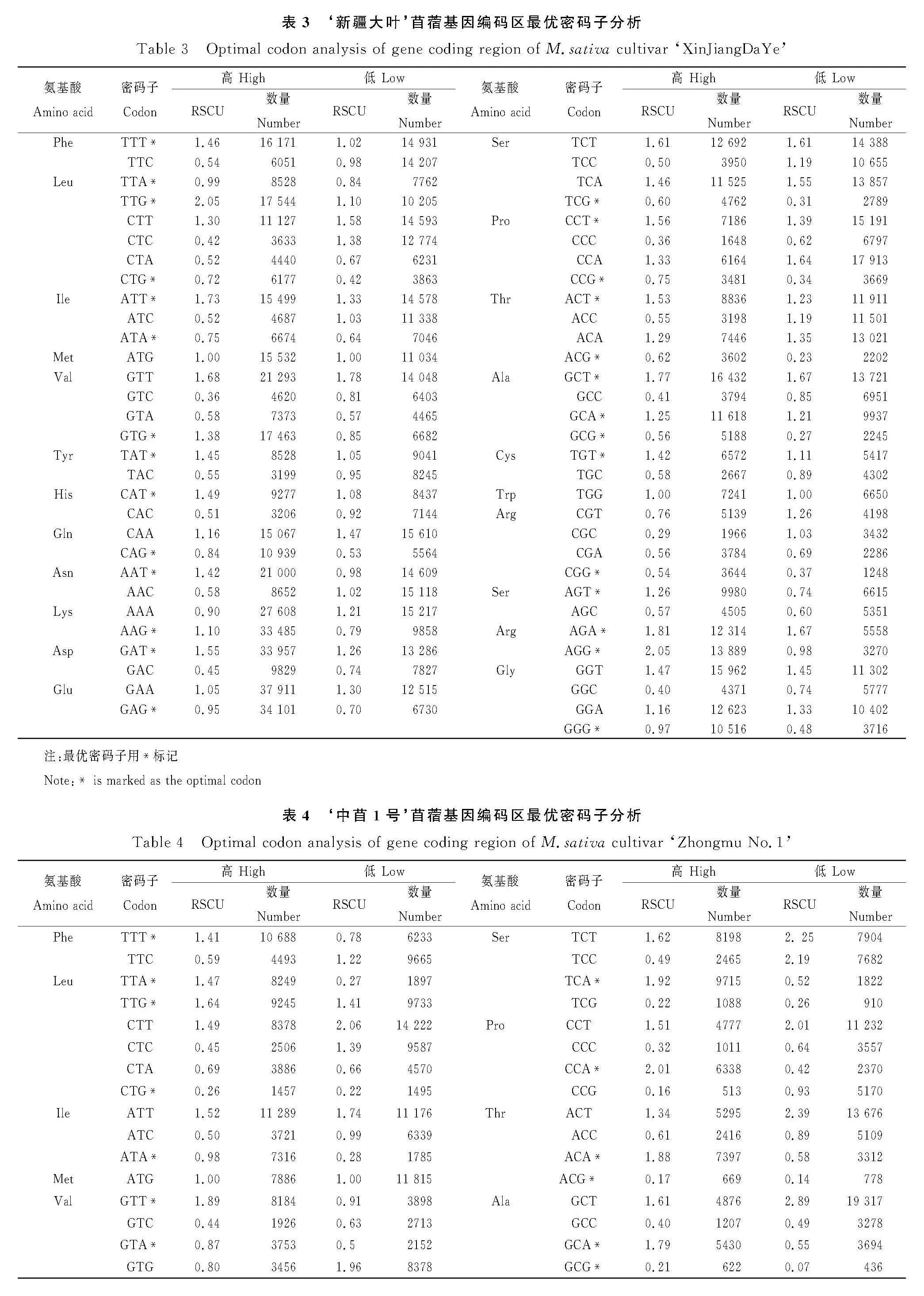
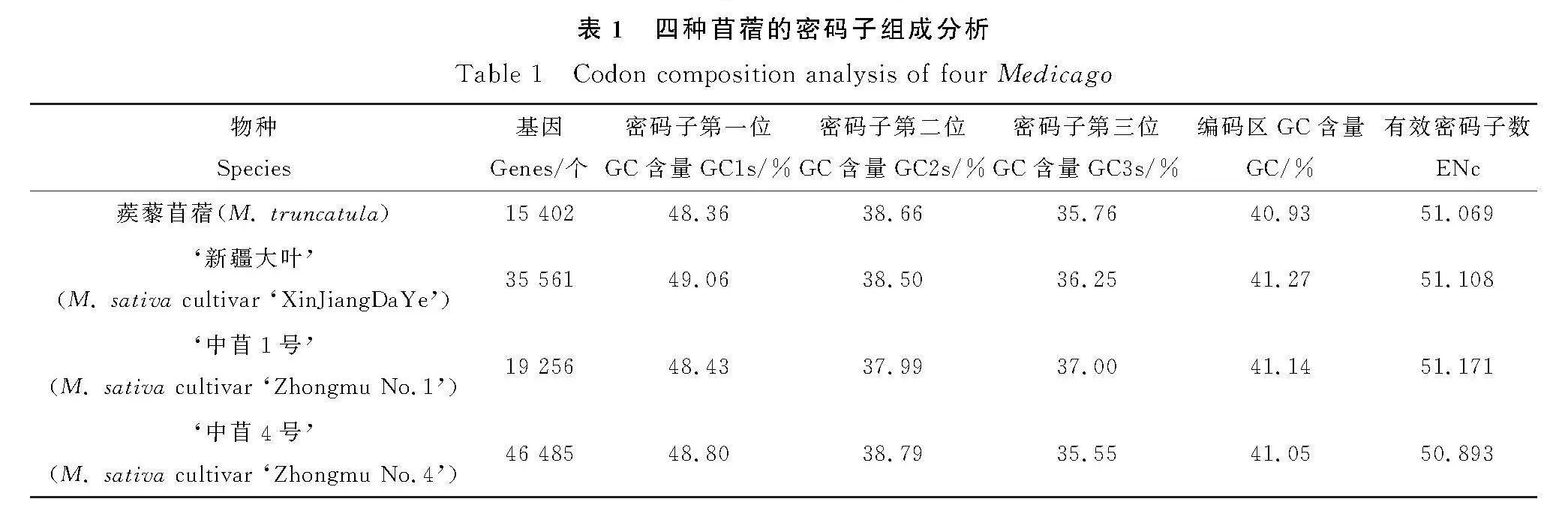
我們選取蒺藜苜蓿、‘新疆大葉’‘中苜1號’和‘中苜4號’共四種苜蓿的基因編碼區序列,按照一定的規則篩選后進行分析[28],不同品種苜蓿基因編碼區的GC含量非常接近(表1),均為41%左右,密碼子各位置的GC含量不同,但呈現出一定的共性:GC1gt;GC2gt;GC3。密碼子第三位的堿基組成對編碼基因的密碼子使用偏好性影響較大,四種苜蓿GC3s含量為35.55%~37.00%之間,說明其偏好以A/T為第3位核苷酸的密碼子。有效密碼子數(ENc)是評價基因組整體密碼子偏好性中具有重要參考價值的參數,參考值為20~61,值越小,表明密碼子偏好性更強[29]。我們使用軟件Codon W的分析結果顯示,ENc值排序為:‘中苜1號’gt;‘新疆大葉’gt;蒺藜苜蓿gt;‘中苜4號’,均大于50,說明苜蓿基因編碼區的密碼子偏好性程度較弱,最弱的是‘中苜1號’。
2.2" 同義密碼子的使用頻率分析
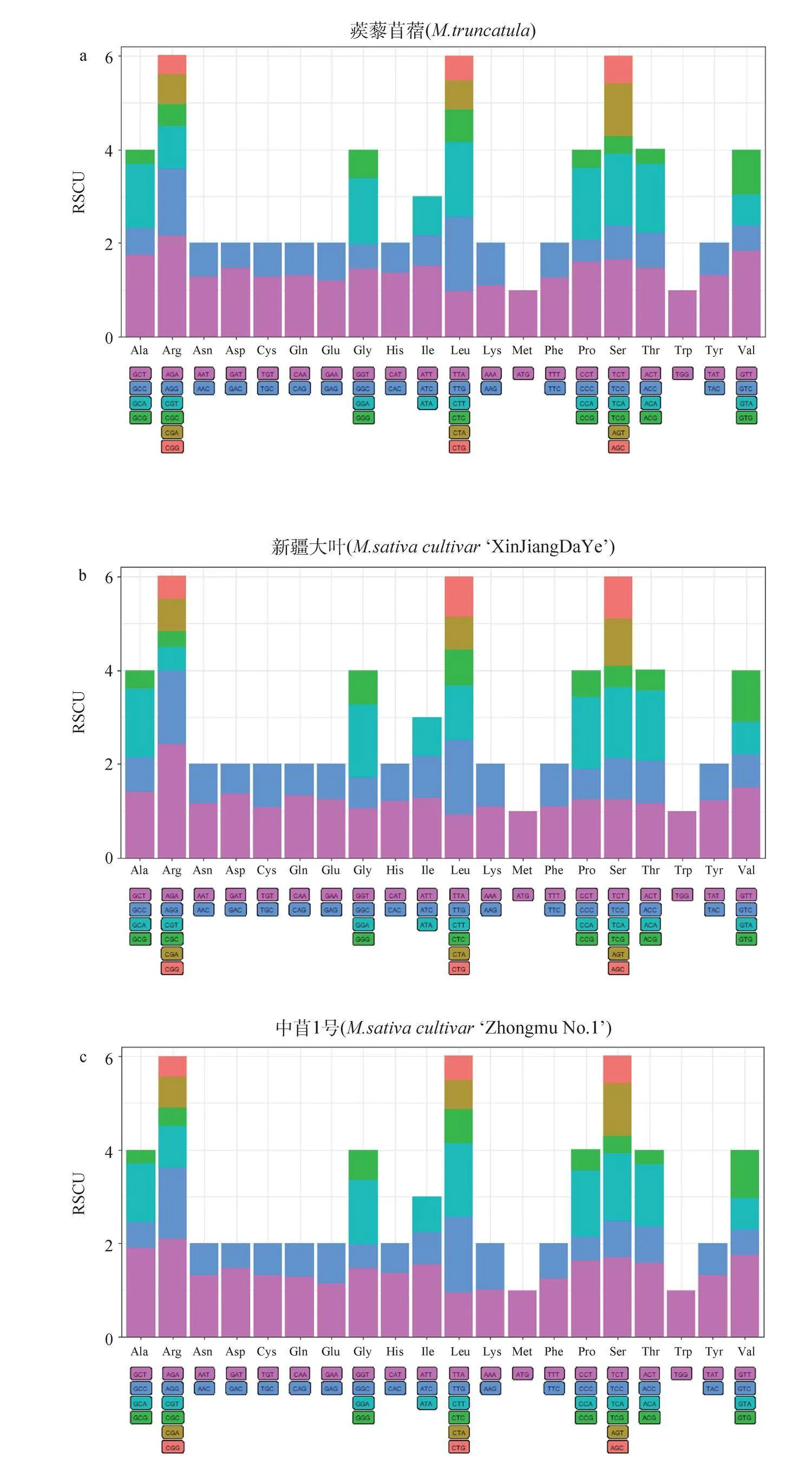
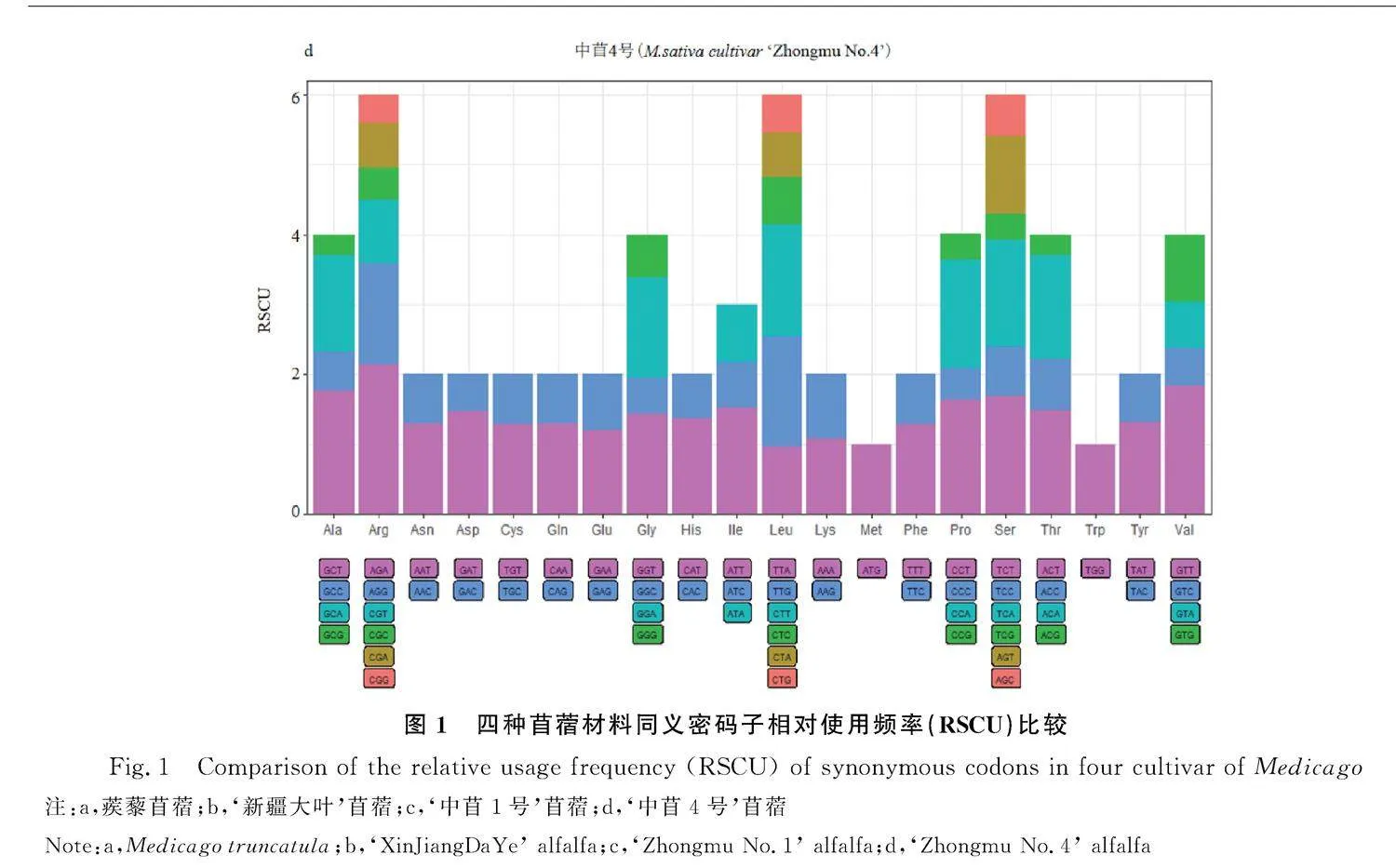
除了甲硫氨酸(Met)和色氨酸(Trp)對應一種密碼子,其余氨基酸對應2~6種密碼子不等。我們對四種苜蓿中20種氨基酸各自對應同義密碼子的相對使用頻率進行比較發現四種苜蓿中相對使用頻率高的密碼子的第三位均以A/T結尾,這些密碼子成為苜蓿的偏愛密碼子。相反,以G/C結尾的密碼子為非偏愛密碼子。蒺藜苜蓿、‘新疆大葉’‘中苜1號’‘中苜4號’中RSCU值最高的密碼子均為AGA(編碼精氨酸),RSCU值分別為2.16,2.43,2.09,2.14(圖1)。
2.3" ENc-plot分析
ENc-plot分析結果顯示在四種苜蓿中均只有少部分點位于標準曲線附近,大部分點位于標準曲線下方且距離遠,說明密碼子的使用模式主要受環境影響大,那些分布在標準曲線上的密碼子使用偏性還受突變的影響;值得注意的是,蒺藜苜蓿中的1個基因和‘中苜4號’中的2個基因位于標準曲線下方且距離相對最遠,說明這少部分基因受環境壓力影響更大(圖2)。綜上,在四種苜蓿基因編碼區的密碼子使用的偏好性受自然選擇壓力的影響大于突變的影響。
2.4" 中性繪圖分析
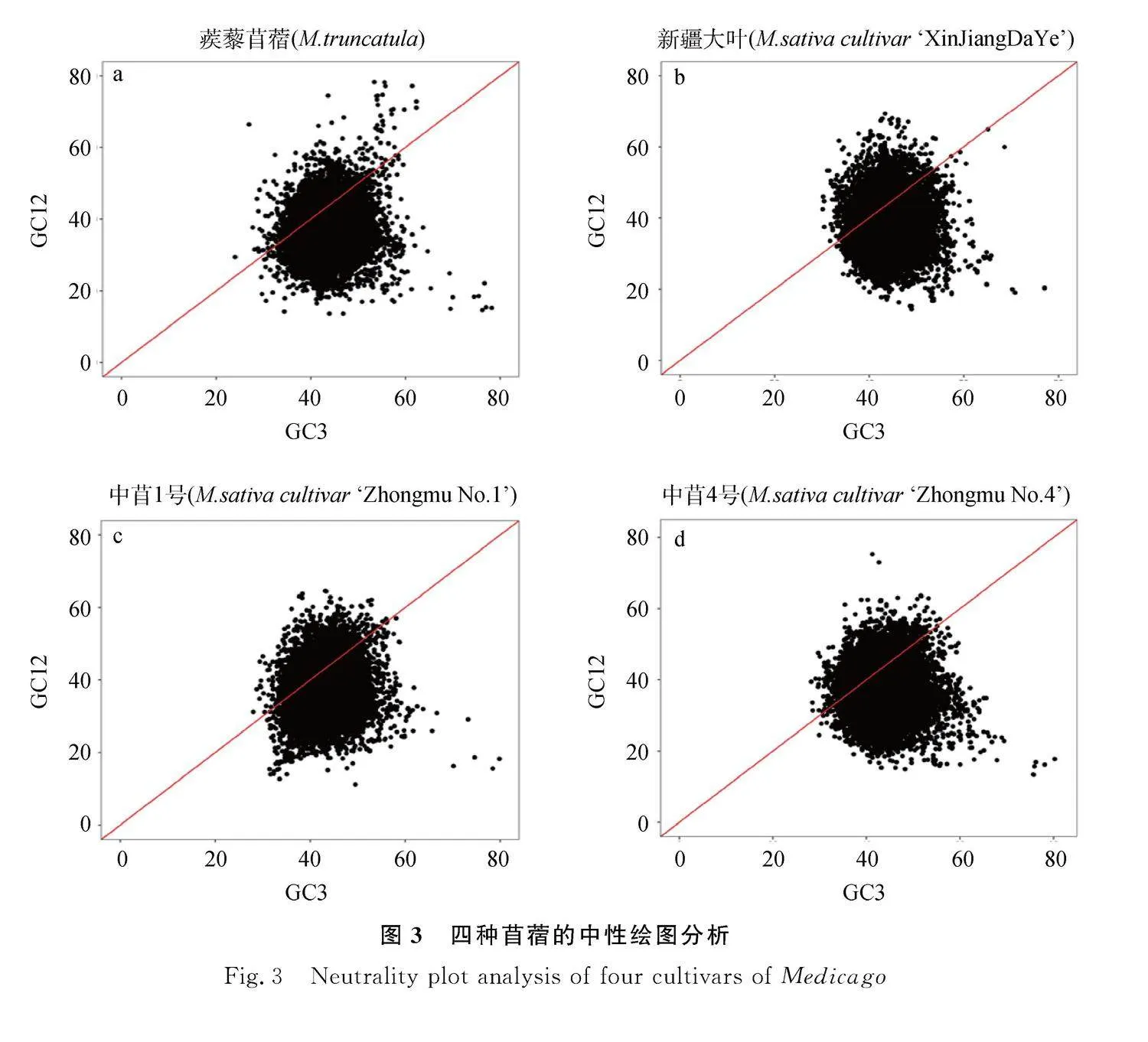
中性繪圖反映GC12和GC3之間的相關關系,反映苜蓿密碼子使用的突變選擇平衡。利用MEGA軟件進行中性繪圖分析,結果顯示:蒺藜苜蓿、‘新疆大葉’‘中苜1號’‘中苜4號’大部分基因均在中性圖對角線偏右位置,表明GC12與GC3相關性弱,密碼子偏好性受自然選擇影響大于堿基突變,蒺藜苜蓿和‘中苜4號’少數基因的GC3值達到80%,這些基因是基因組中受自然選擇最明顯的,與ENc-plot結果一致。從品種來看,‘中苜1號’和‘中苜4號’可能受選擇的影響大于另外兩個材料(圖3)。
2.5" PR2-Plot分析
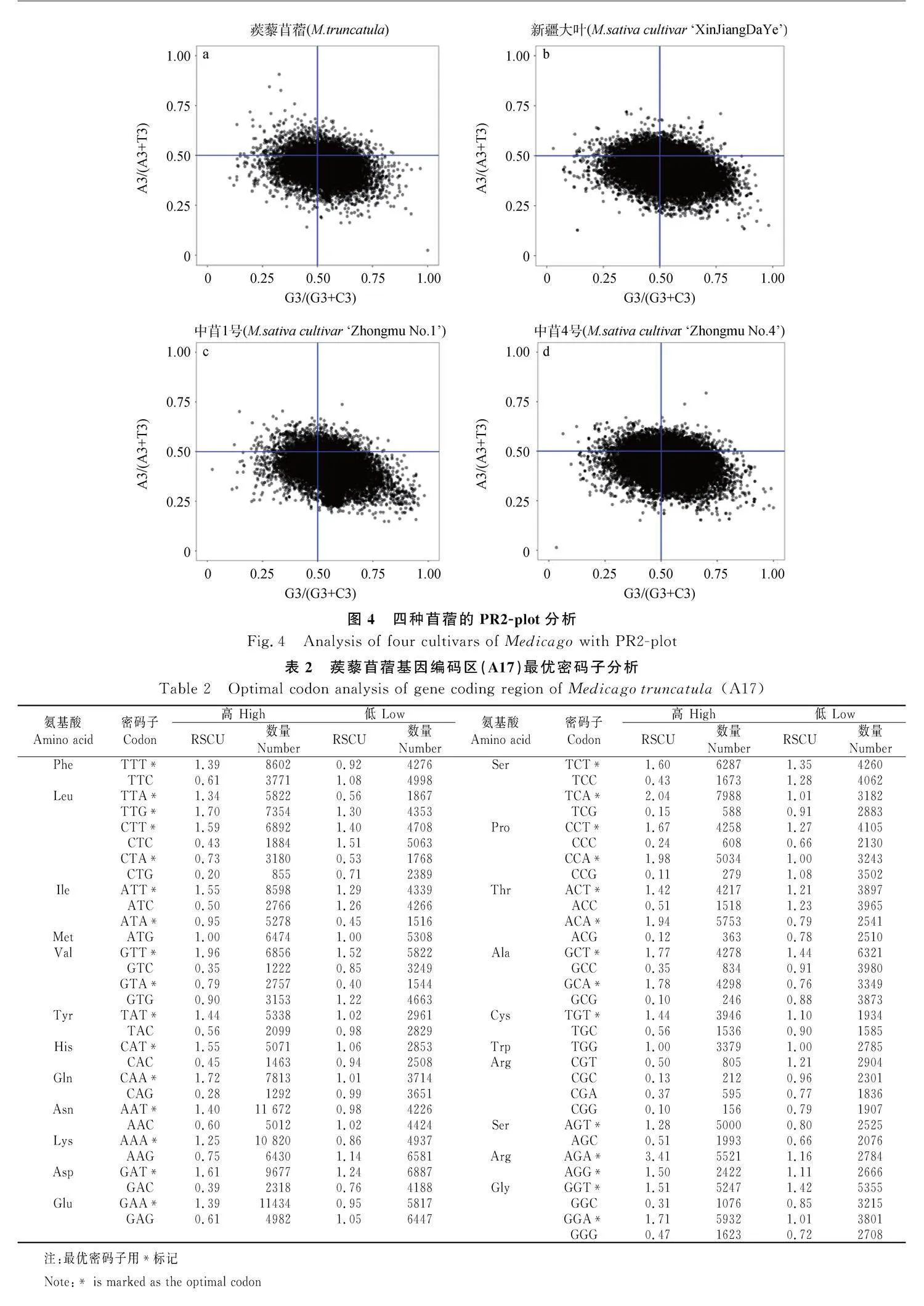
PR2-plot用于分析密碼子第三位嘧啶和嘌呤的比例關系,如果DNA兩條互補鏈之間不存在突變或選擇偏性,則理論上A3/(A3+T3)和G3/(G3+C3)的分布中心的坐標均為0.5。四種苜蓿分布中心均落在第四象限,表明堿基的使用存在突變和選擇偏性,且密碼子的第三位G的使用頻率高于C,T的使用頻率高于A,其中‘中苜1號’分布在第四象限的點所占比例最高,‘新疆大葉’和蒺藜苜蓿居中,‘中苜4號’最低(圖4)。
2.6" 最優密碼子確定
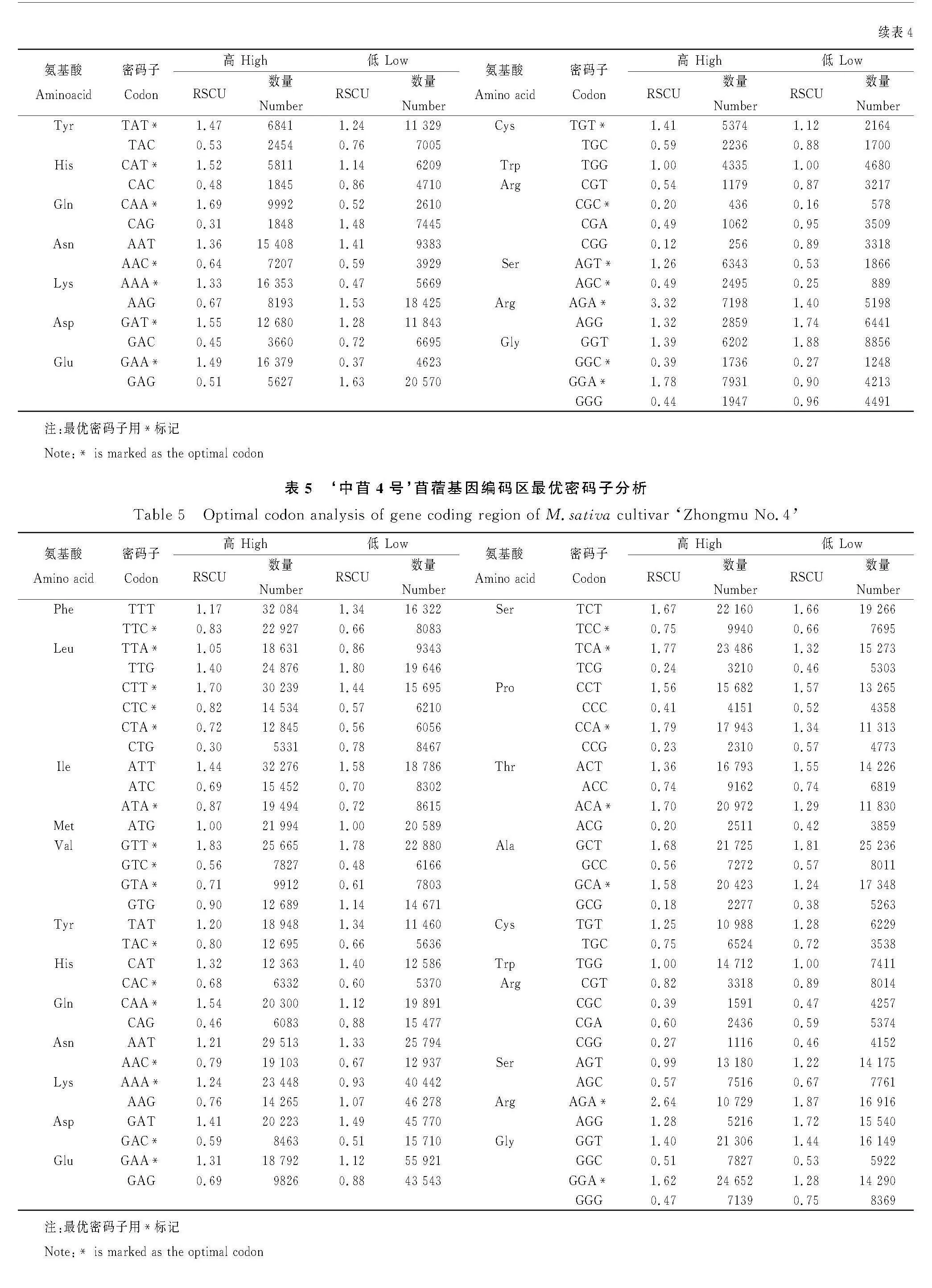
通過計算高/低表達基因之間同義密碼子相對使用頻率之差,經卡方測驗確定苜蓿基因編碼區中的最優密碼子,用*標記,結果顯示:蒺藜苜蓿最優密碼子30個(表2)、‘新疆大葉’最優密碼子28個(表3)、‘中苜1號’最優密碼子27個(表4)、‘中苜4號’最優密碼子23個(表5),四種苜蓿都表現出偏好以A/T作為密碼子的結尾,這與同義密碼子相對使用頻率的分析結果一致。
2.7" 物種間密碼子偏好性的聚類分析
利用密碼子使用頻率得出的聚類結果可以反映物種間的進化關系。聚類結果顯示:蒺藜苜蓿、‘新疆大葉’‘中苜1號’‘中苜4號’四種苜蓿的密碼子使用偏好接近;與雙子葉植物煙草、番茄偏好最為接近,其次是擬南芥、大豆。單子葉小麥和水稻的密碼子使用偏好性接近(圖5)。因此,苜蓿屬密碼子偏好性在進化上保守,而與單子葉植物有較大差異。
3" 討論
本研究分析密碼子使用偏好性的各參數中,四種苜蓿基因編碼區的GC3s含量分別為38.66%,38.50%,37.99%,38.79%,這表明四種苜蓿偏好第三位以A/T結尾的密碼子,該結果與前人對雙子葉植物的研究結果一致[30],即雙子葉植物基因的密碼子大多以A/T結尾。四種苜蓿基因編碼區的ENc均大于50(取值范圍為21~61),除了說明密碼子使用偏好性較弱以外,也表示在翻譯過程中使用同義密碼子中的任一密碼子對翻譯效率及蛋白質功能的影響都比較小。在喻鳳等[31]對紫花苜蓿葉綠體基因組密碼子偏好性的研究中,其ENc取值均高于40.6,普遍偏好性較弱,這與本文研究結果一致。針對RSCU值的分析表明,紫花苜蓿RSCU值大于1的主要是以A/T結尾的密碼子,這個研究結果和苜蓿屬豆科模式植物蒺藜苜蓿葉綠體基因組的研究結果一致[27],表明紫花苜蓿和蒺藜苜蓿的密碼子偏好性在進化上高度保守。本研究還發現四種苜蓿同義密碼子偏好性主要受環境影響。對蒺藜苜蓿葉綠體的研究發現密碼子偏好性主要受到突變的影響,也受到了其他因素的影響[27],這表明特定基因密碼子的偏好性與基因組整體偏好性并不完全匹配,這可能與基因的長度[32],翻譯準確性選擇及表達水平等有關系[33]。而且,葉綠體擁有獨立遺傳系統,其基因組具有單親遺傳性,分子進化速率低可能也有影響[34]。PR2-plot顯示四種苜蓿不均勻分布在以0.5為中心的四個區域中,說明密碼子的偏好性受突變和自然選擇的共同影響,這與前人密碼子偏好性非單一影響因素結果一致[35]。并且,聚類分析結果顯示四種苜蓿之間的密碼子偏好性相似,可能和近緣種間基因組保守性有關。對苜蓿基因編碼區最優密碼子的分析結果與同義密碼子使用頻率分析結果一致,得出不同品種苜蓿的最優密碼子可以作為不同苜蓿密碼子優化的重要組成部分。最后,通過與煙草、大豆等的密碼子使用頻率的聚類分析,確定煙草、番茄均適合作為苜蓿基因的外源表達系統,煙草易于遺傳轉化,這為苜蓿基因的密碼子優化和開展基因工程育種奠定了基礎。但是,值得注意的是,大豆和苜蓿都為豆科植物,在密碼子偏好性的接近程度卻不如番茄和苜蓿,這可能是因為大豆雖然是豆科植物,但二倍體與苜蓿四倍體的倍型差異大導致;同時,水稻、玉米和小麥密碼子偏好性更為接近,由此可知,單雙子葉植物內部在進化中各自具有相似的密碼子使用模式,之前已有學者發現單子葉偏好G或C結尾的密碼子,雙子葉植物偏好A或T結尾的密碼子,這與長期進化中兩者基因組GC含量不同吻合[36]。此外,苜蓿與酵母菌的偏好性也有一定的相似性,可以為酵母的基因在植物表達載體中的功能檢測提供依據[37],而且酵母菌、大腸埃希菌等低等生物更需要優化密碼子,這可能是因為高等生物的基因表達還受到順式作用元件、轉錄后調控、選擇性剪切等多種因素調節[28],同時也暗示了高等生物比低等生物更能適應復雜變化的環境。
轉基因研究經常涉及基因的異源表達,異源物種的選擇是決定基因表達效率的重要因素。因此,研究密碼子偏好性并對其進行優化,選擇與目的基因密碼子使用偏好性相似的異源受體能大大提高基因表達效率。而且,通過比較某一特定基因與外源表達系統之間的密碼子使用偏好性差異來判斷是否會引起甲基化,避免出現基因表達量下降或基因沉默情況,從而實現外源基因在宿主中的表達[38-39]。本文研究結果為苜蓿密碼子的優化提供了理論基礎,提高了Cas9蛋白的表達量從而實現高效率的基因編輯,從而輔助分子設計育種。
4" 結論
本研究選取了四種苜蓿的基因組編碼區序列分析密碼子組成和同義密碼子使用頻率發現,四種苜蓿密碼子使用偏好性較弱,最弱的是‘中苜1號’,且苜蓿屬偏好以A/T結尾的密碼子。多種分析方法顯示四種苜蓿基因編碼區的密碼子使用的偏好性主要受自然選擇影響,最優密碼子偏好以A/T結尾。苜蓿屬密碼子偏好性相似,與煙草、番茄最為接近,與單子葉植物有顯著差異。本研究為苜蓿密碼子優化提供了依據,為提高Cas9表達量和基因編輯效率提供了參考。
參考文獻
[1]" 楊云彭,馬曉焉,霍毅欣. 密碼子優化策略在異源蛋白表達中的應用[J]. 生物工程學報,2019,35(12):2227-2237
[2]" PARVATHY S T,UDAYSURIYAN V,BHADANA V. Codon usage bias[J]. Molecular Biology Reports,2021,49(1):539-565
[3]" 洪園淑,周玉梅,李文娟,等. 苦豆子SaLDC密碼子偏好性分析、優化及原核表達[J]. 中成藥,2021,43(2):429-435
[4]" 陳悅,仇學文,李鈺,等. 辣椒和番茄PIF基因家族密碼子偏好性分析[J]. 分子植物育種,2023,21(17):5602-5610
[5]" 陳哲,胡福初,王祥和,等. 菠蘿轉錄組基因密碼子使用偏好性分析[C]//中國熱帶作物學會,中國熱帶作物學會2016年學術年會論文集.南寧:2016:2
[6]" LI L,HU Y F,HE M,et al. Comparative chloroplast genomes:insights into the evolution of the chloroplast genome of Camellia sinensis and the phylogeny of Camellia[J].BMC Genomics,2021,22(1):101186
[7]" 冉然,張曉明,宛濤,等. 密碼子偏好性的相關研究及方法分析[J]. 草原與草業,2022,34(3):5-10,43
[8]" 任桂萍,董瓔瑩,黨云琨. 密碼子中的密碼:密碼子偏好性與基因表達的精細調控[J]. 中國科學:生命科學,2019,49(7):839-847
[9]" QUAX T E F,CLAASSENS N J,SOELL D,et al. Codon bias as a means to fine-tune gene expression[J]. Molecular Cell,2015,59(2):149-161
[10]JIA J,QING Z X. Codon usage biases of trantransposable elements and host nuclear genes in Arabidopsis thaliana and Oryza sativa[J]. Genomics Proteomics Bioinformatics,2009,7(4):175-184
[11]KOKATE P P,TECHMAN S M,WERNER T. Codon usage bias and dinucleotide preference in 29 Drosophila species[J]. G3-Genes Genomes Genetics,2021,11(8):101093
[12]LIU X Y,LI Y,JI K K,et al. Genome-wide codon usage pattern analysis reveals the correlation between codon usage bias and gene expression in Cuscuta australis[J]. Genomics,2020,112(4):2695-2702
[13]陳徵婷,可小麗,陳剛,等. 基于密碼子優化策略的無乳鏈球菌表面蛋白LrrG的原核表達、純化及免疫原性[J]. 大連海洋大學學報,2021,36(6):920-928
[14]陳凱.CRISPR技術的優化及Cas9酶PAM識別位點的改造[D].北京:中國農業科學院,2018:36-37
[15]時歡,林玉玲,賴鐘雄,等. CRISPR/Cas9介導的植物基因編輯技術研究進展[J]. 應用與環境生物學報,2018,24(3):640-650
[16]吳少平.香蕉CRISPR/Cas9系統的優化及香蕉種質的創制[D].長沙:湖南農業大學,2023:31-32
[17]SHI T Q,GAO J,WANG W J,et al. CRISPR/Cas9-based genome editing in the filamentous fungus Fusarium fujikuroi and its application in strain engineering for gibberellic acid production[J]. ACS Synthetic Biology,2019,8(2):445-454
[18]楊青川,孫彥. 中國苜蓿育種的歷史、現狀與發展趨勢[J]. 中國草地學報,2011,33(6):95-101
[19]毛培勝,王明亞,歐成明. 中國草種業的發展現狀與趨勢分析[J]. 草學,2018(6):1-6
[20]陳筱冉,張銘笑,嚴建萍,等. 紫花苜蓿MsCCD4基因克隆及耐鹽功能鑒定[J]. 草地學報,2022,30(10):2572-2580
[21]BIN PEK H,KLEMENT M,ANG K S,et al. Exploring codon context bias for synthetic gene design of a thermostable invertase in Escherichia coli[J]. Enzyme and Microbial Technology,2015(75-76):57-63
[22]PAN L L,WANG Y,HU J H,et al. Analysis of codon use features of stearoyl-acyl carrier protein desaturase gene in Camellia sinensis[J]. Journal of Theoretical Biology,2013,334:80-86
[23]吉德娟,王占林. 霸王密碼子偏好性分析[J]. 分子植物育種,2023,21(20):6705-6713
[24]郝新艷,劉嘉偉,米福貴. 雜花苜蓿葉綠體基因組密碼子使用偏好性分析[J/OL]. http://kns.cnki.net/kcms/detail/46.1068.S.20230524.1400.012.html.2023-05-25/2024-02-21
[25]李蓉,謝析穎,王雪晶,等. 蘭科植物FNR基因的密碼子偏好性分析[J]. 熱帶作物學報,2018,39(6):1137-1145
[26]丁銳,胡兵,宗小雁,等. 杓蘭葉綠體基因組密碼子偏好性分析[J]. 林業科學研究,2021,34(5):177-185
[27]楊國鋒,蘇昆龍,趙怡然,等. 蒺藜苜蓿葉綠體密碼子偏好性分析[J]. 草業學報,2015,24(12):171-179
[28]劉潮,韓利紅,吳麗芳,等. 辣椒基因組密碼子使用偏性分析[J]. 江蘇農業科學,2022,50(5):16-22
[29]宗秋芳,黃焱杰,吳麗思,等. 豬Claudin家族基因密碼子使用偏好性分析[J]. 浙江農業學報,2018,30(12):2007-2017
[30]KAWABE A,MIYASHITA N T. Patterns of codon usage bias in three dicot and four monocot plant species[J]. Genes amp; Genetic Systems,2003,78(5):343-352
[31]喻鳳,韓明. 紫花苜蓿葉綠體基因組密碼子偏好性分析[J]. 廣西植物,2021,41(12):2069-2076
[32]SUN Z,MA L,MURPHY R,et al. Analysis of codon usage on Wolbachia pipientis wMel genome[J]. Science in China Press:Life Sciences,2009,39(10):948-953
[33]HIRAOKA Y,KAWANMATA K,HARAGUCHI T,et al. Codon usage bias is correlated with gene expression levels in the fission yeast Schizosaccharomyces pombe[J]. Genes to Cells,2009,14(4):499-509
[34]劉文雅,王久利,何彥峰. 紫丁香葉綠體基因組密碼子偏好性分析[J/OL]. http://kns.cnki.net/kcms/detail/46.1068.S.20230908.1300.004.html.2023-09-11/2024-02-11
[35]代江鵬,蔡一鳴,劉巧珍,等. 甘草屬7種植物葉綠體基因組密碼子使用偏好性分析[J]. 中草藥,2023,54(9):2907-2916
[36]劉漢梅,何瑞,張懷渝,等. 玉米同義密碼子偏愛性分析[J]. 農業生物技術學報,2010,18(3):456-461
[37]蔣瑞平,趙辰暉,李文杰,等. 豆科植物IPI基因密碼子偏好性[J]. 浙江農業學報,2022,34(6):1114-1123
[38]SHAO Z Q,ZHANG Y M,FENG X Y,et al. Synonymous codon ordering:a subtle but prevalent strategy of bacteria to improve translational efficiency[J]. Plos One,2012,7(3):101371
[39]QIAN W F,YANG J R,PEARSON N M,et al. Balanced codon usage optimizes eukaryotic translational efficiency[J]. Plos Genetics,2012,8(3):101371
(責任編輯" 彭露茜)
