油莎豆CePIP2;1的克隆、亞細胞定位與表達分析
2024-12-31 00:00:00鄒智鄭玉皎常麗麗趙永國
熱帶作物學報 2024年7期
關鍵詞:分析

摘""要:油莎豆是一種在塊莖中高水平積累油脂的草本油料作物,其未成熟塊莖的含水量高達85%,水分平衡對于塊莖的發育與代謝至關重要。質膜內在蛋白(PIP)是介導細胞間水分跨膜運輸的主要通道。本研究報道1個塊莖高水平表達的PIP基因CePIP2;1,該基因含有3個內含子,預測編碼288個氨基酸(aa),其理論分子量(MW)為30.34"kDa,等電點(pI)為8.60,總平均疏水指數(GRAVY)為0.529,不穩定系數(Ⅱ)為29.60,屬于典型的穩定、堿性、疏水型蛋白。CePIP2;1含有保守的MIP結構域,ar/R選擇性濾器為F-H-T-R,Froger位點為Q-S-A-F-W,符合高水分轉運活性PIP的特征。進化分析顯示,CePIP2;1與OsPIP2;1、OsPIP2;2和OsPIP2;3聚在一起,序列相似性分別為91.72%、90.31%和84.98%,支持其歸為PIP2亞類。亞細胞定位分析顯示,CePIP2;1定位在煙草葉片的細胞膜。進一步的表達分析顯示,雖然CePIP2;1為主要的塊莖表達PIP基因,但其在葉片、葉鞘、匍匐莖和根等組織中的表達豐度更高,最高的為葉片,而在芽尖中的表達豐度與塊莖相當;在塊莖的4個典型發育時期(S1~S4)中,CePIP2;1基因呈現先升后降的鐘形趨勢,表達豐度最高的是S2。這些結果為解析油莎豆的水分平衡機制奠定基礎。
關鍵詞:油料作物;塊莖;水通道蛋白;質膜內在蛋白;亞細胞定位;表達模式中圖分類號:S565.9""""""文獻標志碼:A
Cloning,"Subcellular"Localization,"and"Expression"Analysis"of"CePIP2;1,"an"Aquaporin"Gene"from"Tigernut"(Cyperus"esculentus"L.)
ZOU"Zhi,"ZHENG"Yujiao,"CHANG"Lili,"ZHAO"Yongguo
National"Key"Laboratory"for"Tropical"Crop"Breeding"/"Institute"of"Tropical"Biosciences"and"Biotechnology,"Chinese"Academy"of"Tropical"Agricultural"Sciences"/"Sanya"Research"Institute"of"Chinese"Academy"of"Tropical"Agricultural"Sciences,"Haikou,"Hainan"571101,"China
Abstract:"Tigernut"(Cyperus"esculentus"L.)"is"an"herbaceous"oil"crop"uniquely"accumulating"a"high"level"of"oil"in"the"underground"tubers."Water,"which"accounts"for"approximately"85%"of"immature"tubers,"is"essential"for"tuber"development"and"metabolism."Plasma"membrane"intrinsic"proteins"(PIPs)"represent"the"main"channel"that"mediates"the"efficient"water"transport"across"the"cell"membrane."This"study"presents"the"characterization"of"CePIP2;1,"a"PIP"gene"highly"abundant"in"tigernut"tubers."This"gene"with"three"introns"was"shown"to"encode"288"amino"acids"with"the"theoretical"molecular"weight"of"30.34"kDa,"the"isoelectric"point"of"8.60,"the"grand"average"of"hydropathicity"of"0.529,"and"the"instability"index"of"29.60,"implying"it’s"stable,"basic"and"hydrophobic"characteristics."The"protein"was"shown"to"possess"one"conservative"MIP"(major"intrinsic"protein)"domain,"and"presence"of"the"F-H-T-R"ar/R"selectivity"filter"and"Q-S-A-F-W"Froger’s"positions"implies"a"putatively"efficient"water"transport"activity."Phylogenetic"analysis"revealed"that"CePIP2;1"clusters"with"OsPIP2;1,"OsPIP2;2,"and"OsPIP2;3"with"the"similarities"of"91.72%,"90.31%"and"84.98%,"respectively,"supporting"its"assignment"into"the"PIP2"group"of"the"PIP"subfamily."Subcellular"localization"analysis"showed"that"CePIP2;1"was"localized"to"the"cell"membrane"of"Nicotiana"benthamiana"leaves."Although"CePIP2;1"was"characterized"as"a"dominant"isoform"in"tubers,"expression"analysis"indicated"that"its"transcripts"were"relatively"more"in"leaves,"leaf"sheaths,"rhizomes,"and"roots"with"the"most"in"leaves,"in"contrast"to"a"comparable"level"to"that"in"shoot"apexes."Among"four"typical"stages"(S1–S4)"of"tuber"development"examined"in"this"study,"CePIP2;1"was"shown"to"exhibit"a"bell-shaped"expression"pattern,"peaking"at"S2."These"results"laid"a"solid"foundation"for"further"uncovering"the"mechanism"of"water"balance"in"tigernut.
Keywords:"oil"crop;"tuber;"aquaporin;"plasma"membrane"intrinsic"protein;"subcellular"localization;"expression"profile
DOI:"10.3969/j.issn.1000-2561.2024.07.005
質膜內在蛋白(plasma"membrane"intrinsic"protein,"PIP)隸屬于水通道蛋白(aquaporin,"AQP)家族,其以定位在細胞膜且具有高效的水分轉運活性而著稱[1]。除水分外,PIP還可以轉運尿素、硼酸、CO2、H2O2、O2和Na+[2]。在高等植物中,PIP亞族的成員數介于2~28個不等,可以分為PIP1和PIP2兩個進化小組[3-9]。目前晶體結構已解析的有菠菜(Spinacia"oleracea)PIP2;1、擬南芥(Arabidopsis"thaliana)PIP2;1(PIP2或PIP2A)和AtPIP2;4(PIP2F)[10-12]。研究表明,PIP在細胞膜上以四聚體的形式起作用,每個單體包含6個跨膜螺旋(TM1~TM6)和5個連環(LA、LB、LC、LD和LE),其中,LB、LD、N端和C端位于胞質,而LA、LC和LE位于胞外;LB和LE折向膜內形成2個半螺旋,半螺旋的頂端含有高度保守的NPA基序,起排斥質子并決定通道大小的作用;位于TM2、TM5和LE上的4個保守殘基(H2、H5、LE1和LE2)決定了底物特異性,因H2和LE2分別為芳香氨基酸和精氨酸而被稱為ar/R選擇性濾器[10]。此外,FROGER等[13]從甘油通道蛋白(glyceroporin,"GLP)中鑒定到區別于AQP的5個殘基,即P1為芳香氨基酸、P2為酸性氨基酸、P3為堿性氨基酸、P4為脯氨酸以及P5為非芳香氨基酸。除轉錄外,PIP還受翻譯后修飾(如磷酸化、甲基化、乙酰化、泛素化、糖基化、脫酰胺基)、二聚化、轉運、門控(如二價陽離子、pH、磷酸化)和蛋白互作等調控[10-11,"14]。位于SoPIP2;1"LA的C69為二聚體的形成所必須;位于LD的H193和L197為門控位點;位于LB的S115、LD的S188以及位于C端的S274為門控調控磷酸化位點[10]。
油莎豆(Cyperus"esculentus"L.),俗稱黃色道格拉斯(yellow"nutsedge)、虎堅果(tigernut),是一種隸屬于禾本目莎草科的新型草本油料作物[15-19]。不同于傳統油料作物在種子中積累油脂,油莎豆是迄今唯一已知在塊莖中高水平積累油脂的作物,這使其成為研究營養組織油脂代謝與調控的模式[20-23]。塊莖起源于地下匍匐莖,其發育過程可分為起始、膨大和成熟等3個主要階段;前2個階段的含水量高達85%,而在成熟期發生類似種子的脫水過程,含水量逐漸降低到45%以下[17,"20]。水分平衡對于塊莖的發育乃至油脂和淀粉等儲藏物的積累至關重要[20],因而挖掘塊莖中的關鍵PIP基因具有重要的理論意義和應用價值。本研究對1個塊莖高水平表達的PIP基因進行了克隆和初步鑒定,以期為揭示塊莖的水分平衡機制奠定基礎。
1""材料與方法
1.1""材料
研究所用油莎豆品系為熱研3號,植株種植于中國熱帶農業科學院文昌實驗基地[21],待塊莖起始后分別采集1"d(起始期,S1)、20"d(膨大中期,S2)、25"d(膨大晚期,S3)和35"d(成熟期,S4)的樣本,每樣本3次生物學重復。本氏煙草(Nicotiana"benthamiana)用小缽種在中國熱帶農業科學院熱帶生物技術研究所溫室[24]。大腸桿菌(Escherichia"coli)DH5α和含pSoup-P19的農桿菌(Agrobacterium"tumefaciens)GV3101感受態均由本實驗室制備和保存;亞細胞定位載體pNC-HbPIP2;3-RFP[25]和pNC-Cam1304-SubN由本實驗室保存;各類酶、試劑盒和生化試劑購自相應的試劑公司。本研究所用數據庫、軟件及其網址詳見表1。
1.2""方法
1.2.1""總RNA的提取與反轉錄""采用天根植物多糖多酚RNA提取試劑盒提取上述塊莖的總RNA,經濃度和完整性檢測合格后用賽默飛反轉錄試劑盒合成cDNA第一鏈,用于后續的基因克隆與表達分析。
1.2.2""基因克隆與載體構建""前期通過分析油莎豆的轉錄組數據[21],發現1個在塊莖中高水平表達的PIP基因,根據序列特征和進化關系將其命名為CePIP2;1。為克隆該基因,采用Primer"Premier"5.0設計如表2所示的引物對,首先用CePIP2;1F/R擴增其1127"bp的全長cDNA,接著以稀釋100倍的首輪PCR產物作為模板、CePIP2;1HF/R為引物擴增907"bp的全長編碼區(CDS)。反應體系:模板0.5"μL、引物各0.5"μL、PrimeSTAR"Max"DNA"Polymerase"12.5"μL,加ddH2O至25"μL。PCR擴增程序:95"℃預變性1"min;94"℃變性30"s、50"℃退火2"min,35個循環;72"℃延伸5"min。亞細胞定位載體pNC-Cam1304-CePIP2;1的構建參照NC克隆試劑盒說明書。
1.2.3""基因和蛋白結構分析""采用BLASTn將基因的轉錄本序列比對到基因組,然后用GSDS"2.0軟件展示基因結構。分別采用Protparam、CDD、SOPMA、SWISS-MODEL、WoLF"PSORT等軟件分析CePIP2;1編碼蛋白的理化特性、保守結構域、二級結構、3D結構和亞細胞定位。跨膜螺旋和半螺旋區及保守氨基酸基于與SoPIP2;1、AtPIP2;1和AtPIP2;4的序列比對進行確定。
1.2.4""多序列比對和進化分析""多序列比對采用MUSCLE軟件,選用默認參數;進化樹的構建采用MEGA"6.06軟件,選用最大似然法,1000次重復,其他用默認參數。
1.2.5""亞細胞定位分析""以先前報道的pNC-"HbPIP2;3-RFP[25]為陽性對照,參照文獻[24]進行農桿菌的轉化、培養及浸染液的制備。將含pNC-Cam1304-CePIP2;1的實驗組與陽性對照菌液等量混合后采用微量注射器注射4周齡的煙草葉片,轉化48"h后用激光共聚焦顯微鏡Zeiss"LMS880觀察熒光在細胞內的位置。
1.2.6""基因表達分析""為分析基因的組織特異性,參照文獻[23],采用StringTie(v2.2.0)軟件將不同組織的轉錄組數據(PRJNA703731)比對到基因組,基因的相對表達豐度用FKPM(fragments"per"kilobase"of"exon"per"million"fragments"mapped)法進行均一化。熒光定量分析以先前報道的CeUCE2和CeTIP41[18]作為內參基因,利用引物對CePIP2;1Fq/Rq進行qRT-PCR分析,數據分析采用2-△△CT法,統計分析采用SPSS"20軟件,采用鄧肯法進行單因素多樣本間差異顯著性方差分析。
2""結果與分析
2.1""CePIP2;1基因的克隆與序列分析
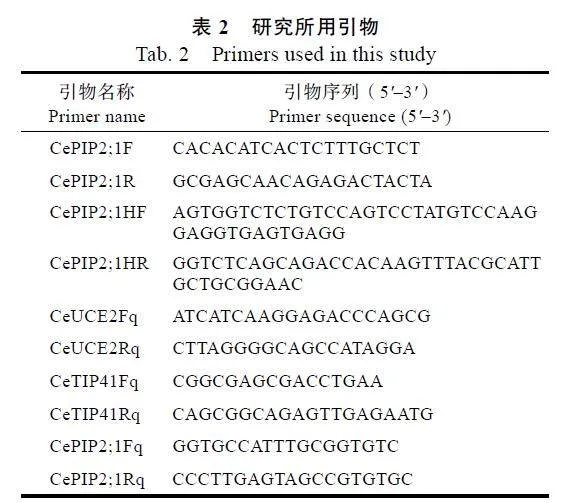
經過2輪巢式PCR擴增,成功分離到CePIP2;1的CDS序列,并采用無縫克隆技術將其克隆到亞細胞定位載體,構建重組質粒pNC-Cam1304-CePIP2;1。通過將克隆到的序列比對到基因組,發現其與CePIP2;1基因的對應區域完成一致;CePIP2;1位于Scaffold9正向鏈的879960~884243位,基于轉錄組的轉錄區界定顯示基因全長為5752"bp,包含3個內含子,其長度分別為806、1745、866"bp;5'和3'"UTR分別為119、1349"bp(圖1A)。
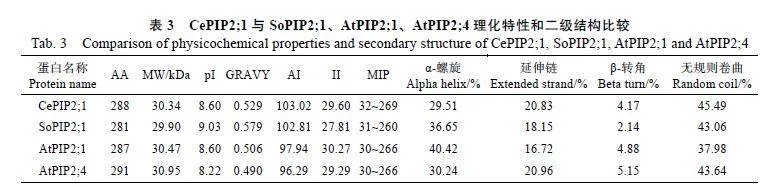
序列分析顯示,CePIP2;1編碼288個氨基酸(表3),其中,A和G的含量最高,分別占12.50%和12.20%,這與SoPIP2;1、AtPIP2;1和AtPIP2;4類似(圖1B);CePIP2;1的理論分子量(MW)為30.34"kDa、等電點(pI)為8.60、脂肪族指數(AI)為103.02、總平均疏水指數(GRAVY)為0.529、不穩定系數(Ⅱ)為29.60,表明其為穩定的堿性疏水型蛋白;CDD分析顯示蛋白的32~269位為MIP結構域(圖1C),略長于SoPIP2;1、AtPIP2;1和AtPIP2;4(表3);CePIP2;1與SoPIP2;1、AtPIP2;1、AtPIP2;4的序列相似性分別為84.88%、84.14%、84.35%,其結構包括TM1~TM6等6個典型的跨膜螺旋、HB和HE"2個半螺旋、2個典型的NPA基序、多個磷酸化位點(S105、S124、S283和S286);ar/R選擇性濾器(F90-H219-T228-R234)和對應于SoPIP2;1"C69的二聚化位點、S115的磷酸化位點及H193和L197門控位點在所比較的4個蛋白中完全一致;在5個Froger位點中,僅P1存在差異,SoPIP2;1為M141-S226-A230-F245-W246,而CePIP2;1為Q150-
S235-A239-F254-W255(圖1D)。
為揭示CePIP2;1的進化關系,將其與SoPIP2;1及水稻、擬南芥中的所有PIP構建如圖1E所示的進化樹。多數成員按物種聚在一起,暗示相關成員在物種分化之后產生。CePIP2;1與OsPIP2;1、OsPIP2;2、OsPIP2;3聚在一起,其序列相似性分別為91.72%、90.31%和84.98%。二級結構預測顯示,CePIP2;1以無規則卷曲(45.49%)和α-螺旋為主(29.51%),其次是延伸鏈(20.83%),最低的為β-轉角(4.17%),與SoPIP2;1、AtPIP2;1和AtPIP2;4類似(表3)。基于SoPIP2;1的同源建模顯示,CePIP2;1可以形成同源四聚體(圖1F)。
2.2""亞細胞定位分析
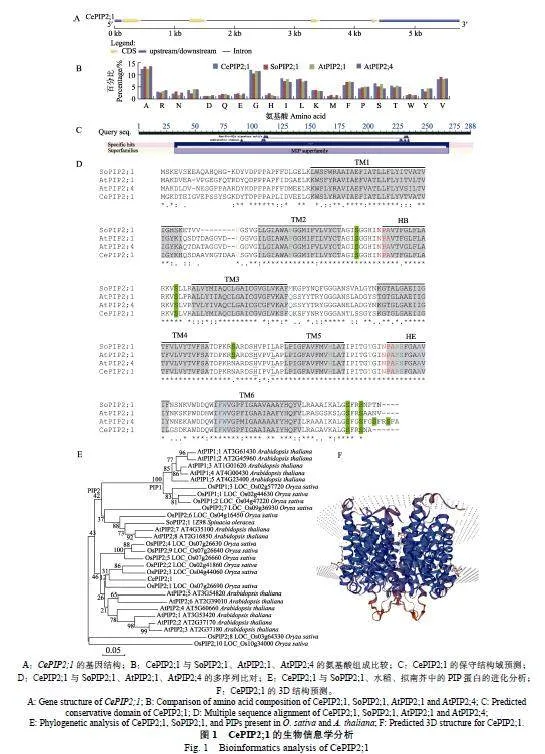
與其他PIP蛋白類似[24-25],WoLF"PSORT在線軟件預測顯示CePIP2;1定位在細胞膜。為驗證該結果,本研究以先前報道的HbPIP2;3[25]作為陽性對照,將含pNC-HbPIP2;3-RFP和pNC-Cam1304-"CePIP2;1的農桿菌工程菌等量混合后微量注射煙草葉片。共聚焦觀察結果顯示,GFP-CePIP2;1的綠色熒光信號出現在細胞膜,并與HbPIP2;3-RFP的紅色熒光信號高度重合,這表明CePIP2;1確實是定位在細胞膜(圖2)。
2.3""基因表達分析
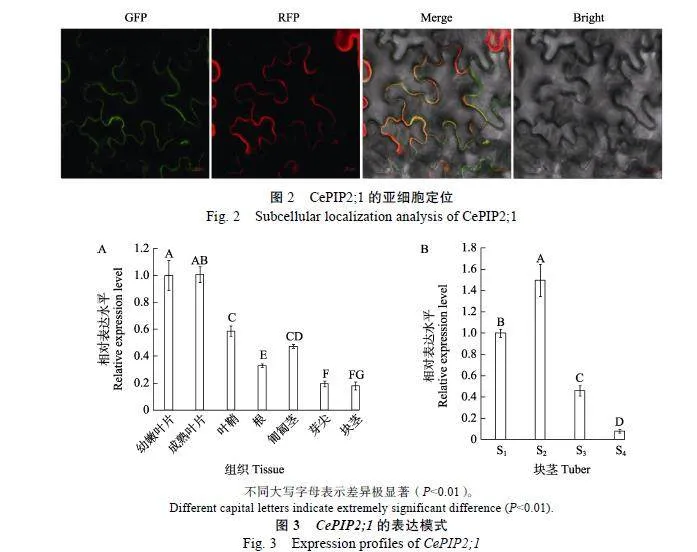
為揭示CePIP2;1基因表達的組織特異性,利用轉錄組數據分析其在幼嫩葉片、成熟葉片、葉鞘、根、匍匐莖、芽尖和塊莖等主要組織中的表達模式。結果顯示,CePIP2;1在這些組織中均高水平表達,豐度最高的是葉片(幼嫩和成熟葉片差異不顯著),最低的為芽尖,其與塊莖豐度相當(圖3A)。
為揭示基因表達與不同發育時期塊莖含水量之間的相關性,挑選自塊莖形成起始期(S1)、膨大中期(S2)、膨大晚期(S3)和成熟期(S4)等4個典型時期,其含水量分別為83.7%、86.0%、74.8%和48.3%[17]。qRT-PCR分析結果如圖3B所示,CePIP2;1基因呈先升后降的趨勢,峰值出現在S2,約為S1的1.5倍;豐度最低的是S4,僅為S1的8%;S3僅為S2的30%。
3""討論
水是生命的主要存在形式,廣泛參與體內的各項生理生化反應,為植物的生長發育和代謝活動所必需[2]。研究表明,植物細胞間水分的跨膜運輸主要由細胞膜定位的PIP介導[1,"2]。進化分析顯示,PIP高度保守,僅包含PIP1和PIP2兩個亞類;雖然他們早在苔蘚中就已分化形成,但二者之間的序列相似性一般在70%以上[3-9,"26]。PIP通常具有高效的水分轉運活性[2]。雖然有研究表明PIP1亞類在酵母和蟾蜍卵母細胞等體外體系中無活性或活性很低,主要歸因于其自身不能有效定位到細胞膜[27-28]。在植物體內,PIP2的存在可以介導PIP1的有效定位[27-29]。
在前期的研究中,本團隊系統分析了塊莖的發育特征及發育過程中的含水動力學,發現在我國海南地區塊莖的整個發育期約為35"d,其中,前、中期伴隨著細胞的快速分裂和擴張,水分含量一直維持在85%,而后期隨著塊莖的成熟,水分含量逐步降低,25"d和35"d分別為74.8%和48.3%[17],發生類似種子的脫水過程[17,"30]。雖然如此,至今對油莎豆塊莖發育的水分平衡機制知之甚少。
本研究首次報道了1個油莎豆PIP2基因CePIP2;1。與大多數PIP基因類似[3-9],CePIP2;1含有3個內含子;其編碼蛋白包含AQP家族特有的MIP結構域,分子量為30.76"kDa,與先前的報道相似[5-9,"25-26]。CePIP2;1具有較高的脂肪族指數(95.28),且總平均疏水指數gt;0、等電點gt;7.0、不穩定系數為lt;40,屬于典型的穩定、堿性、疏水型蛋白。在水稻的13個PIP蛋白中,CePIP2;1與OsPIP2;1的序列相似性最高(91.72%),略高于與OsPIP2;2的90.31%,但遠高于與OsPIP1的72.19%~74.59%,這與本研究的結果一致,支持其劃歸為PIP2亞類。此外,CePIP2;1具有SoPIP2;1[10]、AtPIP2;1[11]和AtPIP2;4[12]相似的結構特征,其中包括典型的NPA基序、F-H-T-R"ar/R選擇性濾器、Q-S-A-F-W"Froger位點、位于LA高度保守的C殘基、可形成四聚體,并且被實驗證實定位在煙草葉片的細胞膜,這些均表明CePIP2;1可能具有高效的水分轉運活性。雖然CePIP2;1首先是作為1個塊莖高豐度PIP基因而被關注,但組織特異性分析顯示它在其他組織中的表達豐度更高,暗示其功能的重要性。在塊莖的發育過程中,CePIP2;1呈現先升后降的表達模式,即隨著細胞的快速分裂和擴張,CePIP2;1的表達迅速上調,至膨大中期達到峰值,而在成熟過程中逐漸下調,這與相應時期的水分含量趨勢基本一致[17]。雖然如此,CePIP2;1具體是如何調控塊莖及其他組織的水分平衡還有待深入研究,比如門控調控、翻譯后修飾、蛋白互作模式以及調控其轉錄的上游轉錄因子。
綜上,本研究完成了塊莖高水平表達CePIP2;1基因的克隆與初步鑒定,明確了其基因結構、序列特征、進化關系、亞細胞定位和表達特性,這為進一步的功能分析及油莎豆遺傳改良奠定了堅實的基礎。
參考文獻
[1]"DANIELS"M"J,"MIRKOV"T"E,"CHRISPEELS"M"J."The"plasma"membrane"of"Arabidopsis"thaliana"contains"a"mercury-insensitive"aquaporin"that"is"a"homolog"of"the"tonoplast"water"channel"protein"TIP[J]."Plant"Physiology,"1994,"106(4):"1325-1333.
[2]"TYERMAN"S"D,"MCGAUGHEY"S"A,"QIU"J,"YOOL"A"J,"BYRT"C"S."Adaptable"and"multifunctional"ion-conducting"aquaporins[J]."Annual"Review"Plant"Biology,"2021,"72:"703-736.
[3]"JOHANSON"U,"KARLSSON"M,"JOHANSSON"I,"GUSTAVSSON"S,"SJ?VALL"S,"FRAYSSE"L,"WEIG"A"R,"KJELLBOM"P."The"complete"set"of"genes"encoding"major"intrinsic"proteins"in"Arabidopsis"provides"a"framework"for"a"new"nomenclature"for"major"intrinsic"proteins"in"plants[J]."Plant"Physiology,"2001,"126(4):"1358-1369.
[4]"SAKURAI"J,"ISHIKAWA"F,"YAMAGUCHI"T,"UEMURA"M,"MAESHIMA"M."Identification"of"33"rice"aquaporin"genes"and"analysis"of"their"expression"and"function[J]."Plant"and"Cell"Physiology,"2005,"46(9):"1568-1577.
[5]"ZOU"Z,"GONG"J,"AN"F,"XIE"G,"WANG"J,"MO"Y,"YANG"L."Genome-wide"identification"of"rubber"tree"(Hevea"brasiliensis"Muell."Arg.)"aquaporin"genes"and"their"response"to"ethephon"stimulation"in"the"laticifer,"a"rubber-producing"tissue[J]."BMC"Genomics,"2015,"16(1):"1001.
[6]"ZOU"Z,"GONG"J,"HUANG"Q,"MO"Y"Y,"YANG"L"F,"XIE"G"S."Gene"structures,"evolution,"classification"and"expression"profiles"of"the"aquaporin"gene"family"in"castor"bean"(Ricinusnbsp;communis"L.)[J]."PLoS"One,"2015,"10(10):"e0141022.
- ZOU"Z,"YANG"L"F,"GONG"J,"MO"Y"Y,"WANG"J"K,"CAO"J"H,"AN"F,"XIE"G"S."Genome-wide"identification"of"Jatropha"curcas"aquaporin"genes"and"the"comparative"analysis"provides"insights"into"the"gene"family"expansion"and"evolution"in"Hevea"brasiliensis[J]."Frontiers"in"Plant"Science,"2016,"7:"395.
- ZOU"Z,"YANG"J"H."Genome-wide"comparison"reveals"divergence"of"cassava"and"rubber"aquaporin"family"genes"after"the"recent"whole-genome"duplication[J]."BMC"Genomics,"2019,"20:"380.
- ZOU"Z,"ZHENG"Y"J,"XIE"Z"N."Analysis"of"Carica"papaya"informs"lineage-specific"evolution"of"the"aquaporin"(AQP)"family"in"Brassicales[J]."Plants"(Basel),"2023,"12(22):"3847.
- TORNROTH-HORSEFIELD"S,"WANG"Y,"HEDFALK"K,"JOHANSON"U,"KARLSSON"M,"TAJKHORSHID"E,"NEUTZE"R,"KJELLBOM"P."Structural"mechanism"of"plant"aquaporin"gating[J]."Nature,"2006,"439(7077):"688-694."
- VERDOUCQ"L,"GRONDIN"A,"MAUREL"C."Structure-function"analysis"of"plant"aquaporin"AtPIP2;1"gating"by"divalent"cations"and"protons[J]."The"Biochemical"Journal,"2008,"415(3):"409-416.
- WANG"H,"SCHOEBEL"S,"SCHMITZ"F,"DONG"H,"HEDFALK"K."Characterization"of"aquaporin-driven"hydrogen"peroxide"transport[J]."Biochimica"et"Biophysica"Acta"(BBA)–Biomembranes,"2020,"1862(2):"183065.
- FROGER"A,"TALLUR"B,"THOMAS"D,"DELAMARCHE"C."Prediction"of"functional"residues"in"water"channels"and"related"proteins[J]."Protein"Science,"1998,"7(6):"1458-168.
- QIU"J,"MCGAUGHEY"S"A,"GROSZMANN"M,"TYERMAN"S"D,"BYRT"C"S."Phosphorylation"influences"water"and"ion"channel"function"of"AtPIP2;1[J]."Plant"Cell"and"Environment,"2020,"43(10):"2428-2442.
- 肖艷華,"鄒智,"趙永國,"郭安平,"張麗."油莎豆乙酰乳酸合酶基因CeALS的克隆與分析[J]."生物技術通報,"2022,"38(4):"184-192.XIAO"Y"H,"ZOU"Z,"ZHAO"Y"G,"GUO"A"P,"ZHANG"L."Molecular"cloning"and"characterization"of"an"acetolactate"synthase"gene"(CeALS)"from"tigernut"(Cyperus"esculentus"L.)[J]."Biotechnology"Bulletin,"2022,"38(4):"184-192."(in"Chinese)
- 鄒智,"肖艷華,"張麗,"趙永國."油莎豆5-烯醇式丙酮酰莽草酸-3-磷酸合酶基因CeEPSPS的克隆與分析[J]."熱帶作物學報,"2023,"44(1):"26-34.ZOU"Z,"XIAO"Y"H,"ZHANG"L,"ZHAO"Y"G."Cloning"and"characterization"of"CeEPSPS,"a"gene"encoding"5-enolpyruvy lshikimate-3-phosphate"synthase"from"tigernut"(Cyperus"esculentus"L.)[J]."Chinese"Journal"of"Tropical"Crops,"2023,"44(1):"26-34."(in"Chinese)
[17]"ZOU"Z,"ZHAO"Y"G,"ZHANG"L,"XIAO"Y"H,"GUO"A"P."Analysis"of"Cyperus"esculentus"SMP"family"genes"reveals"lineage-specific"evolution"and"seed"desiccation-like"transcript"accumulation"during"tuber"maturation[J]."Industrial"Crops"and"Products,"2022,"187:"115382.
[18]"ZOU"Z,"XIAO"Y"H,"ZHANG"L,"ZHAO"Y"G."Analysis"of"Lhc"family"genes"reveals"development"regulation"and"diurnal"fluctuation"expression"patterns"in"Cyperus"esculentus,"a"Cyperaceae"plant[J]."Planta,"2023,"257(3):"59.
[19]"ZOU"Z,"ZHENG"Y"J,"CHANG"L"L,"ZOU"L"P,"ZHANG"L,"MIN"Y,"ZHAO"Y"G."TIP"aquaporins"in"Cyperus"esculentus:"Genome-wide"identification,"expression"profiles,"subcellular"localizations,"and"interaction"patterns[J]."BMC"Plant"Biology,"2024,"24(1):"298.
[20]"TURESSON"H,"MARTTILA"S,"GUSTAVSSON"K"E,"HOFVANDER"P,"OLSSON"M"E,"BüLOW"L,"STYMNE"S,"CARLSSON"A"S."Characterization"of"oil"and"starch"accumulation"in"tubers"of"Cyperus"esculentus"var."sativus"(Cyperaceae):"a"novel"model"system"to"study"oil"reserves"in"nonseed"tissues[J]."American"Journal"of"Botany,"2010,"97(11):"1884-1893.
[21]"鄒智,"趙永國,"張麗,"孔華,"郭運玲,"郭安平."基于單分子實時測序的油莎豆全長轉錄組分析[J]."中國油料作物學報,"2021,"42(2):"229-235.ZOU"Z,"ZHAO"Y"G,"ZHANG"L,"KONG"H,"GUO"Y"L,"GUO"A"P."Single-molecule"real-time"(SMRT)-based"full-length"transcriptome"analysis"of"tigernut"(Cyperus"esculentus"L.)[J]."Chinese"Journal"of"Oil"Crop"Sciences,"2021,"43(2):"229-235."(in"Chinese)
[22]"徐碩,"鄒智,"肖艷華,"張麗,"孔華,"郭靜遠,"郭安平."油莎豆塊莖油脂積累相關基因CeWRI1的克隆與功能分析[J]."熱帶作物學報,"2022,"43(5):"923-929.XU"S,"ZOU"Z,"XIAO"Y"H,"ZHANG"L,"KONG"H,"GUO"J"Y,"GUO"A"P."Cloning"and"functional"characterization"of"CeWRI1,"a"gene"involved"in"oil"accumulation"from"tigernut"(Cyperus"esculentus"L.)"tubers[J]."Chinese"Journal"of"Tropical"Crops,"2022,"43(5):"923-929."(in"Chinese)
[23]"ZOU"Z,"ZHENG"Y"J,"ZHANG"Z"T,"XIAO"Y"H,"XIE"Z"N,"CHANG"L"L,"ZHANG"L,"ZHAO"Y"G."Molecular"characterization"of"oleosin"genes"in"Cyperus"esculentus,"a"Cyperaceae"plant"producing"oil"in"underground"tubers[J]."Plant"Cell"Reports,"2023,"42(11):"1791-1808.
[24]"喬雪瑩,"鄭玉皎,"陽江華,"曾長英,"鄒智."橡膠樹HbPIP1;1基因的克隆及蛋白的亞細胞定位與多聚化分析[J]."熱帶作物學報,"2022,"43(12):"2405-2412."QIAO"X"Y,"ZHENG"Y"J,"YANG"J"H,"ZENG"C"Y,"ZOU"Z."Gene"cloning,"subcellular"localization"and"multimerization"analysis"of"HbPIP1;1"from"Hevea"brasiliensis[J]."Chinese"Journal"of"Tropical"Crops,"2022,"43(12):"2405-2412."(in"Chinese)
[25]"鄒智,"喬雪瑩,"鄭玉皎,"陽江華."橡膠樹HbPIP2;3的亞細胞定位與多聚化分析[J]."熱帶作物學報,"2024,"45(3):"443-449.ZOU"Z,"QIAO"X"Y,"ZHENG"Y"J,"YANG"J"H."Subcellular"localization"and"multimerization"analyses"of"HbPIP2;3,"an"efficientnbsp;water"transporter"from"Hevea"brasiliensis[J]."Chinese"Journal"of"Tropical"Crops,"2024,"45(3):"443-449."(in"Chinese)
[26]"DANIELSON"J"A,"JOHANSON"U."Unexpected"complexity"of"the"aquaporin"gene"family"in"the"moss"Physcomitrella"patens[J]."BMC"Plant"Biology,"2008,"8(1):"45.
[27]"ZELAZNY"E,"MIECIELICA"U,"BORST"J"W,"HEMMINGA"M"A,"CHAUMONT"F."An"N-terminal"diacidic"motif"is"required"for"the"trafficking"of"maize"aquaporins"ZmPIP2;4"and"ZmPIP2;5"to"the"plasma"membrane[J]."Plant"Journal,"2009,"57(2):"346-355.
[28]"FETTER"K,"VAN"WILDER"V,"MOSHELION"M,"CHAUMONT"F."Interactions"between"plasma"membrane"aquaporins"modulate"their"water"channel"activity[J]."The"Plant"Cell,"2004,"16(1):"215-228.
[29]"ZELAZNY"E,"BORST"J"W,"MUYLAERT"M,"BATOKO"H,"HEMMINGA"M"A,"CHAUMONT"F."FRET"imaging"in"living"maize"cells"reveals"that"plasma"membrane"aquaporins"interact"to"regulate"their"subcellular"localization[J]."Proceedings"of"the"National"Academy"of"Sciences,"2007,"104(30):"12359-12364.
[30]"NIEMEYER"P"W,"IRISARRI"I,"SCHOLZ"P,"SCHMITT"K,"VALERIUS"O,"BRAUS"G"H,"HERRFURTH"C,"FEUSSNER"I,"SHARMA"S,"CARLSSON"A"S,"DE"VRIES"J,"HOFVANDER"P,"ISCHEBECK"T."A"seed-like"proteome"in"oil-rich"tubers[J]."Plant"Journal,"2022,"112(2):"518-534.
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
