辣椒雄性不育系的篩選與鑒定
2024-12-31 00:00:00李彩朝龍希洋李承濤譚萍
鄉(xiāng)村科技 2024年7期

摘 要:將細胞質(zhì)雄性不育機制應(yīng)用于辣椒育種工作,可加快辣椒雜交種的生產(chǎn)及利用雜種優(yōu)勢來提高辣椒產(chǎn)量和品質(zhì),并降低制種成本。在眾多F2代辣椒中篩選鑒定出雄性不育源(HG),通過切片分析不育株辣椒細胞組織的發(fā)育情況,進一步篩選鑒定出辣椒的雄性不育株。同時,選用性狀優(yōu)良的父本與篩選出的不育株進行雜交、回交育種,為辣椒三系育種提供理論基礎(chǔ)。
關(guān)鍵詞:辣椒;雄性不育;切片;雜交
中圖分類號:S641.303 文獻標志碼:B 文章編號:1674-7909(2024)7-59-4
DOI:10.19345/j.cnki.1674-7909.2024.07.013
0 引言
辣椒(Capsicum annuum L.)為茄科(Solanaceae)辣椒屬(Capsicum)一年生或有限多年生草本或灌木植物,因其獨特的營養(yǎng)價值而深受消費者喜愛[1]。2022年,貴州省辣椒種植面積約為36.2萬 hm2。
細胞質(zhì)雄性不育(cytoplasmic male sterility,CMS)在辣椒F1代雜交育種中發(fā)揮著重要作用。雄性不育(male sterility)是指植物在生長發(fā)育過程中,產(chǎn)生的雄配子沒有正常可育的遺傳功能[2]。植物雄性不育是一種對植物自身不利的性狀,但因其能與正常花粉受精結(jié)實,在雜交育種方面又具有獨特的優(yōu)勢[3]。植物雄性不育主要特征是小孢子發(fā)育異常,花粉萌發(fā)逐漸變?nèi)跎踔羻适劝l(fā)功能[4]。雄性不育現(xiàn)象基本發(fā)生在植物界。目前,至少在43科162屬320個種和297個種間雜交種中發(fā)現(xiàn)雄性不育現(xiàn)象[5]。
作物雜種優(yōu)勢在生產(chǎn)中發(fā)揮著重要作用,后代產(chǎn)量比親本產(chǎn)量高15%~50% 。目前,雜交育種已被廣泛應(yīng)用于農(nóng)業(yè)生產(chǎn)中,如在辣椒、玉米、油菜籽、高粱、向日葵等的生產(chǎn)中均用到雜種優(yōu)勢。然而,作物花朵去雄問題是限制雜種優(yōu)勢廣泛應(yīng)用的難題之一。作物花朵去雄會消耗大量的人力、物力和財力,而要解決這個問題的最好方法是篩選雄性不育系來進行育種[6]。在辣椒細胞質(zhì)雄性不育方面,已有學(xué)者進行了大量研究。例如,李瑩瑩等[7]認為敗育在減數(shù)分裂的任何時期都有可能發(fā)生,且敗育方式不同,主要表現(xiàn)形式為絨氈層異常發(fā)育;Wang等[8]在不育系138A中篩選出強候選基因。目前,此技術(shù)辣椒三系育種中的實際應(yīng)用還有待進一步研究。基于此,筆者通過田間試驗來選育出新的辣椒不育系,以期為辣椒產(chǎn)業(yè)的發(fā)展提供一定參考。
1 試驗材料與方法
1.1 試驗地概況
于2020—2021年在海南大學(xué)農(nóng)科基地開展試驗。該地屬熱帶季風(fēng)氣候區(qū),年平均氣溫為24.8 ℃。由于溫度適宜,1 a可種植兩茬辣椒。
1.2 試驗材料
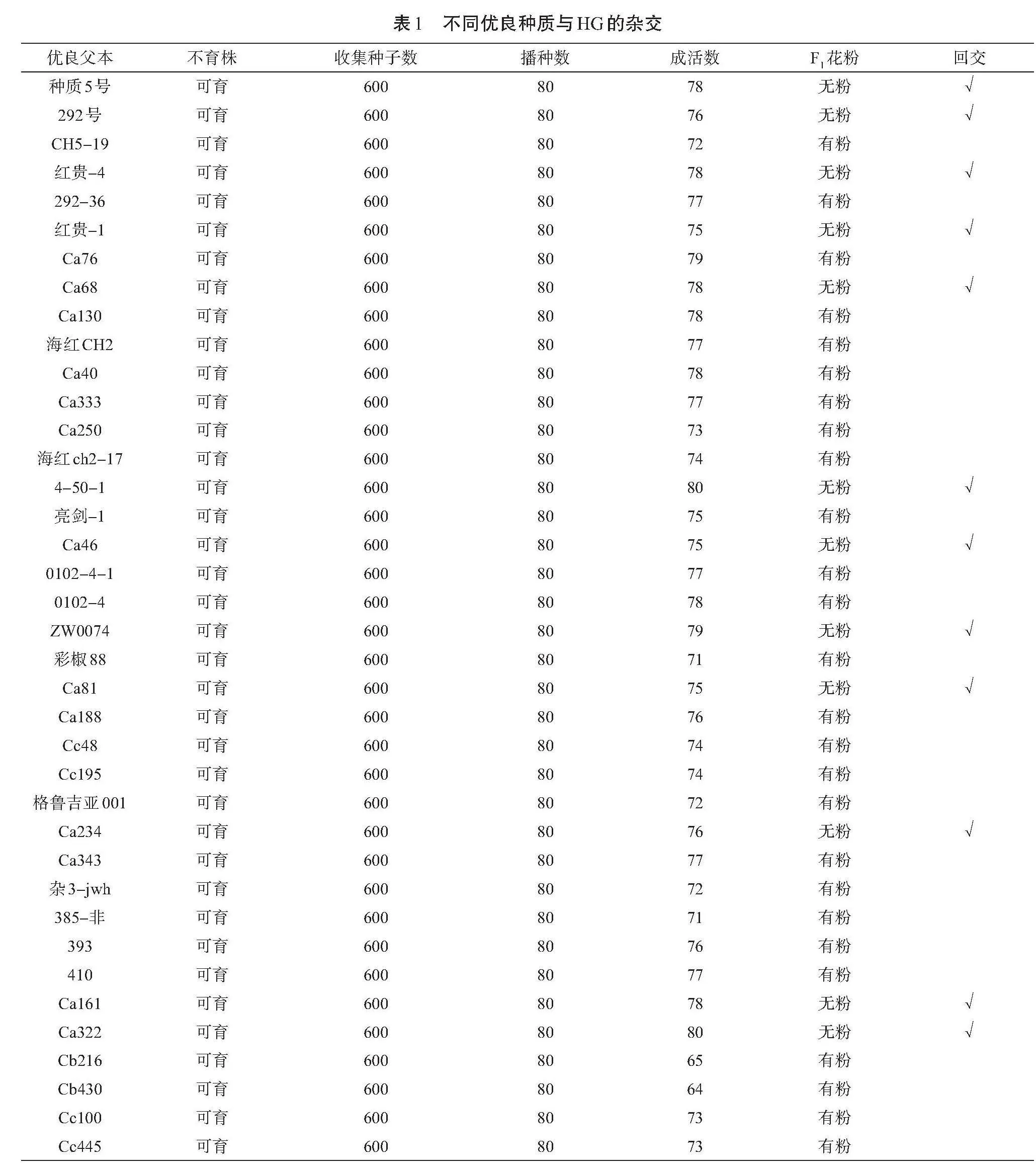
所用到的試驗材料均為筆者所在的課題組在網(wǎng)上購買的純種辣椒,用于篩選不育材料的辣椒品種有HG、SQ、CJHJ、LJ、LST,用于雜交試驗的辣椒品種有種質(zhì)5號、292號、CH5-19、紅貴-4、292-36、紅貴-1、Ca76、Ca68、Ca130、海紅CH2、Ca40、Ca333、Ca250、海紅ch2-17、4-50-1、亮劍-1、Ca46、0102-4-1、0102-4、ZW0074、彩椒88、Ca81、Ca188、Cc48、Cc195、格魯吉亞001、Ca234、Ca343、雜3-jwh、385-非、393、410、Ca161、Ca322、Cb216、Cb430、Cc100、Cc445。在實驗室進行試驗時,根據(jù)辣椒栽培種和試驗?zāi)康倪M行編號。
1.3 試驗方法
1.3.1 播種育苗
對5個不同品種(HG、SQ、CJHJ、LJ、LST)的辣椒種子進行篩選,剔除其中不飽滿和生活力低的種子,并對辣椒種子進行浸種催芽。每個辣椒品種催芽100粒種子,并恒溫浸種。浸種后放在溫室內(nèi)進行催芽,催芽時要及時換水,并用紗布包裹后搓洗,待種子露白后播種,播種時使用的是24孔穴盤。播種后,將穴盤放到恒溫實驗室內(nèi)進行培育,并定期澆水。待幼苗長出6~8片真葉后移栽到基地,每個品種選擇80株(共400株)長勢好的辣椒苗進行移栽,移栽前進行蹲苗,移栽后要及時澆水、施肥、防治病蟲害等。
1.3.2 篩選不育材料
在移栽的400株辣椒中,篩選出花朵沒有花粉的辣椒植株(前期只能根據(jù)辣椒花朵是否有花粉來進行初步篩選)。在篩選出無花粉的辣椒植株后,用鏡檢法對辣椒花蕾進行檢測,觀察其是否有花粉,從而初步判斷是否為不育的辣椒植株。
細胞質(zhì)雄性(CMS)不育株能用于辣椒育種,細胞核雄性(GMS)不育株不能用于辣椒育種。所以,為了進一步確定篩選出的辣椒不育株是GMS不育株還是CMS不育株,要對不育株進行雜交授粉,用其他可育辣椒與不育株進行雜交,收集后代種子,并進行催芽種植,通過有無花粉來判定不育株是GMS不育株還是CMS不育株。
1.4 花粉鑒定
當移栽的400株辣椒完全開花后,分別鑒別其是否有花粉,最終只找到一株(HG)沒有花粉,如圖1所示。辣椒不育系植株的外觀與可育系植株不存在差別,二者的主要區(qū)別是有無花粉。HG可育株花瓣在未開放時很飽滿,待花蕾開放后,能清晰看到可育株有很多花粉[見圖1(a)];HG不育株花瓣還沒有開放時呈皺縮狀態(tài),在花瓣完全打開后,雄蕊干癟,沒有花粉[見圖1(b)]。
2 試驗結(jié)果與分析
2.1 雜交試驗分析
以辣椒HG不育株為母本,選用優(yōu)良的辣椒品種作為父本進行雜交,得到F1代。選擇沒有花粉的株系,用其與父本連續(xù)回交,得到F2代、F3代、F4代。通過這種方式得到的后代細胞核具有父本細胞核的優(yōu)良特征,即含有隱性的不育核基因(ss)。這是因為母性遺傳是細胞質(zhì)雄性不育的特性[9],本身就有不育基因,可以把不育的特征回交轉(zhuǎn)育到優(yōu)良的父本中[10],從而篩選出穩(wěn)定的不育系和保持系。不同優(yōu)良種質(zhì)與HG的雜交情況見表1。
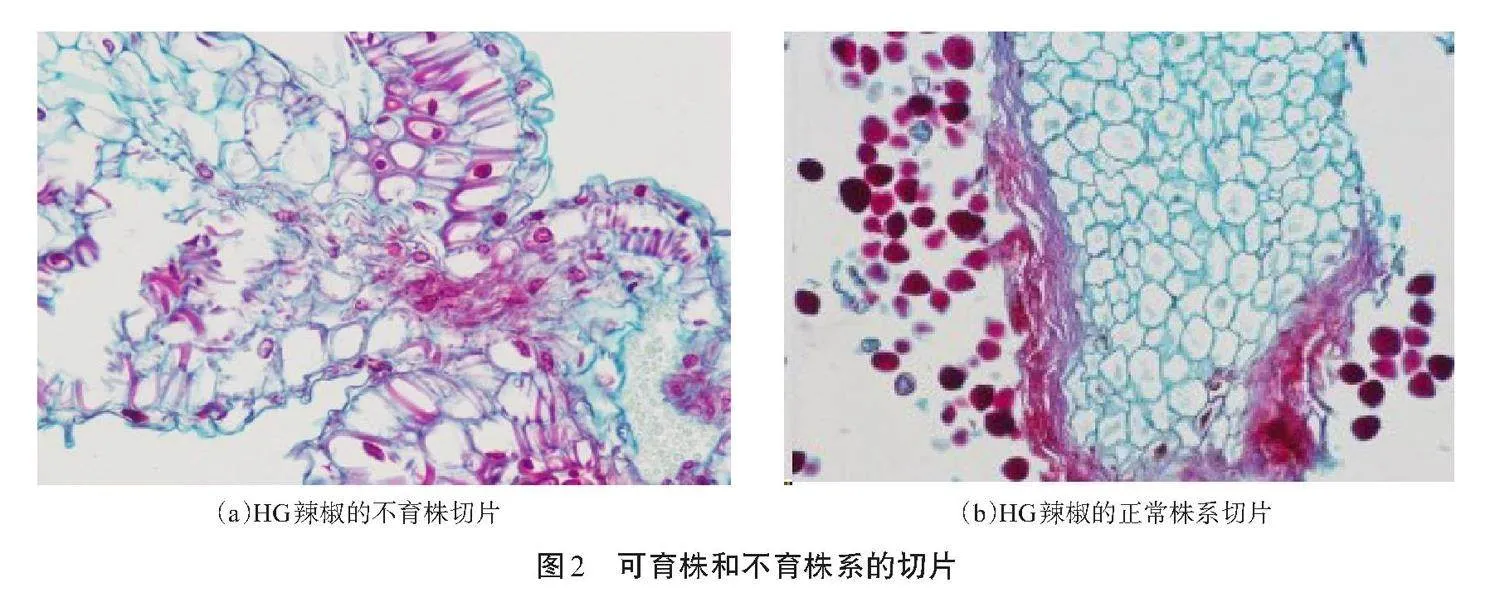
2.2 切片分析
可育株和不育株系的切片如圖2所示。由圖2可知,辣椒敗育發(fā)生在減數(shù)分裂過程中,小孢子母細胞發(fā)育受辣椒細胞絨氈層的影響,導(dǎo)致小孢子母細胞不能正常發(fā)育,辣椒細胞絨氈層因過度膨大而嚴重擠壓花粉粒細胞,導(dǎo)致生長發(fā)育過程中的減數(shù)分裂過程發(fā)生異常,從而使辣椒不能形成正常的花粉粒,導(dǎo)致辣椒出現(xiàn)敗育現(xiàn)象,且沒有花粉的辣椒花蕾切片幾乎沒有花粉粒形成[見圖2(a)],而在減數(shù)分裂后期,正常辣椒花蕾的切片有明顯的花粉粒[見圖2(b)],進一步驗證篩選出的、沒有花粉的辣椒植株是雄性不育辣椒株系。
3 結(jié)束語
辣椒敗育發(fā)生在減數(shù)分裂過程中,由于小孢子母細胞發(fā)育過程受辣椒細胞絨氈層的影響,導(dǎo)致小孢子母細胞不能正常發(fā)育,細胞絨氈層過度膨大會嚴重擠壓花粉粒細胞,導(dǎo)致減數(shù)分裂發(fā)生異常,從而使辣椒不能形成正常的花粉粒。上述過程是辣椒細胞質(zhì)雄性不育的形成過程[11],該研究中的辣椒可育株和不育株的切片分析結(jié)果與上述論證一致。
筆者在5個不同品種(共400株辣椒)中篩選出名為HG的辣椒雄性不育株。通過可育株系和不育株系同一個發(fā)育時期的切片對比分析可知,篩選出的HG是雄性不育辣椒株系,再利用篩選出的不育源與優(yōu)良的父本進行回交,并連續(xù)回交3~4代,即可培育出雄性不育系。這與劉紫垠等[12]的研究結(jié)果基本一致;不同的是,劉紫垠等[12]研究時用到的辣椒不育源是引進的,而該研究的不育源是通過田間試驗篩選得到的。下一步,要篩選出保持系和恢復(fù)系,以期為辣椒三系育種提供一定的技術(shù)支持。
辣椒不育系篩選在辣椒三系育種中具有非常重要的作用,有很高的經(jīng)濟效益。利用細胞質(zhì)雄性不育法育種可降低成本,提高辣椒產(chǎn)量和品質(zhì)。目前,隨著高通量測序技術(shù)的快速發(fā)展,為辣椒線粒體基因組、轉(zhuǎn)錄組、蛋白組的測序和分析提供強有力的技術(shù)支持。未來,可結(jié)合轉(zhuǎn)錄組、代謝組、蛋白組等組學(xué)和辣椒線粒體基因組來研究細胞質(zhì)雄性不育,從而挖掘出更多有價值的信息,為辣椒育種提供更多有價值的參考。
參考文獻:
[1]王本啟,黃樹蘋,陳霞,等.辣椒細胞質(zhì)雄性不育分子生物學(xué)組學(xué)研究進展[J].中國蔬菜,2023(5):20-29.
[2]張銳,尚偉,許旭明.辣椒雄性不育的選育及利用研究進展[J].分子植物育種,2020(18):6143-6157.
[3]時子文,何青,趙卓凡,等.玉米雄性不育資源的發(fā)掘與利用[J].遺傳,2022(2):134-152.
[4]廖麗娟.細胞質(zhì)雄性不育辣椒花粉敗育的轉(zhuǎn)錄組學(xué)研究[D].長沙:湖南師范大學(xué).2017.
[5]郭爽,常紹東,黃貞等.辣椒雄性不育研究進展[J].辣椒雜志,2012(2):1-5.
[6]李彩朝,ZAKARIA G,金威恒,等.辣椒新細胞質(zhì)雄性不育線粒體基因組的測序及分析[J].分子植物育種,2022(16):5250-5257.
[7]李瑩瑩,魏佑營,張瑞華,等.辣椒雄性不育系與可育系小孢子發(fā)生的細胞學(xué)觀察[J].植物研究,2006(4):411-415.
[8]WANG P,LU Q H,AI Y X,et al.Candidate Gene Selection for Cytoplasmic Male Sterility in Pepper (Capsicum annuum L.) through Whole Mitochondrial Genome Sequencing[J].Int J Mol Sci,2019(3):578.
[9]WANG C D,LINA L,NADèGE A,et al.The radish ogura fertility restorer impedes translation elongation along its cognate CMS-causing mRNA[J].Proc Natl Acad Sci U S A.2021(35):1-9.
[10]王作義,楊鳳梅,王志強,等.辣椒雄性不育兩用系選育轉(zhuǎn)育及利用[J].北方園藝,1998(1):10-12.
[11]DASKALOFF S.A male sterile pepper (C. annuum L.) mutant[J].Theor Appl Genet,1968(8):370-372.
[12]劉紫垠,梁宏衛(wèi),張雪玲,等.辣椒雄性不育系304A的選育[J].辣椒雜志,2018(1):11-13.
基金項目:貴陽市科技計劃項目(筑科項目〔2022〕3號)。
作者簡介:李彩朝(1994—),男,碩士,助理農(nóng)藝師,研究方向:蔬菜育種。
通信作者:譚萍(1986—),女,碩士,農(nóng)藝師,研究方向:蔬菜育種及栽培技術(shù)推廣。
