呂梁黑豬BACE2基因克隆及表達分析
2024-12-31 00:00:00楊梓杰張晉趙丹
安徽農學通報 2024年22期



摘要" 為探討呂梁黑豬BACE2基因特點和結構,本研究采用PCR技術對該基因進行克隆,并對所編碼的蛋白質開展生物信息學分析,分析內容包括蛋白質的理化性質、信號肽、親疏水性、跨膜結構、序列比對、二、三級結構預測、亞細胞定位預測,以及互作關系;通過實時熒光定量PCR(qPCR)技術,研究BACE2基因在呂梁黑豬的心臟、肝臟、脾臟、肺臟、腎臟、肌肉和脂肪組織中的表達情況。生物信息學結果顯示,該黑豬BACE2基因的編碼序列大小(CDS)為1 545"bp,包含514個氨基酸;BACE2蛋白是一種疏水性蛋白,無信號肽,具有明顯的跨膜作用,是一種非分泌型的蛋白質;序列比對發現呂梁黑豬同山羊、牛親緣關系較近,同家鼠親緣關系較遠;蛋白的二級結構由無規則卷曲(49.03%)、α-螺旋(18.48%)、延伸連(27.24%)和β-轉角(5.25%)構成,主要分布于內質網(55.6 %)。qPCR 結果顯示,其腎臟組織中BACE2基因的表達水平最高,肌肉組織中的表達量最低。研究結果揭示了BACE2蛋白擁有的多種獨特屬性及BACE2基因的分布特點,為該蛋白更深層次的科學探究提供參考。
關鍵詞" 呂梁黑豬;BACE2基因;生物信息學
中圖分類號" S85"""""" 文獻標識碼" A"""""" 文章編號" 1007-7731(2024)22-0043-07
DOI號" 10.16377/j.cnki.issn1007-7731.2024.22.010
基金項目 山西省教育科學“十四五”規劃課題“高質量教育體系下高校動醫專業基礎課程實驗數字化教學探索與實踐”(SZH-230078)。
作者簡介 楊梓杰(2003—)男,山西太原人,從事動物醫學研究。
通信作者 趙丹(1986—),男,山西太原人,碩士,講師,從事動物生物化學研究。
收稿日期 2024-08-07
Cloning and expression analysis of BACE2 gene in Lyuliang black pig
YANG Zijie""" ZHANG Jin""" ZHAO Dan
(Jinzhong College of Information, Taigu 030800, China)
Abstract" In order to investigate the characteristics and structure of the BACE2 gene in Lyuliang black pigs, the BACE2 gene was cloned by PCR technology,and the encoded protein was analyzed by bioinformatics, including physical and chemical properties, signal peptide, hydrophobicity, transmembrane structure, sequence alignment, secondary and tertiary structure prediction, subcellular localization, and interaction. The expression of the BACE2 gene in the heart, liver, spleen, lung, kidney, muscle and adipose tissue of this pigs was measured by real-time quantitative PCR. The results of bioinformatics showed that the coding sequence (CDS) of BACE2 gene size was 1 545"bp, containing 514"amino acids. BACE2 protein is a hydrophobic protein, no signal peptide, has obvious transmembrane function, and is a non-secretory protein. Sequence alignment showed that the pig had a close genetic relationship with goat and cattle, and the furthest genetic relationship with Mus musculus. The secondary structure of the protein was composed of random curl (49.03%), α-helix (18.48%), extension link (27.24%) and β-corner (5.25%), mainly distributed in the endoplasmic reticulum (55.6%).qPCR results showed that the expression level of BACE2 gene was the higher in kidney tissue and the lower in muscle tissue of this pigs. The results revealed a variety of unique properties of BACE2 protein and the distribution characteristics of BACE2 gene, which provided a reference for further scientific exploration of this protein.
Keywords" Lyuliang black pigs; BACE2 gene; bioinformatics
豬是一種脊椎動物,也是雜食類哺乳動物。由于品種的不同,其體貌特征也各不相同,通常以耳大、頭長、四肢短小、鼻直、身體肥壯和腰背窄為主要形體特征[1]。呂梁黑豬是引進生豬進行自繁自養得到的品種,其全身體毛黑色、結構勻稱、四肢健壯,頭型稍長,額部有橫行皺紋,具有肉質鮮美、抗病力強等特點[2]。BACE2(Beta-site app-cleaving enzyme 2,BACE2)是BACE1的同源物,是一種θ分泌酶,也是一種條件性β分泌酶,其能夠水解β淀粉樣前體蛋白(β-amyloid precursor protein,APP)產生β淀粉樣蛋白(Amyloid-β peptide,Aβ)[3]。在體外,發現BACE2是一種特別活躍的Aβ降解蛋白酶(Aβ-degrading proteases,AβDP),其催化效率超過了除胰島素降解酶(Insulin degrading enzymes,IDE)以外的已知的 AβDPs[3],是治療或預防阿爾茲海默癥(Alzheimer’s disease,AD)的一種強有力的候選藥物。謝建等[4]研究表明,敲除BACE2可抑制細胞侵襲、遷移和腫瘤生長,過表達BACE2可促進細胞間質轉化和增殖。王會志[5]通過BACE2激活NF-κB通路促進膠質瘤惡性進展的機制研究,發現BACE2在膠質瘤的發生與發展中扮演關鍵角色,敲低BACE2可減弱膠質瘤侵襲遷移及增殖能力,是治療該疾病的潛在靶點。
目前,將呂梁黑豬作為研究對象的相關研究相對較少,在其肌肉發育和脂肪沉積中,BACE2基因的作用機制有待進行充分研究。因此,對其BACE2基因的生物信息學進行分析,有助于深入理解該物種在遺傳層面的特性,尤其是在肌肉發育和脂肪沉積等性狀方面。本研究采用PCR技術對BACE2基因進行克隆,并對所編碼的蛋白質開展生物信息學分析,通過實時熒光定量PCR(qPCR)技術,研究BACE2在呂梁黑豬的心臟、肝臟、脾臟、肺臟、腎臟、卵巢、肌肉和脂肪組織中的表達情況,為BACE2蛋白更深層次的科學探究提供參考,為該豬種的保種和改良工作提供分子生物學層面的數據參考。
1 材料與方法
1.1 試驗材料
在山西呂梁萊蕪黑豬養殖基地,挑選3頭體質強健的呂梁黑豬作為試驗對象。屠宰后,迅速采集其心臟、肝臟、脾臟、肺臟、腎臟、卵巢、肌肉以及脂肪8個組織。為確保組織的完整性和活性,采集后立即放入液氮中進行冷凍保存。隨后,將組織樣本運送回實驗室,轉移至-80"℃冰箱中,備用。
1.2 試驗儀器與試劑
PCR儀(NEW VeritiPro)、電泳儀(GH600E)、超微量分光光度計(NanoDrop 2000),凝膠成像系統(Invitrogen E-Gel Power Snap G8100);DL2000 DNA Marker、LATaq DNA聚合酶以及M-MLV反轉錄測試盒,均購自寶日醫生物技術(北京)有限公司。
1.3 試驗方法
1.3.1 總RNA提取及反轉錄 采用Trizol法提取組織的總RNA。采用超微量分光光度計NanoDrop 2000檢測RNA的質量和濃度,其OD260和OD280在1.8~2.0。通過以上處理,得到一個完備且有代表性的核酸序列數據,滿足相關必要的標準后,按照M-MLV反轉錄測試盒的說明書進行反轉錄,反轉錄得到的cDNA樣品存置于-20"℃的冰箱,備用。
1.3.2 BACE2基因引物設計與合成 基于GenBank提供的BACE2基因序列(登記編號:XP 020926661.1.1),設計用于克隆和實時熒光定量PCR(qPCR)的引物。引物序列送至西安擎科生物科技公司合成,詳情如表1所示。
1.3.3 BACE2基因的克隆與測序 PCR試驗以呂梁黑豬肝臟組織cDNA為模板進行特異性擴增。反應總體系為25 μL,具體組成:ddH2O 8.5 μL,2×TaqPCR Master Mix 12.5 μL,cDNA 2 μL,上下游引物各1 μL(濃度10 μmol/L)。擴增條件:首先94"℃預變性30"s;然后進行35個擴增循環,每個循環98"℃變性10"s,56.02"℃退火30"s,72"℃延長1"min,循環結束后72"℃延伸2"min。將產物保存于4"℃冰箱。為驗證擴增結果的準確性,采用高達瓊脂糖凝膠電泳及凝膠成像系統對目的條帶進行檢測,將符合預期的條帶送至西安擎科生物科技公司進行測序分析。
1.3.4 BACE2基因的生物信息學分析 采用生物信息學方法對測序分析得到的氨基酸殘基序列及其次級結構進行了深入研究,以揭示蛋白質的空間結構特點,為后續的實驗研究提供了一定的數據支撐。使用Protparam工具,分析蛋白質理化性質。在https//prosite.expasy.org網站查找蛋白質結構的研究資料。采用基于密度泛函原理的策略,對蛋白質的分子構造與其綁定作用之間的聯系進行初步研究。通過SignaIP軟件預測蛋白質信號肽;通過https://web.expasy.org/protscale預測蛋白質的親疏水性;利用TMHMM2.0軟件進行蛋白質的跨膜結構預測;利用NCBI軟件里的BLASTX工具進行蛋白質同源性比對,利用MEGA7軟件繪制蛋白質演變樹狀結構。通過https://predictprotein.org預測蛋白質的次級結構;通過https://swissmodel.expasy.org預測蛋白質三級結構。通過PSORT 在線軟件進行蛋白亞細胞定位。基于以上數據,應用機械學習和人工神經網絡的技術,成功建立了一個以計算機模擬為核心的三維蛋白質模型。利用在線軟件STRING對來自多個數據平臺的蛋白質氨基酸組成及其之間的功能性關聯進行詳細的比較分析,構建BACE2蛋白互作網絡。
1.3.5 實時熒光定量PCR 以8個組織的cDNA為模板,GAPDH為內參,進行qPCR檢測。試驗的反應總體系為20 μL,詳細組分:10 μL的2× SYBR Green Pro Taq HS Premix,上游和下游引物(濃度為10 μmol/μL)各0.4 μL,cDNA模板1 μL,以及8.2 μL ddH2O。反應程序:首先進行95"℃預變性30"s;隨后進行40個擴增循環,每個循環包括95"℃變性5"s,57"℃退火30"s,65"℃延伸5"s;循環結束后95"℃延伸5"min。
1.4 數據處理
用Excel軟件和2-ΔΔCt方法對qPCR結果進行統計分析。
2 結果與分析
2.1 BACE2基因克隆與測序
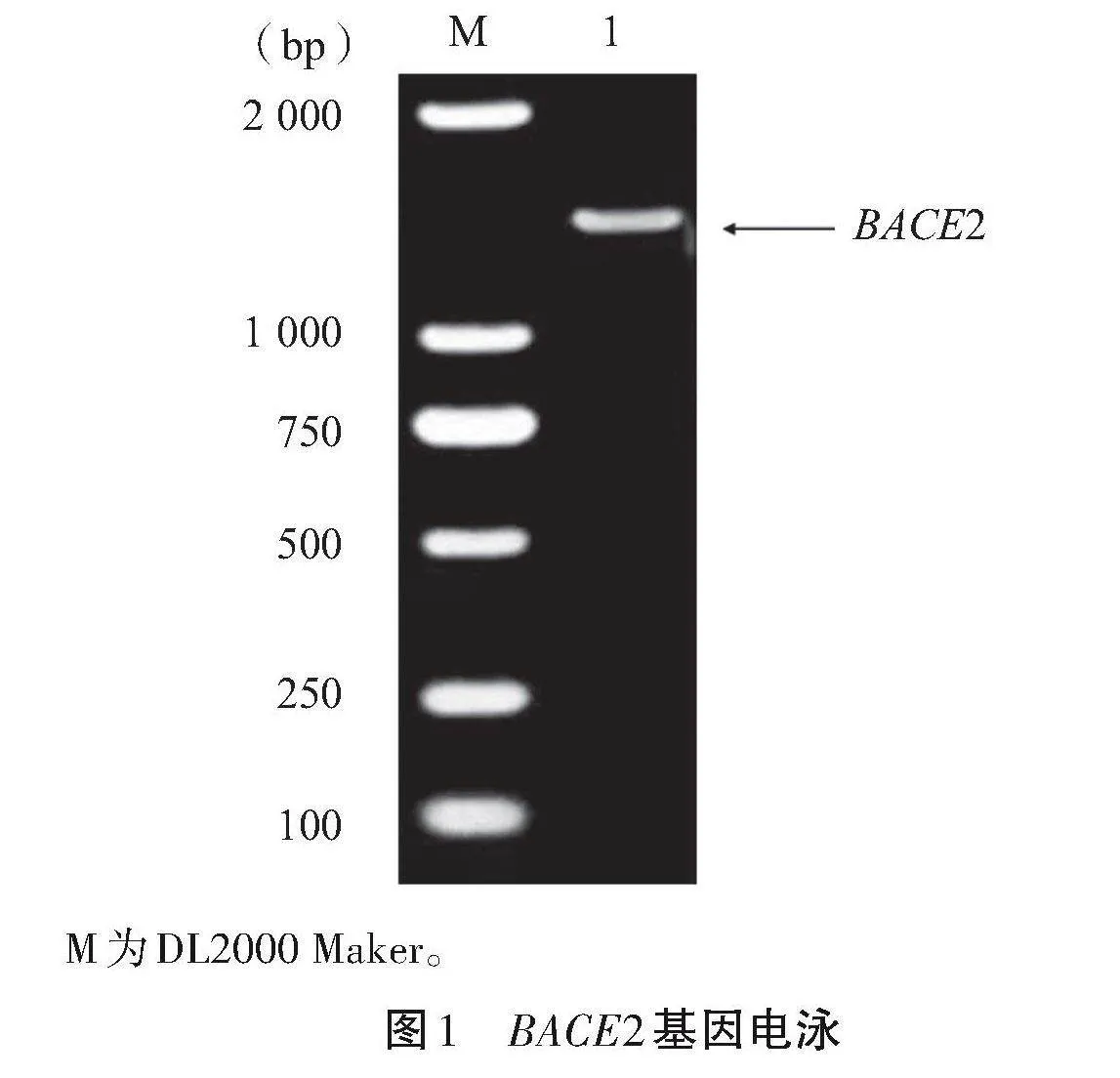
以cDNA為模板,利用特異性引物進行PCR擴增該基因片段,1%瓊脂糖凝膠電泳檢測結果顯示,目的條帶大小約為1 500"bp,符合預期(圖1)。收集測序數據后,采用DNAstar軟件中的Seqman子程序來完成序列的拼接工作。將拼接序列與NCBI數據庫中BACE2基因的CDS區使用BLAST軟件進行比對,結果表明,呂梁黑豬的BACE2基因編碼區域長度達到了1 545"bp。基于開放閱讀框(ORF Finder)的計算,這一序列被編碼為514個氨基酸。
M為DL2000 Maker。
2.2 BACE2蛋白的理化性質
使用 Protparam 軟件分析該蛋白理化性質,結果如表2所示,BACE2蛋白的分子式C2541H3948N646O725S17、氨基酸數量為514個,其分子量為55 692.21 Da、理論等電點(PI)5.27,在氨基酸組分中含量最高的是亮氨酸(Leu),占總氨基酸數的11.7%,組氨酸(Cys)含量最低,占總氨基酸數的0.6%。BACE2蛋白帶正、負電荷殘基總數分別為38、45個,脂肪系數(AI)98.52、不穩定系數()44.41、總平均親水性指數(GRAVY)0.183。
2.3 BACE2蛋白的信號肽
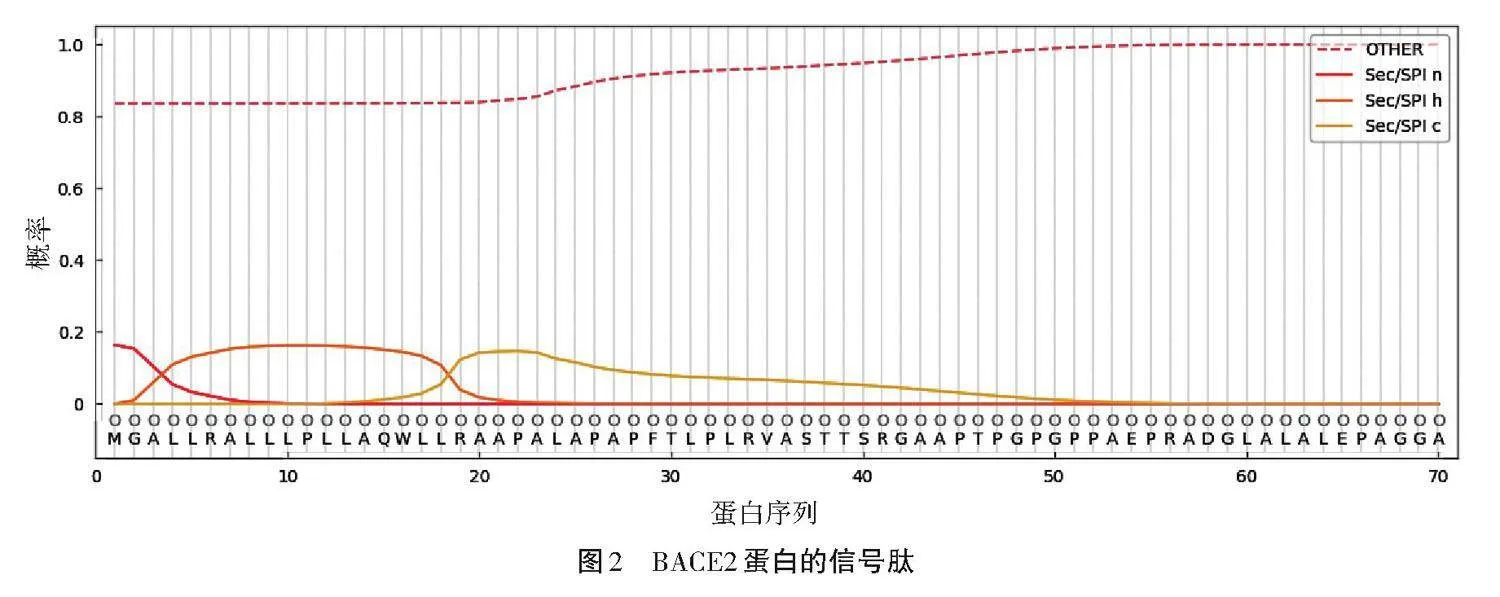
信號肽的構造可細分為3個主要部分。首先是n區(n-region),是一個富含正電荷的氨基酸序列;緊接著是疏水區(h-region),由9個以上的中性氨基酸組成,共同構建了一個疏水性的界面;最后加工區(c-region),是信號肽酶作用的關鍵區域,負責切割信號肽,從而完成其生物學功能。
使用了SignaIP工具,對BACE2蛋白的信號肽狀態進行預測,結果如圖2所示。經過分析,該黑豬BACE2蛋白的信號肽實際上是不存在的,并且未形成跨膜的結構。這一發現有利于更深入地了解該基因編碼產物在細胞中的位置和其調節機制。BACE2蛋白可能作為一種酶,直接與細胞質基質中的代謝底物發生相互作用,促進細胞內產生各種化學變化,被歸類為非分泌型蛋白質。
2.4 BACE2蛋白的親疏水性
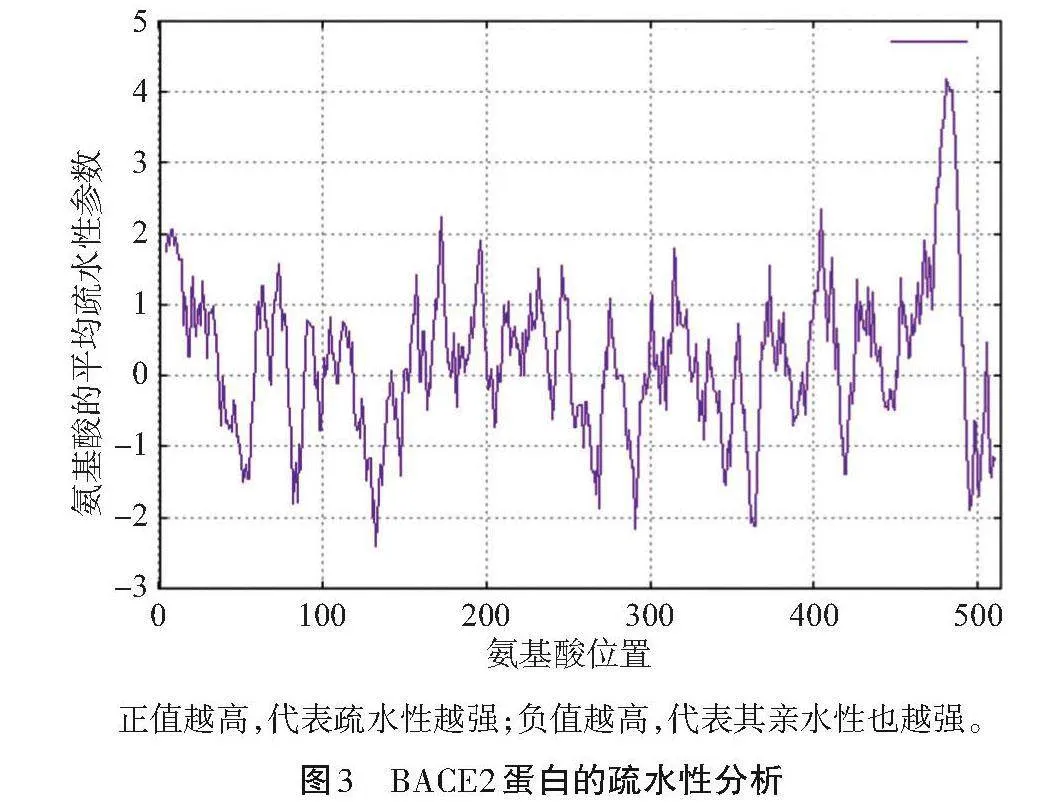
通過在線分析平臺研究BACE2蛋白的疏水特性,結果如圖3所示。研究發現,黑豬的BACE2蛋白疏水性氨基酸的數量多于親水性氨基酸,其中第133號氨基酸的位置展現了最高的疏水性(最小的親水性);在第481號氨基酸的位置,其疏水性最低(親水性最高)。表明BACE2蛋白實際上是一種疏水性質的蛋白質。
正值越高,代表疏水性越強;負值越高,代表其親水性也越強。
2.5 BACE2蛋白的跨膜結構
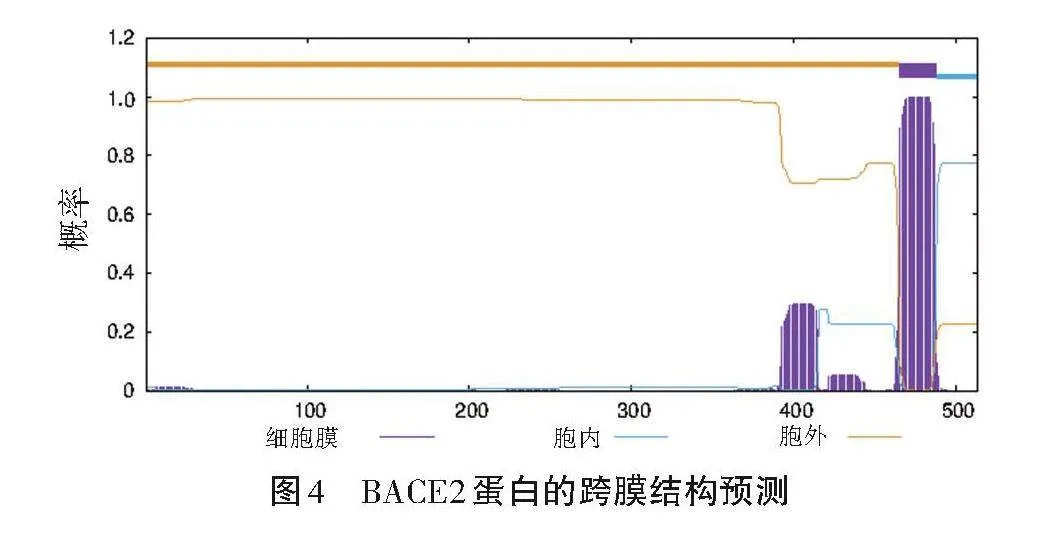
運用在線技術平臺對BACE2蛋白的跨膜結構進行研究,結果如圖4所示。圖4橫軸呈現了氨基酸在蛋白質中的位置,而縱軸揭示了可能存在的膜上跨越結構。與現存生物材料進行比較,認為該理論模型可精確地解釋BACE2蛋白的膜層跨接結構的形成。結果表明,呂梁黑豬體內的BACE2蛋白中有12個區域具備穿透細胞膜的能力。基于這些數據,可以假設該蛋白質具有3個膜間結構。
2.6 BACE2蛋白序列比對與進化
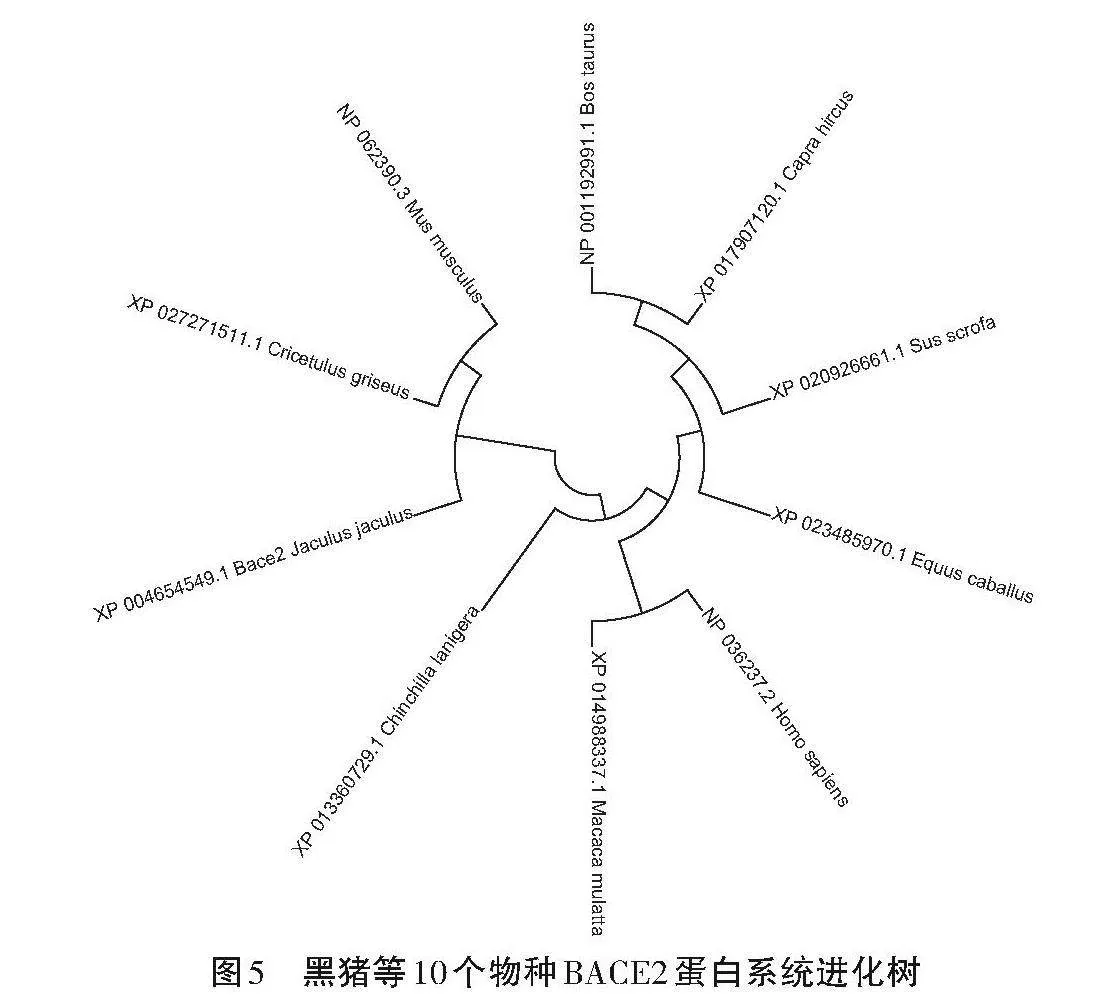
使用DNA MAN軟件將呂梁黑豬BACE2蛋白的氨基酸序列與牛(NP 001192991.1)、山羊(XP 017907120.1)、馬(XP 023485970.1)、智人(NP 036237.2)、獼猴(XP 014988337.1)、長尾龍貓(XP 013360729.1)、小埃及跳鼠(XP 004654549.1)、中國倉鼠(XP 027271511.1)和家鼠(NP 062390.3)的BACE2序列進行比對,繪制系統進化樹,如圖5所示。呂梁黑豬與山羊和牛的親緣關系較近,與家鼠的較遠。不同物種的BACE2蛋白序列之間存在一定程度的相似性。
2.7 BACE2蛋白的二級、三級結構預測
蛋白質二級結構作為一級結構和三級結構的紐帶,對其預測可為三級結構和功能提供大量信息,如設計蛋白質突變體或確定蛋白質空間結構及功能。使用SWISS-MODEL軟件進行BACE2蛋白的二級、三級結構模擬和預測,結果如圖6所示。
由圖6可知,BACE2蛋白的二級結構主要表現為不規則的卷曲形態,包含252個氨基酸,占總氨基酸的49.03%。α-螺旋結構包含95種氨基酸,占總氨基酸的18.48%。擴展部分包含140種氨基酸,占總氨基酸的27.24%;除此之外,這些也是由不包含肽鍵和不規則連接點的短鏈組成的。在β-轉角中,存在27種氨基酸,占總氨基酸的5.25%;還存在其他蛋白質中的一些小片段,這些片段呈現了一定的相似性,這些特征表明其是一個具有巨大開發潛力的動物資源庫。
在對呂梁黑豬BACE2蛋白的結構特性進行深入研究的過程中,發現其三級結構主要是基于二級結構來構建的,并且通過多肽鏈進行進一步的盤繞和折疊,最終形成了一種復雜的三維形態(圖6C)。在這些元素中,存在許多具有不同構型空間的非共價鍵連接模式的二級結構單元。這些次級結構單元具有多樣的形狀,包括α-螺旋、β-折疊、β-轉角,以及各種不規則的卷曲感。β-折疊與其他3個類別之間有一定的聯系。在次級鍵的驅動下,這些結構在側鏈基團間的相互作用中共同轉變為蛋白質的三級結構。由于氨基酸殘基間的相互作用力不同,以及受分子內氫和氧形成共價鍵等各種因素影響,其空間結構呈現明顯的異質性。分析結果表明,該蛋白達到了三級結構的標準,但仍然呈現出不規則的卷縮形態,這進一步突顯了其結構成分的多樣性和高度的復雜性。
2.8 BACE2蛋白亞細胞定位
通過PSORT Ⅱ在線軟件分析發現,呂梁黑豬的BACE2蛋白主要分布于內質網(55.6%),同時還分布于高爾基(22.2%)、細胞外(11.1%)、細胞壁和質膜,占比11.1%。
2.9 BACE2蛋白互作網絡
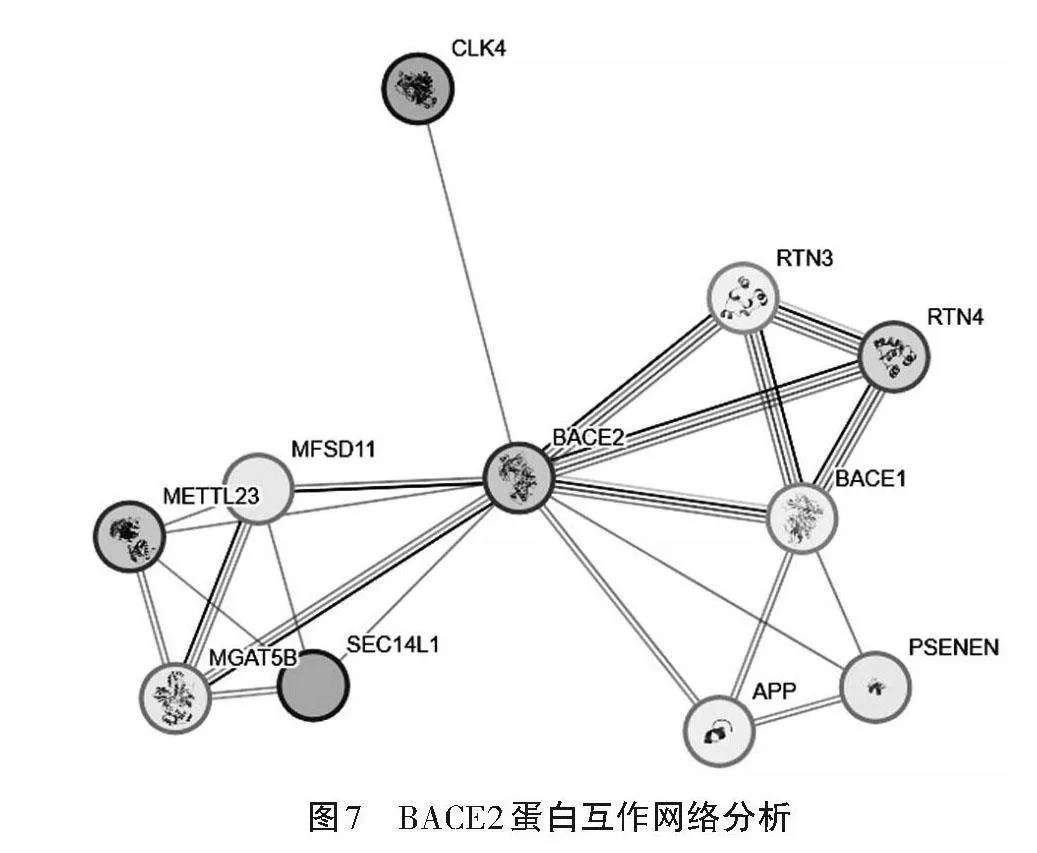
利用在線軟件STRING構建BACE2蛋白互作網絡,結果如圖7所示,BACE2蛋白與CLK4、RTN3、RTN4、AKT3、MFSD11、BACE1、METTL23、PSENEN、MGAT5B和SEC14L1蛋白存在相互作用。網絡互作節點11個,邊數22個,平均局部聚類系數0.897。
2.10 BACE2基因在不同組織中的表達量
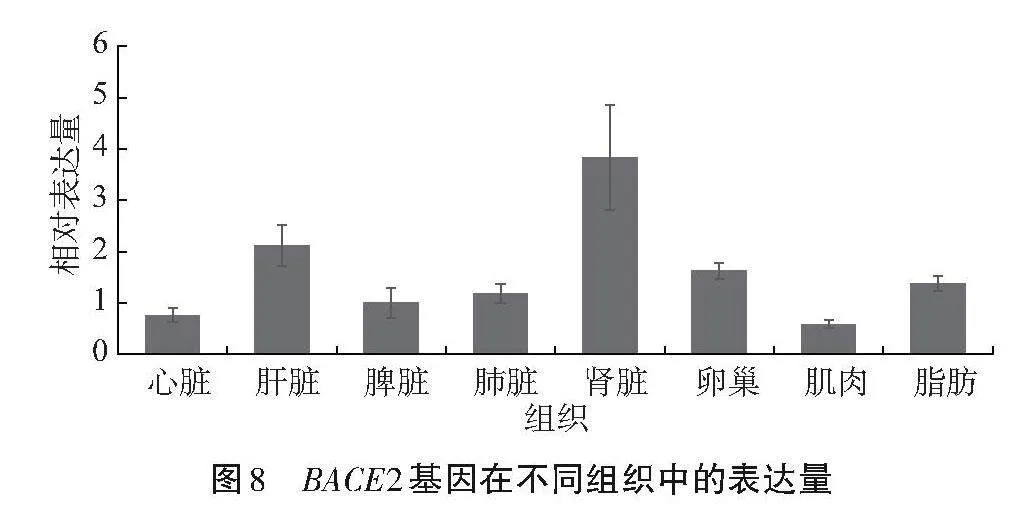
由圖8可知,BACE2基因在呂梁黑豬心臟、肝臟、脾臟、肺臟、腎臟、肌肉、卵巢和脂肪組織中均有表達,其中在腎臟中的表達量最高,其次是肝臟,肌肉組織中的表達量最低。
3 結論與討論
BACE2與BACE1具有同源性,在APP代謝途徑中扮演類似的角色,可以水解APP。BACE2的重要功能之一是調節Aβ的生成,而Aβ是AD中神經纖維斑塊的主要成分[6]。目前,已有研究證明BACE2在其他生理和病理過程中同樣發揮重要作用。Sun等[7]和虞永峰[8]在非小細胞肺癌細胞中敲減BACE2基因,發現其能明顯影響癌細胞的遷移、侵襲能力以及骨轉移。劉希等[9]研究發現,引入人源性BACE2基因可以降低小鼠腦組織內APP的表達,減少海馬區Aβ淀粉樣斑塊沉積,明顯改善了小鼠的學習和記憶能力。鑒于BACE2基因在動物生理和病理過程中的重要作用[10-11] ,本研究在呂梁黑豬中開展了相關研究,成功克隆了黑豬BACE2基因的CDS區序列,全長1 545"bp,編碼514個氨基酸。BACE2蛋白是堿性的,缺少信號肽,表明其合成后會直接在核糖體中發揮作用,不需要經歷分泌過程,因此被歸類為非分泌型蛋白。這類蛋白質不僅展現了出色的疏水性和穩定性,而且還表現出明顯的跨膜作用。此外,從蛋白質二級結構看,呂梁黑豬BACE2蛋白以無規則卷曲為主,并伴有α-螺旋及片狀結構分散分布。三級結構預測結果對二級結構成分預測精度有進一步驗證的作用,兩者結論高度吻合。Maloney等[12]和Sáez-valero等[13]研究表明,BACE2蛋白在同系同物種的進化過程中發揮了至關重要的作用。本研究將黑豬BACE2基因序列與牛、山羊等9個物種進行比對,結果表明,呂梁黑豬與山羊和牛的親緣關系較近,與家鼠的較遠,表明哺乳動物的BACE2基因序列相似性較高。
Sáez-valero等[13]分析表明,BACE2"mRNA在人體外周組織中表達水平較低,而在結腸、腎臟、胰腺、胎盤、前列腺、胃和氣管中表達水平較高;成人和胎兒的全腦和大多數成人腦亞區表達很低或檢測不到,成年大鼠腦原位雜交顯示,BACE2"mRNA在大部分腦區以極低水平表達。本研究檢測BACE2基因在呂梁黑豬心臟、肝臟、脾臟、肺臟、腎臟、肌肉、卵巢、脂肪組織中均有表達,以在腎臟中的表達量最高,其次是肝臟,肌肉組織中的表達量最低。結合亞細胞定位結果,BACE2蛋白主要分布于內質網上,亞細胞定位結果可能與其作為酶的功能有關。
本研究成功地克隆了黑豬BACE2基因,并對該基因編碼的BACE2蛋白使用生物信息學軟件進行詳細的預測與分析,包括其是否含有信號肽、親水和疏水屬性、膜之間的結構特征,以及蛋白質的二級和三級結構等。結果表明,黑豬BACE2基因的CDS序列長度1 545"bp,編碼了514種氨基酸;BACE2蛋白是堿性的,缺少信號肽,展現了出色的疏水性和穩定性,而且還表現出了明顯的跨膜作用;其二級和三級結構高度吻合,主要存在與內質網,與多種蛋白具有互作作用;在不同的哺乳物種之間表現出高度的同源性。本研究通過檢測不同組織的BACE2基因表達量,發現其在腎臟中的表達量最高,肌肉中的表達量最低。本研究為BACE2蛋白未知生物學功能的研究提供一個新的角度,為了解動物細胞侵襲機制及色素沉著預防與控制奠定基礎,為相關方面的研究工作提供參考。未來將綜合運用生物信息學研究手段,對黑豬BACE2基因表達機制進行進一步的探索。
參考文獻
[1] 周亭,呂月琴. 憨態敦厚的豬[J]. 少兒美術,2023(11):24-27.
[2] 崔海紅. 丹麥養豬業生態建設對中國養豬業可持續發展的啟示[J]. 世界農業,2020(6):98-103.
[3] WANG Z,XU Q,CAI F,et al. BACE2,a conditional β-secretase,contributes to Alzheimer’s disease pathogenesis[J]. JCI insight,2019,4(1):e123431.
[4] 謝建,劉勝春. BACE2在人體胰腺和胰島素瘤中的表達[J]. 重慶醫科大學學報,2012,37(1):6-9.
[5] 王會志. BACE2激活NF-κB通路促進膠質瘤惡性進展的機制研究[D]. 濟南:山東大學,2022.
[6] 王瑞山,張云武,張弦,等. HIF-1調控小鼠bace1基因的轉錄表達[J]. 廈門大學學報(自然科學版),2007,46(4):564-568.
[7] SUN X L,HE G Q,SONG W H. BACE2,as a novel APP θ-secretase,is not responsible for the pathogenesis of Alzheimer’s disease in Down syndrome[J]. The FASEB journal,2006,20(9):1369-1376.
[8] 虞永峰. BACE2在非小細胞肺癌骨轉移中的作用及機制[D]. 上海:上海交通大學,2018.
[9] 劉希,鄭娜,盧宏. 人源性BACE2對阿爾茨海默病小鼠海馬區淀粉樣斑塊形成及空間學習記憶能力的影響[J]. 鄭州大學學報(醫學版),2021,56(2):195-199.
[10] QIU K X,LIANG W P,WANG S,et al. BACE2"degradation is mediated by both the proteasome and lysosome pathways[J]. BMC molecular and cell biology,2020,21(1):13.
[11] DíAZ-CATALáN D,ALCARRAZ-VIZáN G,CASTA?O C,et al. BACE2"suppression in mice aggravates the adverse metabolic consequences of an obesogenic diet[J]. Molecular metabolism,2021,53:101251.
[12] MALONEY B,GE Y W,GREIG N H,et al. Characterization of the human beta-secretase 2 (BACE2) 5’-flanking region:identification of a 268-bp region as the basal BACE2 promoter[J]. Journal of molecular neuroscience,2006,29(1):81-99.
[13] SáEZ-VALERO J,PéREZ-GONZáLEZ R. BACE2 beyond β-processing of APP,its neuroprotective role in cerebrovascular endothelium[J]. Journal of neurochemistry,2023,166(6):887-890.
(責任編輯:胡立萍)
