歐前胡素調節ThPOK表達對胃癌細胞惡性生物學行為的影響
2025-02-11 00:00:00陳蘭夏伶俐陳穎張剛文峰
中國藥房 2025年2期
中圖分類號 R965;R735.2 文獻標志碼 A 文章編號 1001-0408(2025)02-0191-06
DOI 10.6039/j.issn.1001-0408.2025.02.10
摘要 目的 探究歐前胡素(IMP-SD)調節含鋅指和BTB結構域7B(ThPOK)表達對胃癌(GC)細胞惡性生物學行為的影響。方法 取人GC細胞MKN-7,分為對照組(不給藥),IMP-SD低、中、高濃度組(分別給予40、80、160 μmol/L 的IMP-SD),si-ThPOK和si-NC組[先給予160 μmol/L 的IMP-SD,再分別轉染ThPOK小分子干擾RNA(si-ThPOK)及其陰性對照(si-NC)])。經相應處理后,檢測各組細胞的克隆形成、遷移、侵襲能力及凋亡情況,自然殺傷(NK)細胞的殺傷作用,T細胞的分布情況,以及ThPOK、程序性死亡受體1(PD-1)及其配體(PD-L1)的表達情況。結果 與對照組比較,IMP-SD各濃度組的細胞克隆數、遷移細胞數、侵襲細胞數和PD-1、PD-L1 蛋白的表達均顯著降低或下調,細胞凋亡率、NK細胞的殺傷活性、CD4+ T細胞比例、CD4+ T細胞比例與CD8+ T細胞比例的比值(CD4+ T/CD8+ T)、ThPOK蛋白的表達均顯著升高或上調,且有濃度依賴性(P<0.05);與IMP-SD高濃度組和si-NC組比較,si-ThPOK組的細胞克隆數、遷移細胞數、侵襲細胞數和PD-L1、PD-1 蛋白的表達均顯著升高或上調,細胞凋亡率、NK細胞的殺傷活性、CD4+ T細胞比例、CD4+ T/CD8+ T、ThPOK蛋白的表達均顯著降低或下調(P<0.05)。結論 IMP-SD可能通過促進ThPOK蛋白的表達,從而減弱GC細胞的克隆形成、遷移、侵襲能力,促進其凋亡,抑制其免疫逃逸。
關鍵詞 歐前胡素;ThPOK;胃癌細胞;免疫逃逸;惡性生物學行為
近年來,盡管胃癌(gastric cancer,GC)的發病率和相關死亡率持續降低,但2020 年全球新發病例近109萬、死亡病例約77 萬,GC仍是全球第三大癌癥死亡原因[1]。GC患者確診時常處于晚期,主要通過手術切除治療,但可因轉移、復發而導致患者總體預后不佳。研究指出,轉移、免疫逃逸和對治療的抵抗與GC患者的不良預后密切相關[2]。腫瘤細胞的免疫逃逸是其通過修飾自身表面抗原、募集抑制性免疫細胞/分子及改變腫瘤組織周圍微環境等途徑來逃避機體免疫系統的監控、識別及攻擊,從而繼續分裂生長的過程[3]。因此,了解GC細胞的免疫逃逸機制并開發針對該機制的藥物,對于GC的臨床治療具有重要的意義。
歐前胡素(imperatorin,IMP-SD)能抑制腫瘤細胞的活力,使細胞周期停滯,抑制喉癌、橫紋肌肉瘤、骨肉瘤和GC等腫瘤細胞的增殖、遷移,并促進其凋亡[4―7]。含鋅指和BTB結構域7B(zinc finger and BTB domain 7B,ThPOK)是POZ 結構域Krüppel 樣鋅指(POZ domainKrüppel-like zinc finger,POK)家族成員,對成熟輔助性T細胞的穩態和分化至關重要,可控制CD4+ T細胞的分化并調節其功能,在直腸癌、胸腺癌、GC細胞的免疫逃逸過程中具有重要作用[8―9]。ThPOK轉錄沉默元件可抑制ThPOK 的表達,從而啟動腸黏膜中CD4+腸上皮淋巴細胞的分化[10]。可見,調控ThPOK的表達是調節T細胞分化的重要機制。此外,過表達的ThPOK可減弱GC細胞的活力,增加T 細胞的分化,并可通過誘導精子尾部富含PG的重復序列1 使胞外信號調節激酶信號通路失活,從而抑制GC細胞的免疫逃逸[11]。然而,IMP-SD 能否通過調節ThPOK 的表達來影響GC細胞免疫逃逸等惡性生物學行為尚不明確。基于此,本研究擬探討IMPSD對GC細胞多種惡性生物學行為的影響,并探討相應作用的發揮是否與調控ThPOK的表達有關,以期為GC有效治療藥物的研發及應用提供參考。
1 材料
1.1 主要儀器
本研究所有主要儀器包括NanoFCM型流式細胞儀(廈門福流生物科技有限公司),ChemiDocTM型化學發光成像儀、CriterionTM型垂直電泳儀(美國Bio-Rad 公司),CX40M 型顯微鏡[舜宇光學科技(集團)有限公司],FormaTM Steri-Cult型CO2細胞培養箱、Multiskan SkyHigh型生物酶標儀(美國Thermo Fisher Scientific公司)等。
1.2 主要藥品與試劑
IMP-SD 對照品( 批號M71520230321,純度≥95.0%)購自美國MSD公司;GC細胞專用培養基RPMI-1640、胎牛血清、青- 鏈霉素雙抗溶液(貨號分別為PM150110、164210-50、PB180120)均購自武漢普諾賽生命科技有限公司;鼠源ThPOK單克隆抗體、鼠源程序性死亡受體1(programmed death-1,PD-1)單克隆抗體、兔源程序性死亡受體配體1(programmed death-ligand 1,PD-L1)單克隆抗體、兔源甘油醛-3-磷酸脫氫酶(GAPDH)多克隆抗體(貨號分別為GTX83353、GTX20256、GTX635975、GTX100118)均購自美國GenTex 公司;CD4+、CD8+熒光抗體和山羊抗兔、山羊抗鼠IgG(Hamp;L)二抗(批號分別為ab95591、ab210372、ab6702、ab6708)均購自英國Abcam 公司;人CD4+ T、CD8+ T細胞檢測試劑盒(貨號分別為TBDCH100154、TBDCH100140)均購自上海研瑾生物科技有限公司;人自然殺傷(naturalkill,NK)細胞檢測試劑盒(貨號qy-1953R)購自齊一生物科技(上海)有限公司;CCK-8 試劑(批號96992)購自德國Merck 公司;Annexin Ⅴ-FITC/PI 細胞凋亡檢測試劑盒(批號MA0220)購自大連美侖生物技術有限公司;基質膠(批號AC-M082704)購自百普賽斯生物科技有限公司;Lipofectamine 2000 轉染試劑(批號11668019)購自美國Thermo Fisher Scientific 公司;ThPOK 小分子干擾RNA(si-ThPOK)及其陰性對照(si-NC)(批號分別為siB112503026314-1-5、siB108436814-123-1-5)均購自廣州市銳博生物科技有限公司。
1.3 細胞
人GC細胞MKN-7(貨號CL-0574)購自武漢普諾賽生命科技有限公司;人外周血CD8+ T細胞(貨號PRI-H-00121)購自上海中喬新舟生物科技有限公司;人外周血CD4+ T 細胞(貨號ABC-TC3993)購自北京百奧創新科技有限公司;人NK細胞(貨號CP-H168)購自武漢益普生物科技有限公司。
2 方法
2.1 IMP-SD干預濃度篩選
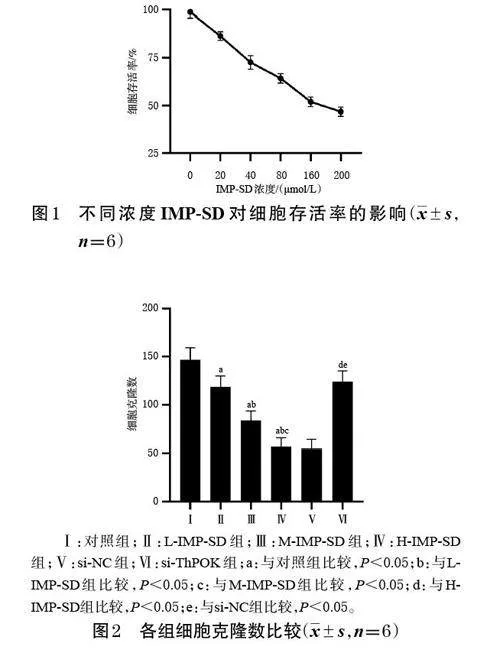
采用CCK-8 法檢測、篩選。取對數生長期的MKN-7 細胞,按每孔3 000 個接種于96 孔板中,待細胞貼壁生長后,分別加入含不同濃度IMP-SD[0(對照)、20、40、80、160、200 μmol/L[12]]的含胎牛血清、青-鏈霉素雙抗的RPMI-1640 培養基(以下稱“完全培養基”),并設置不含藥物、不含細胞的空白對照(每濃度設置6 個復孔)。培養48 h 后,每孔加入CCK-8 試劑10 μL,孵育2 h,使用酶標儀在450 nm波長處檢測各孔的吸光度,并計算細胞存活率[細胞存活率=(實驗孔吸光度值-空白對照孔吸光度值)/(對照孔吸光度值-空白對照吸光度值)×100%]和半數抑制濃度(IC50),以確定后續實驗IMP-SD的低、中、高濃度。
2.2 MKN-7 細胞分組與干預
取對數生長期的MKN-7 細胞,隨機分為對照組,IMP-SD 低、中、高濃度組(L-IMP-SD 組、M-IMP-SD 組、H-IMP-SD組),si-NC組和si-ThPOK組,每組設置6 個復孔。對照組細胞加入完全培養基;IMP-SD 各濃度組細胞分別加入含相應濃度IMP-SD(根據“2.1”項下結果設置)的完全培養基;si-NC、si-ThPOK 組細胞經含高濃度IMP-SD 的完全培養基處理后,分別轉染si-NC、si-ThPOK(以si-NC組的ThPOK 表達較H-IMP-SD 組基本不變且si-ThPOK組的ThPOK表達較H-IMP-SD組顯著下降為轉染成功)。
2.3 MKN-7 細胞克隆形成能力檢測
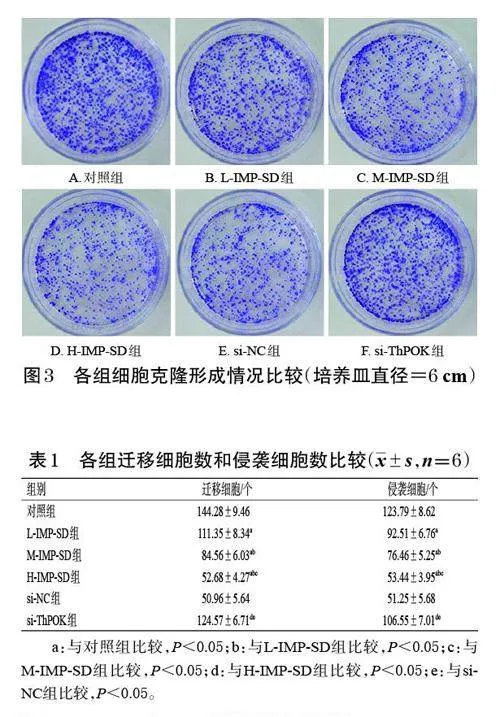
采用克隆形成實驗檢測。取對數生長期的MKN-7細胞,按“2.2”項下方法分組、處理。培養10 d(每3 d 更換培養基并重新給藥)后,棄去培養基,細胞以4% 多聚甲醛溶液固定30 min,再以0.1% 結晶紫染液染色30min;移除染液,細胞用磷酸鹽緩沖液(PBS)漂洗,風干后,使用顯微鏡觀察并記錄各組的細胞克隆數(>10 個細胞即算1個克隆)。
2.4 MKN-7 細胞遷移和侵襲能力檢測
均采用Transwell 實驗檢測。取對數生長期的MKN-7 細胞,經消化后以PBS 漂洗,并以不含血清的RPMI-1640 培養基重懸,制成1×105個/mL的細胞懸液。取上述懸液100 μL,加至Transwell 小室上層;取完全培養基600 μL,置于小室下層。上層細胞按“2.2”項下方法分組、處理。培養24 h 后,收集穿膜細胞,以4% 多聚甲醛溶液固定30 min,再以0.1%結晶紫染液染色10min,自然風干后,使用顯微鏡觀察并記錄各組的遷移細胞數。以基質膠處理小室上層,其余操作同前,使用顯微鏡觀察并記錄各組的侵襲細胞數。
2.5 MKN-7 細胞凋亡檢測
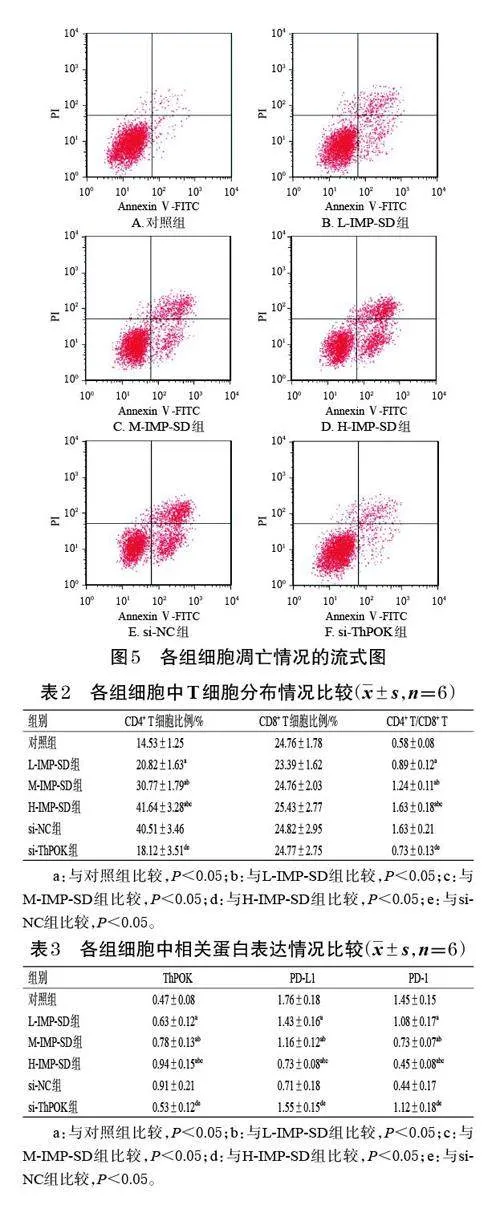
采用流式細胞術檢測。取對數生長期的MKN-7 細胞,按“2.2”項下方法分組、處理。培養24 h 后,收集各組細胞,以PBS漂洗后,加入binding buffer 200 μL重懸;加入Annexin Ⅴ-FITC 染液5 μL,于室溫下孵育10 min;加入PI 染液10 μL,于避光條件下孵育10 min,使用流式細胞儀檢測各組細胞凋亡情況。
2.6 NK細胞對MKN-7細胞的殺傷作用檢測
采用乳酸脫氫酶釋放法檢測。取對數生長期的MKN-7 細胞,按“2.2”項下方法分組、處理。培養24 h后,收集各組細胞,與NK細胞(各組MKN-7 細胞與NK細胞的數量比均為1∶10)混合,于37 ℃下共同培養6 h;收集細胞,加入乳酸脫氫酶50 μL,于室溫下孵育30min,使用酶標儀于450 nm波長處檢測其光密度(OD450)值,再按相應試劑盒說明書方法計算NK細胞殺傷活性。
2.7 MKN-7 細胞的T細胞分布情況檢測
采用流式細胞術檢測。取對數生長期的MKN-7 細胞,按“2.2”項下方法分組、處理。培養24 h 后,收集各組細胞,分別與CD4+ T、CD8+ T 細胞(各組MKN-7 細胞與T細胞的數量比均為1∶10)混合,于37 ℃下共同培養6 h;收集細胞,分別加入CD4+、CD8+熒光抗體(稀釋比例均為1∶500),于室溫下避光孵育20 min;收集細胞,以PBS重懸后,使用流式細胞儀分析CD4+ T細胞、CD8+ T細胞比例及兩者比值(CD4+ T/CD8+ T)。
2.8 MKN-7 細胞中ThPOK、PD-1、PD-L1 蛋白表達檢測
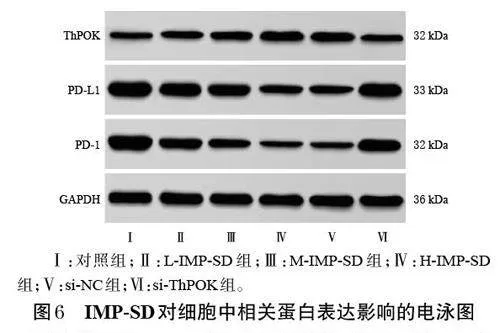
采用Western blot 法檢測。取對數生長期的MKN-7細胞,按“2.2”項下方法分組、處理。培養24 h 后,收集各組細胞,提取蛋白。蛋白經定量、變性后進行電泳分離、轉膜、封閉;洗膜后,加入ThPOK、PD-1、PD-L1、GAPDH一抗(稀釋比例分別為1∶500、1∶1 000、1∶1 000、1∶5 000),于4 ℃下孵育過夜;洗膜后,加入相應二抗(稀釋比例均為1∶5 000),于室溫下孵育1 h,使用化學發光試劑顯影后,于化學發光成像儀下成像。使用Image J 軟件,以GAPDH為內參,量化各目的蛋白的表達情況。
2.9 統計學方法
采用SPSS 26.00 軟件對數據進行統計分析。數據以x±s 表示,多組間比較采用單因素方差分析,進一步兩兩比較采用SNK-q 檢驗。檢驗水準α=0.05。
3 結果
3.1 IMP-SD干預濃度的篩選結果
經0、20、40、80、160、200 μmol/L IMP-SD干預后,細胞存活率隨其濃度的增加而逐漸降低,IC50 為168.42μmol/L,遂確定以40、80、160 μmol/L 作為后續實驗的低、中、高濃度。結果見圖1。
3.2 IMP-SD對MKN-7 細胞克隆形成能力的影響
與對照組比較,L-IMP-SD、M-IMP-SD、H-IMP-SD組的細胞克隆數均顯著降低,且有濃度依賴性(P<0.05);與H-IMP-SD 組、si-NC組比較,si-ThPOK 組的細胞克隆數顯著升高(P<0.05)。結果見圖2、圖3。
3.3 IMP-SD對MKN-7 細胞遷移和侵襲的影響
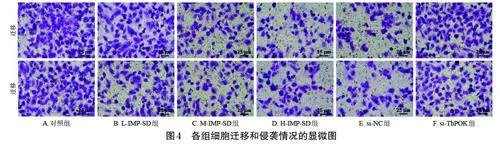
與對照組比較,L-IMP-SD、M-IMP-SD、H-IMP-SD組的遷移細胞數、侵襲細胞數均顯著減少,且有濃度依賴性(P<0.05);與H-IMP-SD 組、si-NC 組比較,si-ThPOK 組的遷移細胞數、侵襲細胞數均顯著升高(P<0.05)。結果見表1、圖4。
3.4 IMP-SD對MKN-7 細胞凋亡的影響
與對照組[(2.96±0.28)%]比較,L-IMP-SD、M-IMPSD、H-IMP-SD 組細胞的凋亡率[(18.47±1.29)% 、(31.85±2.57)%、(49.55±3.64)%]均顯著升高,且有濃度依賴性(P<0.05);與H-IMP-SD組、si-NC組[(47.26±3.78)% ] 比較,si-ThPOK 組細胞的凋亡率[(12.78±3.19)%]顯著降低(P<0.05)。結果見圖5。
3.5 IMP-SD對NK細胞殺傷活性的影響
與對照組[(8.50±0.43)%]比較,L-IMP-SD、M-IMPSD、H-IMP-SD 組細胞的NK 細胞殺傷活性[(16.28±0.54)%、(28.77±1.08)%、(43.39±1.24)%]均顯著升高,且有濃度依賴性(P<0.05);與H-IMP-SD 組、si-NC 組[(42.67±1.36)%]比較,si-ThPOK 組細胞的NK 細胞殺傷活性[(14.95±1.07)%]顯著降低(P<0.05)。
3.6 IMP-SD對MKN-7 細胞中T細胞分布情況的影響
與對照組比較,L-IMP-SD、M-IMP-SD、H-IMP-SD組的CD4+ T細胞比例和CD4+ T/CD8+ T均顯著升高,且有濃度依賴性(P<0.05);與H-IMP-SD 組、si-NC 組比較,si-ThPOK組的CD4+ T細胞比例和CD4+ T/CD8+ T均顯著降低(P<0.05);而各組的CD8+ T細胞比例比較,差異均無統計學意義(P>0.05)。結果見表2。
3.7 IMP-SD對MKN-7 細胞中相關蛋白表達的影響
與對照組比較,L-IMP-SD、M-IMP-SD、H-IMP-SD組細胞中ThPOK蛋白的表達顯著上調,PD-L1、PD-1 蛋白的表達均顯著下調,且有濃度依賴性(P<0.05);與HIMP-SD組、si-NC組比較,si-ThPOK組細胞中ThPOK蛋白的表達顯著下調,PD-L1、PD-1 蛋白的表達均顯著上調(P<0.05)。結果見表3、圖6。
4 討論
GC是全球第四大常見癌癥、第三大癌癥死亡原因,在全球范圍內造成了嚴重的健康負擔[1]。由于GC早期癥狀不明顯、定期篩查率低,大多數患者在確診時已處于晚期,手術切除和圍手術期化療的效果有限,加之轉移率、復發率較高,導致GC患者的總體預后不佳[1]。因此,闡釋GC發生發展的具體機制并挖掘新的治療策略是有必要的。
IMP-SD 是香豆素類天然化合物,具有抑制腫瘤細胞增殖、誘導細胞凋亡的活性,并可增強多種抗腫瘤藥物的細胞毒性[4―6]。IMP-SD 能拮抗三磷酸腺苷結合轉運蛋白G 家族成員2(ATP-binding cassette subfamily Gmember 2,ABCG2)的活性,并可濃度依賴性地逆轉ABCG2 蛋白介導的多重耐藥性,以提高治療藥物用于ABCG2 高表達惡性腫瘤患者的有效性[4]。以往研究表明,IMP-SD 對人橫紋肌肉瘤和喉癌細胞表現出明顯的生長抑制作用,這種作用主要是通過阻滯與特定基因表達變化相關的細胞周期進程而實現的[5]。研究指出,IMP-SD可通過第10 號染色體上缺失的磷酸酶及張力蛋白同源物(phosphatase and tensin homolog deleted onchromosome ten,PTEN)/磷脂酰肌醇3 激酶/蛋白激酶B/哺乳動物雷帕霉素靶蛋白/p21 信號通路靶向上調人骨肉瘤細胞中的PTEN,使細胞周期停滯,從而抑制骨肉瘤細胞的增殖、遷移[6]。另有研究指出,IMP-SD 可下調環磷腺苷應答元件結合蛋白1 的表達,從而阻斷食管癌細胞的侵襲,進而減弱成纖維細胞旁分泌對腫瘤血管生成和轉移的影響[13]。還有研究指出,IMP-SD 可濃度依賴性地抑制人GC細胞的生長,誘導DNA片段化,使細胞周期停滯在G1 期并促進其凋亡[7]。以上研究均證實,IMP-SD 具有抗腫瘤活性。本研究也得出了相似的結果,即IMP-SD能抑制GC細胞的活力,可濃度依賴性地抑制GC細胞的克隆形成、遷移、侵襲,并誘導其凋亡。
ThPOK是轉錄因子POK家族成員,其Krüppel 樣鋅指結構域(負責與DNA結合)和調節性POZ/BTB結構域(介導與其他因子的相互作用)對成熟輔助性T 細胞的穩態和分化至關重要,與CD4+ T細胞的功能密切相關,也是介導CD4+ T 細胞轉化的關鍵結構[14]。過表達的ThPOK 可抑制GC細胞的活力,可促進與GC細胞共培養的T 細胞的活化,進而抑制GC 細胞的免疫逃逸[11]。此外,GC細胞可提高CD4+ T 細胞中ThPOK 的表達,促進CD4+ T 細胞的分化,使后者參與GC 細胞的免疫逃逸[15]。效應細胞在免疫系統的監視下優先清除免疫原性高的腫瘤細胞,而腫瘤細胞的有效清除需要CD4+ T和CD8+ T細胞[16―17]。其中,CD4+ T細胞對適應性免疫應答的發生和維持具有核心調控作用;CD8+ T細胞與腫瘤細胞的免疫反應有關,可識別腫瘤細胞的表面抗原并轉移、浸潤至細胞內部,從而殺死腫瘤細胞,釋放更多的抗原,進一步激活T 淋巴細胞介導的免疫反應;兩者比例失衡(如CD8+ T細胞耗竭而CD4+ T細胞生成)可促進免疫逃逸的發生[18]。NK細胞是一種先天免疫細胞,在先天免疫防御中發揮重要作用,是抵御惡性腫瘤的第一道防線,其活力降低與GC進展有關[2]。PD-1、PD-L1 對腫瘤細胞免疫逃逸至關重要,腫瘤細胞可通過高表達PDL1而與CD8+ T 細胞表面的PD-1 結合,抑制T 細胞的活化,從而協助腫瘤細胞逃避免疫監視,實現免疫逃逸[19]。本研究結果表明,經IMP-SD 處理后,MKN-7 細胞中ThPOK 蛋白的表達、NK細胞殺傷活性,CD4+ T 細胞比例、CD4+ T/CD8+ T 均顯著升高或上調,PD-L1、PD-1 蛋白的表達均顯著下調;而轉染si-ThPOK后,細胞上述指標的變化趨勢則剛好與H-IMP-SD 組相反。這表明,IMP-SD 能夠促進ThPOK 蛋白的表達,抑制GC細胞的免疫逃逸。
綜上所述,IMP-SD可能通過促進ThPOK蛋白的表達,從而減弱GC細胞的克隆形成、遷移、侵襲能力,促進其凋亡,抑制其免疫逃逸。但本研究僅基于細胞水平進行了相關探索,尚需通過體內實驗進一步對結果進行驗證。
參考文獻
[ 1 ] MA X X,JIA S Q,WANG G J,et al. TRIM28 promotes
the escape of gastric cancer cells from immune surveillance
by increasing PD-L1 abundance[J]. Signal Transduct
Target Ther,2023,8(1):246.
[ 2 ] MAO D L,ZHOU Z J,CHEN H X,et al. Pleckstrin-2 promotes
tumor immune escape from NK cells by activating
the MT1-MMP-MICA signaling axis in gastric cancer[J].
Cancer Lett,2023,572:216351.
[ 3 ] MILIOTIS C,MA Y L,KATOPODI X L,et al. Determinants
of gastric cancer immune escape identified from
non-coding immune-landscape quantitative trait loci[J].
Nat Commun,2024,15(1):4319.
[ 4 ] WU C P,MURAKAMI M,LI Y C,et al. Imperatorin
restores chemosensitivity of multidrug-resistant cancer
cells by antagonizing ABCG2-mediated drug transport[J].
Pharmaceuticals,2023,16(11):1595.
[ 5 ] GRABARSKA A,SKALICKA-WO?NIAK K,KIE?BUS
M,et al. Imperatorin as a promising chemotherapeutic
agent against human larynx cancer and rhabdomyosarcoma
cells[J]. Molecules,2020,25(9):2046.
[ 6 ] LV M C,XU Q X,ZHANG B,et al. Imperatorin induces
autophagy and G0/G1 phase arrest via PTEN-PI3K-AKTmTOR/
p21 signaling pathway in human osteosarcoma
cells in vitro and in vivo[J]. Cancer Cell Int,2021,21
(1):689.
[ 7 ] AM J U,GONG W J,SU Y,et al. Imperatorin shows selective
antitumor effects in SGC-7901 human gastric adenocarcinoma
cells by inducing apoptosis,cell cycle arrest
and targeting PI3K/Akt/m-TOR signalling pathway[J]. J
BUON,2017,22(6):1471-1476.
[ 8 ] GAO Y Y,ZAMISCH M,VACCHIO M,et al. NuRD
complex recruitment to ThPOK mediates CD4+ T cell lineage
differentiation[J]. Sci Immunol,2022,7(72):
eabn5917.
[ 9 ] MARIANI F,SENA,PEDRONI M,et al. Th inducing
POZ-Krüppel factor(ThPOK)is a key regulator of the immune
response since the early steps of colorectal carcinogenesis[
J]. PLoS One,2013,8(1):e54488.
[10] TEREKHOVA M,SWAIN A,BOHACOVA P,et al.
Single-cell atlas of healthy human blood unveils agerelated
loss of NKG2C+GZMB-CD8+ memory T cells and
accumulation of type 2 memory T cells[J]. Immunity,
2024,57(1):188-192.
[11] CHEN Y,JIANG L L,XIA L L,et al. ThPOK inhibits the
immune escape of gastric cancer cells by inducing STPG1
to inactivate the ERK pathway[J]. BMC Immunol,2022,
23(1):16.
[12] 景紅梅,樊海燕,黃小勇,等. 歐前胡素抑制人腎癌786-
O細胞增殖并促進其凋亡[J]. 山西醫科大學學報,2023,
54(10):1323-1330.
JING H M,FAN H Y,HUANG X Y,et al. Imperatorin inhibits
proliferation and promotes apoptosis of human renal
carcinoma cell line 786-O[J]. J Shanxi Med Univ,2023,
54(10):1323-1330.
[13] XU W W,HUANG Z H,LIAO L,et al. Direct targeting
of CREB1 with imperatorin inhibits TGF β2-ERK signaling
to suppress esophageal cancer metastasis[J]. Adv Sci
(Weinh),2020,7(16):2000925.
[14] NUNES-CABA?O H,RAMALHO-DOS-SANTOS A,
PIRES A R,et al. Human CD4 T cells from thymus and
cord blood are convertible into CD8 T cells by IL-4[J].
Front Immunol,2022,13:834033.
[15] 黃麗芳. 胃癌細胞對CD4+ T細胞ThPOK蛋白表達的影
響及其與CD4+ T細胞亞群分化的關系[D]. 南昌:南昌大
學,2017.
HUANG L F. Effect of gastric cancer cells on ThPOK protein
expression in CD4+ T cells and its relationship with
CD4+ T cell subsets differentiation[D]. Nanchang: Nanchang
University,2017.
[16] BASU J,REIS B S,PERI S,et al. Essential role of a
ThPOK autoregulatory loop in the maintenance of mature
CD4+ T cell identity and function[J]. Nat Immunol,2021,
22(8):969-982.
[17] ANDREWS L P, VIGNALI K M, SZYMCZAKWORKMAN
A L,et al. A Cre-driven allele-conditioning
line to interrogate CD4+ conventional T cells[J]. Immunity,
2021,54(10):2209-2217.e6.
[18] OUYANG Y,GU Y,ZHANG X H,et al. AMPKα2 promotes
tumor immune escape by inducing CD8+ T-cell exhaustion
and CD4+ Treg cell formation in liver hepatocellular
carcinoma[J]. BMC Cancer,2024,24(1):276.
[19] CHAKRABARTI J,KOH V,STEELE N,et al. Disruption
of Her2-induced PD-L1 inhibits tumor cell immune evasion
in patient-derived gastric cancer organoids[J]. Cancers,
2021,13(24):6158.
(收稿日期:2024-06-07 修回日期:2024-10-16)
(編輯:張元媛)
