內蒙古中東部草原克氏針茅和大針茅的葉性分析
2010-01-05 08:16:04賈美清高玉葆
天津師范大學學報(自然科學版) 2010年1期
賈美清,高玉葆,楊 勇
(1.南開大學生命科學學院,天津 300071;2.天津師范大學水環境與水資源重點實驗室,天津 300387;3.中國民航大學,天津 300300)
內蒙古中東部草原克氏針茅和大針茅的葉性分析
賈美清1,2,高玉葆1,楊 勇3
(1.南開大學生命科學學院,天津 300071;2.天津師范大學水環境與水資源重點實驗室,天津 300387;3.中國民航大學,天津 300300)
以內蒙古中東部草原分布的克氏針茅和大針茅為研究對象,對不同生境和相同生境下二者的葉性特征進行了分析研究.結果表明:在不同生境下,從典型草原西部邊緣到典型草原東部,貧瘠生境中克氏針茅和大針茅采取單位面積高氮含量的生存策略、相對肥沃生境中的克氏針茅和大針茅則采取單位面積低氮含量的生存策略,二者比葉面積的變化趨勢均與生境中前期獲得的實際降雨量變化趨勢一致,即生境中前期獲得的實際降雨量高則比葉面積相對較高;在相同生境下,克氏針茅采取高比葉面積、低單位面積氮含量的生存適應策略,大針茅采取低比葉面積、高單位面積氮含量的生存適應策略.
克氏針茅;大針茅;比葉面積;單位面積氮含量;單位重量氮含量
植物生態理論的主要目的之一是預測物種、群落乃至整個生態系統對競爭、氣候變化以及土地利用狀況的響應[1—2].水、熱分配不均勻和過度放牧使得內蒙古錫林郭勒盟草原退化日趨嚴重.認清草原植被尤其是草原建群種的生長及結構特性與環境因子的關系,對于草原恢復策略的制定具有重要的生態意義.
葉片是植物進化過程中對環境變化較敏感且可塑性較大的器官,在不同選擇壓力下已經形成各種適應類型,其結構特征最能體現環境因子對植物的影響或植物對環境的適應[2].植物的葉性狀可分為結構型性狀和功能型性狀.結構型性狀是植物葉片的生物化學結構特征,主要包括比葉面積(Nmass)、葉氮含量(Narea)和葉壽命等,在特定環境下保持相對穩定.葉氮含量的增加能夠提高植物水分利用效率[3],也可以提高葉片光合能力[4—5],比葉面積的減少可以防止植物體內水分散失.葉功能型性狀體現了葉片的生長代謝指標,主要包括光合速率、呼吸速率和氣孔導度等,隨時間和空間的變化而變化的程度相對較大.植物的這些葉性狀共同體現了植物為了獲得最大化碳收獲所采取的生存適應策略[6—7].克氏針茅和大針茅是內蒙古錫林郭勒盟重要的草場資源又是津京地區的重要綠色屏障,因此,具有不可忽視的經濟價值和生態意義.
克氏針茅和大針茅一直受到我國科技工作者的重視 ,二者在地理分布[8]、放牧管理[9—10]、種子和種子庫[11—12]、生長與繁殖[13—15]、遺傳分化[16—18]、形態[19—20]以及抗旱性[21—23]等方面的研究已經取得了顯著成果,在葉的結構[24]和功能(光合)[25]方面也有少量報道.然而,有關葉片結構型性狀對環境的適應性研究至今鮮見報道.由于葉片結構型性狀中的葉氮含量和比葉面積相對穩定,其變化可以反映物種在長期進化過程中對其生存環境的適應特征[26].因此,本研究以內蒙古中東部草原克氏針茅和大針茅葉片為研究對象,通過測定在不同生境和相同生境下大針茅和克氏針茅的比葉面積(SLA)和葉氮含量(包括Nmass和Narea)等葉性特征,分析和探討克氏針茅和大針茅在葉性方面對生境的適應特征,為制定草原的保護和恢復策略提供理論依據.
1 材料與方法
1.1 樣地的選取
2007年7月,在內蒙古錫林郭勒盟草原從西到東選取5個樣地.樣地的選取參考歷年氣象資料并結合實地考察的結果進行設置,體現了水分梯度和無人為干擾的原則.5個樣地從西到東依次分布于滿都拉圖鎮東部(滿都拉圖種群)、阿巴嘎旗新浩特東部(新浩特種群)、錫林浩特市西部(錫林浩特西種群)、錫林浩特市東部(錫林浩特東種群)及西烏珠穆沁旗巴彥烏拉東北部(巴彥烏拉種群).各樣地的位置及生境特點見表1和表2.
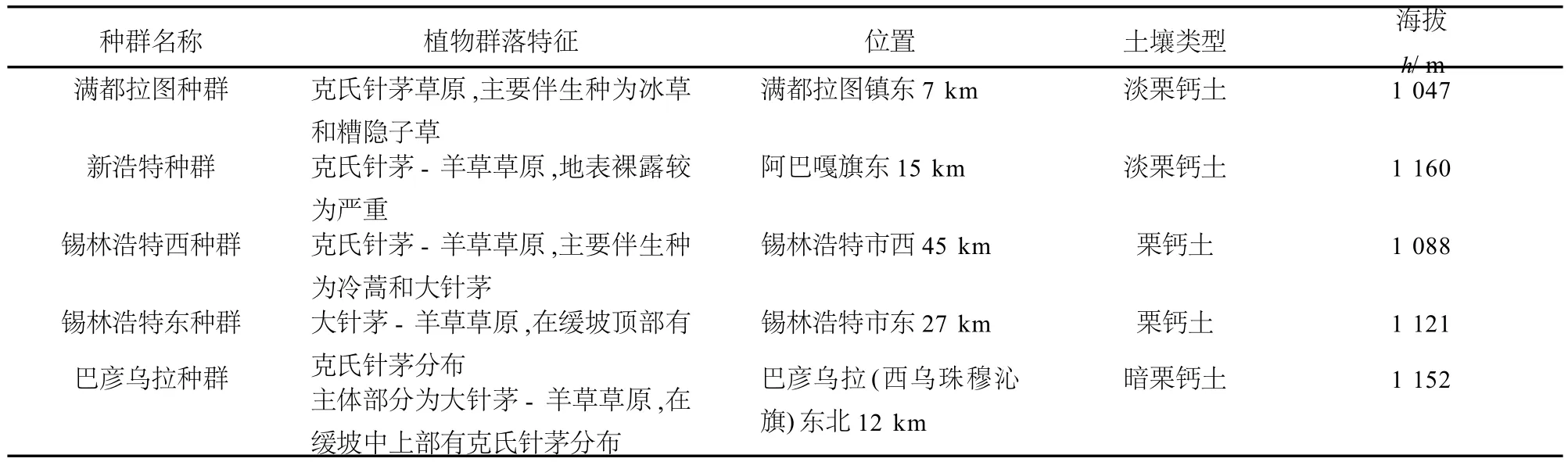
表1 種群的位置及地理特點Table1 Locationandhabitatcharactersoffivepopulations
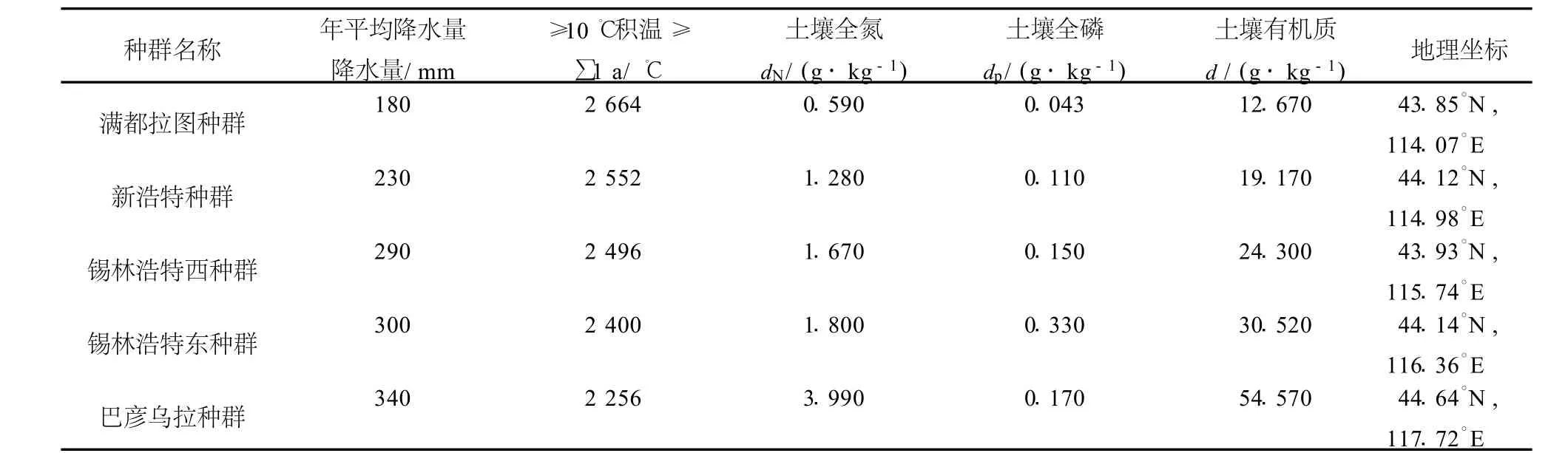
表2 種群的生境特點Table2 Habitafcharacferoffivepopulations
1.2 樣地的降水量測量
在設計5個樣地時,充分考慮了水分梯度對生境的影響,但是,在2007年7、8月2次采樣時,各樣地獲得的實際降雨量與樣地設置時的水分梯度仍不一致.2006年10月至2007年4月各樣地獲得的實際降雨量(降雨量-蒸發量)基本持平.2007年5月至6月各樣地獲得的實際降雨量從高到低的順序為:錫林浩特、新浩特、巴彥烏拉、滿都拉圖;5月至7月各樣地3個月內獲得的實際降雨量順序為:滿都拉圖、新浩特、錫林浩特、巴彥烏拉(見表3).平均溫度在各樣地基本持平.上述氣象資料均由錫林浩特市氣象局提供.
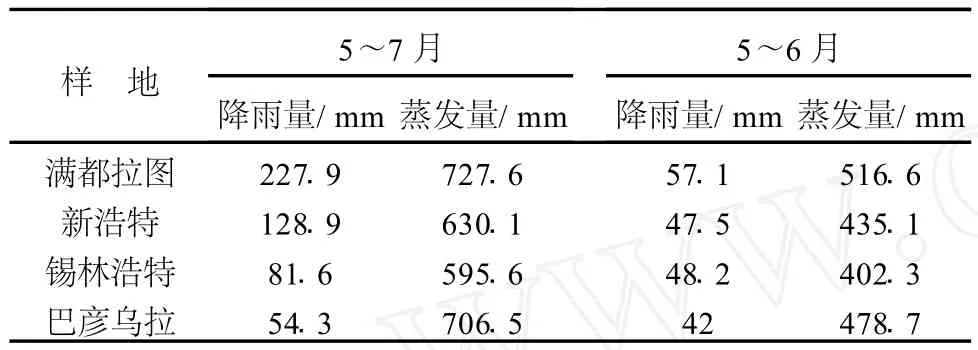
表3 2007年5~6月和5~7月各樣地的降雨量和蒸發量Table 3 Amount of precipitation and evaporation in every plot from May to July in 2007
2 實驗方法
2.1 取樣及葉性指標的測定
分別于2007年7月上旬和8月上旬,在每個樣地隨機選取50叢克氏針茅或大針茅.從每個株叢中隨機剪取一個沒有病蟲害的營養枝分蘗,同時剪取該分蘗從內到外的第二片葉片,置于兩片濕潤的濾紙之間,放入塑封袋內后封口,儲藏于黑暗的容器中帶回室內;剩余的營養枝分蘗放入信封中風干,帶回實驗室在60℃下烘干48 h[27]測定葉片氮含量.
葉氮含量(Nmass)的測定:采用半微量凱氏定氮法.
葉面積的測定:從塑封袋取出葉片,迅速用吸水紙吸去葉片表面的水分,測量葉片的長和寬,然后用透明膠帶將葉片分別粘在白紙上,復印,掃描,再用Pho toshop軟件進行分析[28—29].
葉干重(LDW)的測定:取下膠帶,烘干葉片48 h,用萬分之一的天平測量葉干重.
比葉面積、單位面積氮含量分別用下面的公式計算.
比葉面積(SLA)=葉面積/葉干重
單位面積氮含量(Narea)=單位重量氮含量(Nmass)/SLA
2.2 樣地土壤分析
2007年7月1至15日,對各樣地的土壤進行隨機取樣.各樣地分別取0~10,10~20,20~30 cm三層的土壤樣品,設10次重復,測定土壤有機質、全氮和全磷.土壤有機質測定采用重鉻酸鉀容量法——外加熱法,土壤全氮的測定采用半微量凱氏定氮法,土壤全磷的測定采用 HClO4—H2SO4法.
2.3 數據處理
利用 Excel和 SPSS(SPSS for Window s 13.0)對獲得的數據進行統計分析.在SPSS13.0軟件上,用單因素方差分析(ANOVA)中的LSD比較不同生境下,大針茅或克氏針茅的各葉性指標差異顯著性;利用配對樣本-t檢驗,對相同生境下大針茅和克氏針茅的各葉性指標進行差異顯著性檢驗.對大針茅和克氏針茅葉面積計算利用 Photoshop軟件完成.
3 結果分析
3.1 不同地理種群克氏針茅和大針茅的葉性分析
3.1.1 不同地理種群克氏針茅的葉性分析
7月份克氏針茅從典型草原西部的新浩特種群到典型草原較東部的錫林浩特東種群比葉面積(SLA)呈顯著升高趨勢;葉干重和單位面積氮含量呈顯著降低的趨勢;單位重量氮含量隨生境的變化沒有明顯的變化趨勢.8月份不同地理種群指標克氏針茅比葉面積從西部的滿都拉圖種群到東部的錫林浩特東種群先降低后升高,而葉干重呈相反的變化趨勢;除滿都拉圖種群外,其它3個種群單位面積氮含量和單位重量氮含量從西部的新浩特到東部的錫林浩特東均表現出降低的趨勢(表4).
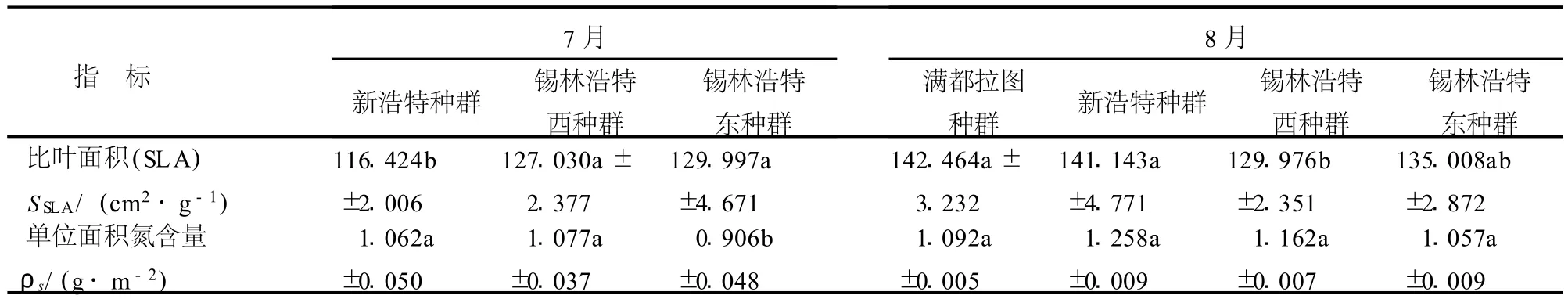
表4 不同地理種群克氏針茅葉性分析Table 4 Analysis of leaf traits within different S.k rylovii populations in July or August
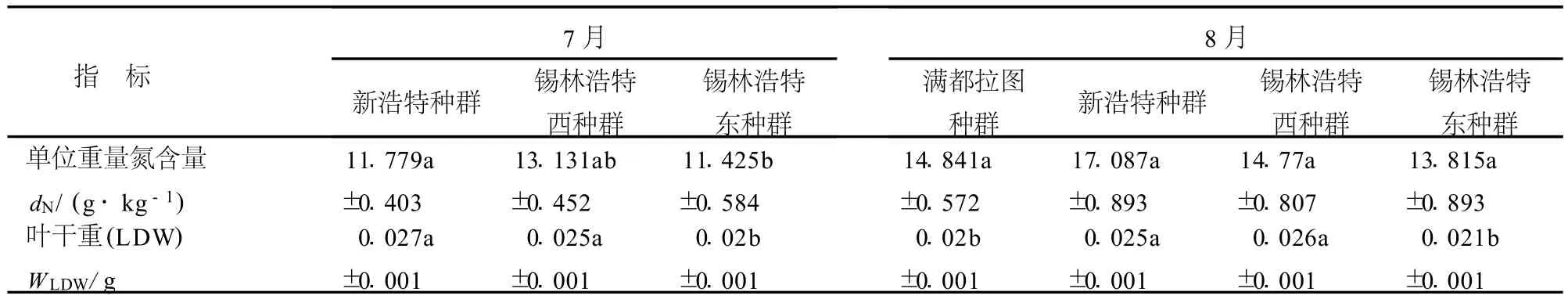
續表4
3.1.2 不同地理種群大針茅的葉性分析
從典型草原西部的錫林浩特西種群到東部的巴彥烏拉種群,7月份大針茅的單位重量氮含量、單位面積氮含量和比葉面積均呈顯著降低趨勢;葉干重在生境較優越的巴顏烏拉種群中最高,各種群間沒有明顯變化趨勢(表5).8月份典型草原東部的錫林浩特東種群比葉面積顯著高于其它2個種群.單位面積氮含量從西部的新浩特種群到東部錫林浩特東種群呈顯著降低的趨勢;單位重量氮含量呈降低的趨勢,但是差異不顯著.葉干重在錫林浩特西種群中顯著高于其它2個種群(表5).
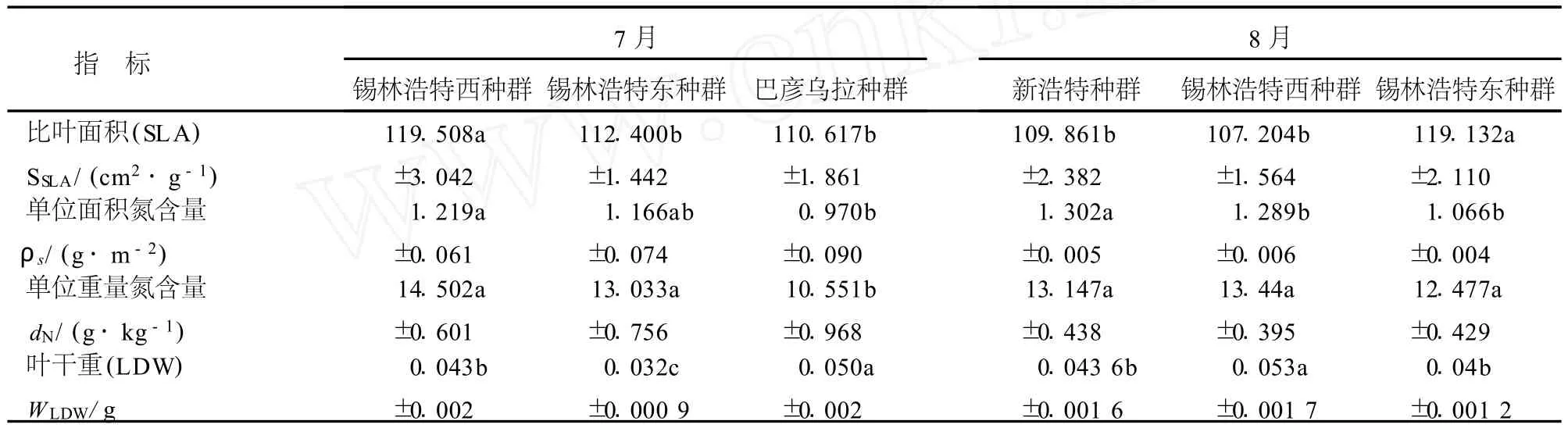
表5 不同地理種群大針茅葉性分析Table5 AnalysisofleaftraitswithindifferentS.grandispopulationsinJulyorAugust
3.2 不同時間相同生境下克氏針茅和大針茅的葉性分析
3.21 7月份
7月份在錫林浩特西和錫林浩特東樣地相同生境下大針茅的單位重量氮含量高于克氏針茅,但是差異不顯著;大針茅的單位面積氮含量和葉干重均顯著高于克氏針茅;大針茅的比葉面積低于克氏針茅,在錫林浩特東種群中差異極顯著(表6).
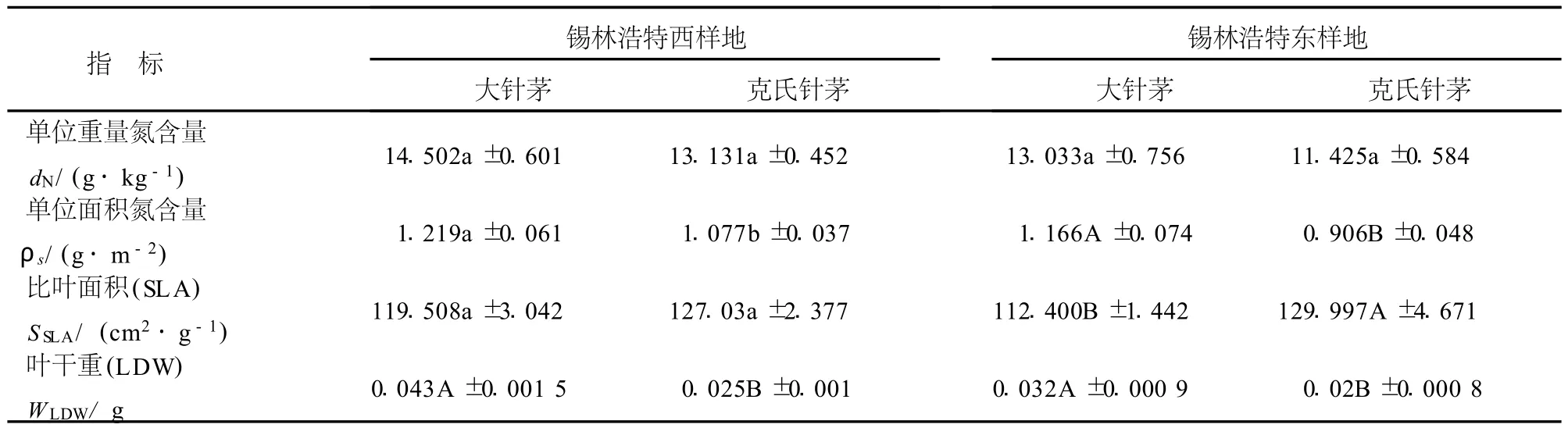
表6 7月份相同生境下大針茅和克氏針茅葉性分析Table6 LeaftraitsanalysesofS.grandisandS.kryloviiinthesamehabitatsinJuly
3.2.2 8月份
8月份相同生境下大針茅和克氏針茅的葉性分析表明,大針茅單位重量氮含量低于克氏針茅,在錫林浩特東和新浩特樣地表現出顯著或極顯著的差異;單位面積氮含量大針茅高于克氏針茅,在錫林浩特西樣地表現出顯著差異;在3個樣地中比葉面積均表現出克氏針茅極顯著的高于大針茅;而葉干重在3個樣地中均表現為大針茅極顯著高于克氏針茅(表7).
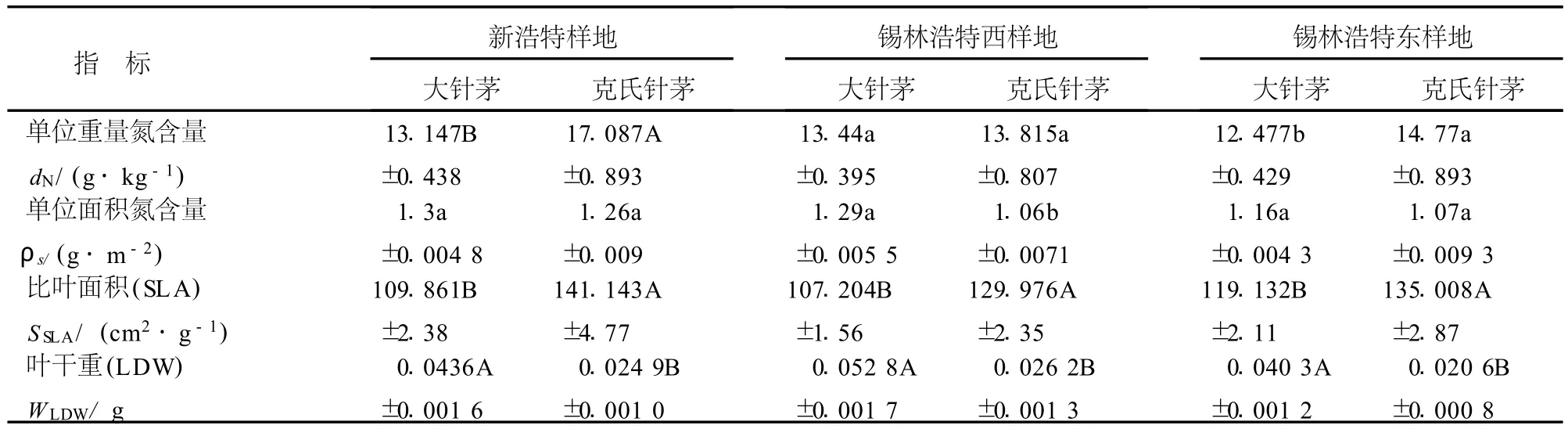
表7 8月份相同生境下大針茅和克氏針茅的葉性分析Table 7 Leaf traitsanalyses of S.grandis and S.k rylovii in the same habitats in August
4 討論與結論
4.1 不同生境下克氏針茅和大針茅SLA的適應策略
從典型草原西部的滿都拉圖到東部的巴彥烏拉,土壤類型、土壤全氮、全磷、有機質、降水量都發生了明顯的變化(表1、表2).這種變化趨勢反映出從典型草原西部到東部生境變得越來越肥沃、濕潤.這種生境變化特征影響著克氏針茅和大針茅的生長策略.SLA是物種生長策略的一個重要組成部分[30],它可以反映植物獲取資源的能力[31—32].有研究認為SLA較小的葉片可能有較多細胞或單個細胞有較大生物量,而具有較小SLA的植物一般有較強的支持和抵御功能,這在資源缺乏的環境中表現得尤為突出[33].具有高SLA的植物能適應資源豐富的環境,具有較低SLA的植物能很好地適應貧瘠的環境[34].
7月份典型草原西部新浩特克氏針茅種群的SLA與典型草原中、東部種群的SLA相比有顯著降低的變化趨勢,反映出生境較差的克氏針茅以較小的SLA適應環境的特征.8月份不同地理種群克氏針茅SLA表現出從典型草原西部向東部顯著減小的趨勢,這與7月份SLA的變化趨勢相反.結合表3中各樣地的實際降雨量分析可知:不同生境克氏針茅種群7月份和8月份SLA呈相反變化趨勢,這與生境中前期獲得的實際降雨量存在密切的關系.因為5月至6月各樣地獲得實際降雨量錫林浩特最高,新浩特次之,巴彥烏拉第三,滿都拉圖最低;各樣地5月至7月獲得的實際降雨量滿都拉圖最高,新浩特次之,錫林浩特第三,巴彥烏拉最低.8月初克氏針茅的采樣地分別是滿都拉圖、新浩特、錫林浩特,結合表3采樣之前克氏針茅種群從典型草原西部到東部獲得的實際降雨量逐漸減少,而7月初采樣之前4種群實際獲得的雨量是從典型草原西部種群到東部種群逐漸增加.7、8月克氏針茅SLA的變化趨勢與各樣地在采樣前期獲得的實際降雨量的變化趨勢相同,即樣地前期獲得的實際降雨高,克氏針茅SLA相應就高.
大針茅8月份的SLA從典型草原西部向東部呈升高的趨勢,與7月份大針茅SLA從典型草原西部到東部的變化也呈相反的趨勢(表5).本研究中大針茅7月初的采樣地分別是錫林浩特(2個種群)和巴顏烏拉,結合表3采樣之前,錫林浩特的實際降雨量高于巴彥烏拉;即7月份不同地理種群大針茅SLA的變化趨勢與采樣前各樣地的實際降雨量變化趨勢相同.8月初的采樣地為新浩特和錫林浩特(2個種群),結合表3采樣之前,新浩特獲得的實際降水量高于錫林浩特,新浩特種群和錫林浩特西種群SLA沒有顯著差異,錫林浩特東種群顯著高于其它兩個種群,這可能是由于所測氣象數據沒有定位到取樣點造成的.因此,本研究認為,克氏針茅和大針茅SLA的變化趨勢與生境中前期的實際降雨量的變化趨勢基本一致,即采樣前期生境中獲得的實際降雨量較高,克氏針茅和大針茅的SLA就相對較高.
4.2 不同生境中克氏針茅和大針茅葉氮含量的適應策略
氮是植物生長的主要限制因子之一[35—36],它在植物葉內的含量遠高于在其它器官中的含量.葉氮含量的增加直接影響光合酶(如RuBP羧化酶)的含量和活性,進而影響光合作用.通過對各生境中土壤全氮、全磷和有機質進行測定,結果表明從典型草原東部的巴彥烏拉到西部邊緣的滿都拉圖,生境變得越來越貧瘠.結合表4和表5對克氏針茅和大針茅葉氮含量的測定結果,本研究認為,從典型草原西部到東部7、8月份不同地理種群大針茅和7月份不同地理種群克氏針茅的Narea隨著生境變差(貧瘠)表現出顯著升高的趨勢;8月份滿都拉圖樣地克氏針茅由于整個生長季獲得的實際降雨較高,在8月取樣時已經進入果后營養期,可能造成營養枝葉氮的流失,表現出葉氮含量降低.除8月份滿都拉圖樣地克氏針茅外,其它生境中克氏針茅和大針茅Narea從典型草原西部到東部均表現出降低的趨勢,這與以往對干旱區植物葉性策略[37—39]的研究結果一致,反映出克氏針茅和大針茅Narea對不同生境的生存適應策略,即克氏針茅和大針茅在相對肥沃的生境中采取低Narea的生存適應策略,在相對貧瘠的生境中采取Narea的生存適應策略.
4.3 相同生境中大針茅和克氏針茅的生存策略
相同生境下,大針茅單位面積的葉氮含量高于克氏針茅,相同生境下大針茅的SLA顯著低于克氏針茅,而葉干重顯著高于克氏針茅.這表明在相同生境下大針茅采取高Narea、低SLA的生存策略;而克氏針茅則是采取低Narea,高SLA的生存策略.李永華等結合其他學者的研究結果揭示 SLA和Narea對植物水分利用效率具有重要的指示意義,在一定范圍內葉氮的增加和比葉面積的減少仍就能夠提高植物水分利用效率[40],本研究初步認為,與克氏針茅相比大針茅具有較高的水分利用效率.有學者研究表明,Nmass的多寡直接決定著葉片光合能力的高低[36],Narea高的葉片光合能力強[41].關于相同生境下克氏針茅和大針茅二者的葉氮含量與光合能力之間的關系還有待進一步研究.根據一些學者的研究揭示SLA易受到葉化學組成和解剖特征的影響,包括葉的干物質含量[33]、葉厚度[42]、葉含水量[43,44]等,Witkow ski ETF和 Ship ley B研究認為,SLA是葉厚度與葉組織密度的函數[42,44].草本植物的葉厚度影響CO2同化速率[45],與營養的獲得有關[42,46].Ian JW right和 Mark Westoby(2001)的研究[47]認為,假如SLA的變化是由葉厚度的變化引起,葉厚度由于富含氮的葉肉增加而增加,那么較低的SLA伴隨較高的Narea;假如SLA的變化大部分是由于葉組織密度,較高的組織密度來自于氮貧乏的細胞壁,就能夠得到相反的結論[4].本研究中克氏針茅的葉干重低于大針茅是由種群的特異性決定的,而在相同的生境下克氏針茅的SLA高于大針茅的原因,本文推測一方面可能是受葉厚度的影響,另一方面可能是由于二者葉片組織密度差異所造成的.
[1] M acGillivray CW,Grime J P.the ISP team.Testing p redic
tions of resistance and resilience of vegetation subjected to extreme events[J].Functional Ecology,1995,9:640-649.
[2] Co rnelissen J H C,Perez-Harguindeguy N,Diaz S,et al.Leaf structure and defence control litter decomposition rate across species and life fo rm s in regional flo ras on two continents[J].New Phytologist,1999,143:191-200.
[3] Field C,Merino J,Mooney H A.Comp romises between water-use efficiency and nitrogen-use efficiency in five species of California evergreens[J].Oecologia,1983,60:384-389.
[4] Field C.A llocating leaf nitrogen fo r the maximization of carbon gain:leaf age as a control on the allocation p rogram[J].Oecologia,1983,56:341-347.
[5] Reich PB,Walters M B,Ellsworth D S.From tropics to tundra convergence in plant functioning[J].Proceedings National Academy of Science,USA.1997,94:13730-13734.
[6] Kikuzawa K.A cost-benefit analysisof leaf habit and leaf longevity of trees and their geographical pattern[J] Am Nat,1991,138:1250-1263.
[7] Kikuzawa K.Leaf phenology as an op timal strategy for carbon gain in p lants[J].Canadian Journal of Botany,1995,73:158-163.
[8] 陳世鐄,李銀鵬,孟君,等.內蒙古幾種針茅特性和生態地理分布的研究[J].內蒙古農牧學院學報,1997,18(1):40-46.
[9] 李德新.放牧對克氏針茅草原影響的初步研究[J].中國草地學報,1980(2):1-8.
[10] 陳世鐄,李存福.強度放牧與天然草場資源的關系[J].內蒙古農牧學院學報,1986,7(5):81-86.
[11] 李青豐,易津,張力君,等.針茅種子萌發檢驗標準及幼苗發育特征的研究[J].草業科學,1995,12(4):51-52.
[12] 詹學命,李凌浩,李鑫,等.放牧和圍封條件下克氏針茅草原土壤種子庫的比較[J].植物生態學報,2005,29(5):747-752.
[13] 白永飛,徐志信.典型草原9種牧草生長發育規律的研究[J].中國草地,1994(6):21-27.
[14] 孟君,陳世鐄.克氏針茅繁殖的生態生物學特性[J].內蒙古農牧學院學報,1997,18(2):33-37.
[15] 張昊,李鑫,姜鳳和,等.水分對克氏針茅和冷蒿生殖生長的影響[J].草地學報,2005,13(2):106-110.
[16] 韓冰,趙萌麗,珊丹.不同退化梯度克氏針茅種群形態及等位酶的分析[J].草業科學,2004,21(12):78-82.
[17] 韓冰,王俊,趙萌麗,等.退化梯度對克氏針茅種群遺傳分化的影響[J].草地學報,2003,11(2):146-153.
[18] 趙念席,高玉葆,王金龍,等.內蒙古中東部草原區克氏針茅種群遺傳分化的RAPD研究[J].生態學報,2006,26(5):1312-1319.
[19] 賈美清,高玉葆,劉海英,等.內蒙古中東部草原不同生境克氏針茅(Stipa kry loviiRoshev.)種群的形態差異分析[J].植物研究,2008,28(5):608-613.
[20] 王金龍,高玉葆,趙念席,等.內蒙古中東部草原克氏針茅形態特征和RAPD遺傳分化的相關性研究[J].植物研究,2006,26(6):709-714.
[21] 王金龍,高玉葆,白宇,等.大針茅(Stipa grandis)和克氏針茅(Stipa kry lovii)對 PEG滲透脅迫適應性反應的比較研究[J].南開大學學報:自然科學版,2005,38(4):127-131.
[22] 何興東,叢培芳,高玉葆,等.利用壓力-容積曲線研究四種草本植物的抗旱性[J].南開大學學報:自然科學版,2006,39(3):16-21.
[23] 王艷芳,韓冰,張占雄.錫林郭勒盟草原克氏針茅抗旱生理變化的研究[J].草業科學,2006,23(2):22-26.
[24] 萬宏偉,楊陽,白世勤,等.羊草草原群落6種植物葉片功能特性對氮素添加的響應[J].植物生態學報.2008,32(3):611-621.
[25] 朱玉潔,高瓊,劉峻杉,等.基于氣孔導度和光合模型的植物功能類群合并問題[J].植物生態學報,2007,31(5):873-882.
[26] 張林,羅天祥.植物葉壽命及其相關葉性狀的生態學研究進展[J].植物生態學報,2004,28(6):844-852
[27] Garnier E,Shipley B,Roumet C,et al.A standardized protocol for the determination of specific leaf area and leaf dry matter content[J].Functional Ecology,2001,15:688-695.
[28] 白由路,楊俐蘋.基于圖像處理的植物葉面積測定方法[J].農業網絡信息,2004(1):36-38.
[29] 肖強,葉文景,朱珠,等.利用數碼相機和Photoshop軟件非破壞性測定葉面積的簡便方法[J].生態學雜志,2005,24(6):711-714.
[30] Westoby M.A leaf-height-seed(LHS)p lant ecology strategy scheme[J].Plant and Soil,1998,199:213-227.
[31] Wilson P,Thompson K,Hodgson J.Specific leaf area and leaf drymatter content as alternative p redictorsof plant strategies[J].New Phytologist,1999,143:155-162.
[32] Garnier E,Laurent G,Bellmann S,et al.Consistency of species ranking based on functional leaf traits[J].New Phytologist,2001,152:69-83.
[33] Castro-Díez P,Puyravaud J P,Cornelissen J H C.Leaf structure and anatomy as related to leaf mass per area variation in seedlings of a wide range of woody plant species and types[J].Oecologia,2000,124:476-486.
[34] 李玉霖,崔建垣,蘇永中.不同沙丘生境主要植物比葉面積和葉干物質含量的比較[J].生態學報,2005,25(2):305-311.
[35] ChapinⅢF S.Themineral nutrition of wild plants[J].Annual Review of Ecology and Systematics,1980(11):233-260.
[36] ChapinⅢF S,Bloom A J,Field C B,et al.Plant response to multiple environmental factors[J].Bio-Science,1987,37:49-57.
[37] W right IJ,Westoby M,Reich P B.Convergence towards higher leaf mass per area in dry and nutrient-poor habitats has different consequences for leaf life span[J].Journal of Ecology,2002,90:534-543.
[38] Reich PB,Ellsworth D S,Walters M B,et al.Generality of leaf trait relationships:a test across six biomes[J].Ecology,1999,80:1955-1969.
[39] W right IJ,Reich P B,Westoby M.Strategy shifts in leaf physiology structure and nutrient content between species of high-and low-rainfall and high-and low-nutrient habitats[J].Functional Ecology,2001,15:423-434.
[40] 李永華,羅天祥,盧琦,等.青海省沙珠玉治沙站17種主要植物葉性因子的比較[J].生態學報,2005(25):994-999.
[41] Evans J R.Photosynthesis and nitrogen relationships in leaves of C3plants[J].Oecologia,1989,78:9-19.
[42] Witkow ski E T F,Lamont B B.Leaf specificmass confounds leaf density and thickness[J].Oecologia,1991,88:486-493.
[43] Garnier E,Laurent G.Leaf anatomy,specific mass and water content in congenerics annual and perennial grass species[J].New Phytologist,1994,128,725-736.
[44] Shipley B.Structured interspecific determinants of specific leaf area in 34 species of herbaceous angiosperms[J].Functional Ecology,1995(9):312-319.
[45] Garnier E,Salager J L,Laurent G,et al.Relationships between photosynthesis,nitrogen and leaf structure in 14 grass species and their dependence on the basis of exp ression[J].New Phytologist,1999,143,119-129.
[46] Reich PB,Walters M B,Ellsworth D S,et al.Relationships of leaf dark respiration to leaf nitrogen,specific leaf area and leaf life-span-A test across biomes and functional groups[J].Oecologia,1998,114:471-482.
[47] W right IJ,Westoby M.Understanding seedling grow th relationships through specific leaf area and leaf nitrogen concentration:generalisations across grow th forms and grow th irradiance[J].Oecologia,2001,127:21-29.
Leaf traits of Stipa kryloviii Roshev.and S.grandis P.Sm irn.in m iddle and eastern Inner M ongolia steppe
J IA M eiqing1,2,GAO Yubao2,YANG Yong3,
(1.Key Laboratory of Water Environment and Water Resource,Tianjin Normal University,Tianjin 300387,China;2.College of Life Sciences,Nankai University,Tianjin 300071,China;3.Civil Aviation University of China,Tianjin 300300,China)
Three populationsofStipa kry loviiRoshev.in July and four populations of it in August,three populations ofS.grandisP.Smirn.collected from M andulatu,Xinhot,West Xilinhot,East Xilinho t,Bayanw ula,were used for the study of their leaf traits.From the western margin of the typical steppe in Inner Mongolia to the eastern of it,S.grandisandS.kry loviihad high area-based leaf nitrogen content in poor habitats,and in less poo r habitatsS.grandisandS.kry loviihad low area-based leaf nitrogen content.And the specific leaf area(SLA)ofS.grandisandS.kry loviichanged w ith the early actual p recipitation of their habitats,that is to say,high early p recipitation in the habitat lead to relatively high SLA.While in the same habitats,S.kry loviihad high SLA and low Narea,at the same timeS.grandishad low SLA and high Narea.Obviously,they took very different survivalmode in both SLA and Narea in order to adap ted environments.
Stipa kry loviiRoshev.;S.grand isP.Smirn.;specific leaf area(SLA);area based leaf nitrogen concentrations(Narea);mass based lesa nitrogen concentrations(Nmass)
Q143,Q948
A
1671-1114(2010)01-0057-07
2009-09-10
國家重點基礎研究發展規劃項目(2007CB106802);天津市科技發展計劃項目(06YFSZSF05100);國家自然基金項目(40803002);天津市科技計劃重點項目(09JCZDJC25900);天津市科技計劃一般項目(09JCYBJC07900)
賈美清(1978—),女,碩士,主要從事種群生態學研究.E-mail:jiameiqing@mail.nankai.edu.cn
高玉葆(1955—),男,教授,博士生導師,主要從事植物生理與生態學方面的研究.
(責任編校 李宏偉)
