利用小鼠感染模型篩選與鑒定幽門螺桿菌外膜蛋白抗原①
2010-02-06 04:37:58陶好霞袁盛凌展德文王令春王艷春劉純杰
中國免疫學雜志 2010年3期
陶好霞 王 袁盛凌 展德文 王令春 王艷春 劉純杰
(軍事醫學科學院生物工程研究所,北京 100071)
利用小鼠感染模型篩選與鑒定幽門螺桿菌外膜蛋白抗原①
陶好霞②王②袁盛凌 展德文 王令春 王艷春 劉純杰③
(軍事醫學科學院生物工程研究所,北京 100071)
目的:利用小鼠感染模型篩選與鑒定幽門螺桿菌SS1株的外膜蛋白抗原。方法:提取SS1株的外膜蛋白進行雙向電泳,用幽門螺桿菌感染的小鼠血清作免疫印跡實驗,將陽性反應蛋白點進行質譜鑒定分析,將肽質量指紋譜數據輸入互聯網上的蛋白質數據庫進行檢索。結果:獲得32種抗原相關蛋白。通過與已有報道的幽門螺桿菌感染人抗原比較分析,發現大部分典型的保護性抗原在本實驗中都可以檢測到。結論:幽門螺桿菌感染的小鼠模型適用于人用保護性幽門螺桿菌抗原的篩選;而且此研究中得到的相關抗原蛋白對于尋找與鑒定幽門螺桿菌未知保護性抗原也有參考價值。
幽門螺桿菌;動物模型;雙向電泳;免疫方法;抗原篩選
幽門螺桿菌(Helicobacterpylori,Hp)是一種革蘭氏陰性微需氧菌,寄生于人胃粘膜表面的粘液層。目前全世界一半以上的人感染Hp,而且最終有近1%~2%的感染者會發展為胃癌[1]。Hp感染的藥物治療可以采用抗生素、質子泵抑制劑和鉍劑聯合用藥來進行,但這些藥物的有效率只有80%~90%,并且對細菌清除后并不能防止Hp的再次感染,而現在面臨的最大問題就是Hp耐藥株的出現[2]。考慮到目前全球感染Hp需要治療的人數大大超過了三聯抗生素用藥可以治療的人數,因此研制出Hp疫苗成為控制Hp全球感染的唯一可行的途徑。已經證明的具有免疫保護作用的Hp抗原包括尿素酶(UreA和UreB)、VacA、CagA、過氧化氫酶(KatA)、分子伴侶GroES和GroEL[3-6]。但還沒有一種抗原可以在人體中實現100%的保護作用,在動物模型中的實驗表明多個抗原的聯合應用可以提高免疫保護效果。為了得到具有高效保護作用的Hp疫苗,首要任務是在Hp的蛋白質組中找到可以引發較強免疫反應的保護性抗原。Hp定植于胃粘膜層和胃上皮細胞的頂端,因此它的位于細菌外膜的蛋白和分泌性蛋白更易于激發宿主的CD4+T細胞反應,而且大部分已知的Hp保護性抗原都是位于外膜或是分泌性的[7]。
大多數的Hp疫苗臨床前研究都用小鼠作為感染治療模型。小鼠并不是Hp的天然宿主,但有一些Hp的菌株經過幾代馴化后可以定植于小鼠的胃中,如本實驗中用到的HpSS1株。在小鼠模型中有一些研究的疫苗能夠在Hp攻毒時產生較好的免疫保護效果,但用于臨床受試者時效果卻不十分理想。為了了解Hp的外膜蛋白在小鼠體內的表達情況,從而對小鼠模型用于Hp人類疫苗研究的可行性作出評價,我們采用了蛋白質組學的方法將Hp的外膜蛋白進行雙向電泳,然后用Hp感染的小鼠血清對得到的雙向分離蛋白點進行免疫印跡實驗,對得到免疫反應蛋白點進行鑒定分析,與已有報道的Hp感染病人的數據結果進行比較分析,從而對小鼠模型在用于Hp感染人類的保護性抗原的篩選的可行性上進行初步的研究。
1 材料與方法
1.1 材料 幽門螺桿菌HpSS1(Sydney strain1)由廣州中山醫科大學陳湖教授惠贈。免疫用Hp小鼠血清通過對小鼠進行活菌口服灌胃獲得。
1.2HpSS1外膜蛋白的制備及含量測定[8]采用空腸彎曲菌瓊脂基礎培養基(其中含10μg/ml鹽酸萬古霉素,2.5μg/ml多粘菌素B,2μg/m l兩性霉素B及7.5%脫纖維羊血),復蘇和擴增SS1菌種,用20 mmol/LTris·Cl(pH7.4)溶液收集和洗滌菌體2次,菌體沉淀用20mmol/LTris·Cl(pH 7.4)溶液懸浮,冰浴超聲之后加入RNA酶和DNA酶室溫消化核酸,離心棄去沉淀,將上清于4℃離心1小時。沉淀中加入10ml 20mmol/L Tris·Cl(pH7.4)和 2%十二烷基肌氨酸鈉(SKL),于4℃離心1小時。沉淀中加入30m l純水懸浮,離心,收集沉淀,用300μl純水徹底溶解,用2-D Quanti-kit試劑盒對蛋白進行定量分裝,1mg/支。直接上樣或-70℃保存備用。
1.3 雙向電泳 參照操作手冊[9],并根據G?rg等[10,11]的文獻進行了一些改進。
具體方法是:取兩份外膜蛋白各400μl(1mg)加入IPG緩沖液和水化液至總體積為350μl,然后于室溫振蕩30分鐘,使溶解徹底。4℃離心輕取上清,加于IPG phor電泳槽中,輕覆以 IPG干膠條(pH3-10),按照如下條件于17℃進行等電聚焦:水化0 V,6小時;30V,6小時;500 V,1小時;1 000 V,1小時;8 000 V,24小時。當電壓時間積達到4-6萬Vhrs時終止等電聚焦程序。第二向SDS-聚丙烯酰胺凝膠(SDS-PAGE):在Bio-Rad ProteanⅡXi垂直電泳儀上進行。將兩個膠條依次用100 mg DTT加入10 ml平衡緩沖液和450mg碘乙酰胺加入10m l平衡緩沖液中進行平衡,再將兩個膠條同時轉入兩塊12.5%SDS-PAGE膠面上端進行第二向SDS-PAGE電泳。其中一塊SDS-PAGE膠用考馬斯亮蘭G-250染色,再用ImageScanner進行掃描,所用掃描軟件為LabScan;考染凝膠掃描后用保鮮膜包裹,4℃保存以便取點時使用。圖像分析用ImageMaster 2D Elite 3.1和ImageMaster 2D Elite Platnum進行。蛋白質點的分子量根據同步電泳的標準蛋白的位置計算,等電點直接按所用IEF膠條的pH范圍計算,并通過軟件獲取每個蛋白點的豐度值。
1.4 Western blot 將平行操作的另一塊SDS-PAGE膠電轉1小時至硝酸纖維素膜。然后用麗春紅室溫染色10分鐘,純水沖洗脫色至清晰,掃描圖像。加入5%脫脂奶粉于4℃封閉過夜,加入用SS1免疫的小鼠血清,37℃輕搖1小時,用TBST洗膜3次后,加入用5%脫脂奶粉稀釋的二抗,37℃輕搖1小時,用TBST洗膜3次后,進行ECL顯色。
1.5 膠內酶切、MALDI-TOF-MS質譜鑒定與數據庫檢索[12]將與Western blot中起反應的免疫反應蛋白點對應于SDS-PAGE膠上的點切下,放入50μl脫色液中脫色,真空離心干燥,再分別用 5%三氟乙酸、2.5%三氟乙酸/50%乙腈和100%乙腈抽提,真空離心干燥,用0.5%三氟乙酸充分溶解沉淀,用于質譜分析。將a-氰基-4-羥基肉桂酸(a-CCA)溶于含0.1%三氟乙酸的50%乙腈溶液中,制成飽和溶液,離心,取1μl上清與1μl肽段提取液等體積混合,取1μl點在Scorce384靶上,送入離子源中進行檢測。檢測:反射檢測方式;飛行管長 3m;氮激光器:波長337 nm;加速電壓20 kV;反射電壓為23 kV。登陸http://www.matrixscience.com,用 Mascot程序對MALDI-TOF質譜檢測得到的肽質量指紋圖譜進行檢索。數據庫選NCBInr或MSDB,種屬選Proteobacteria。氨基酸固定修飾方式一般選擇Carbam idomethyl(C)修飾(或選擇為可能修飾)、可能的修飾方式選Oxidation(M)修飾。檢索時選取單同位素峰,一個誤切,可接受的肽段分子量誤差一般設為0.3 Da。為使鑒定結果更可靠,一般情況下要求至少有5個肽匹配,在蛋白質整個序列的覆蓋率至少15%,但對于小于20 kD的蛋白質,則要求至少3個肽匹配和20%的覆蓋率。
1.6 蛋白質表達情況的分析 根據基因組注釋(www.tigr.org)所預測的等電點和分子量與電泳凝膠上蛋白質點在不同區段的分布,分析雙向電泳參考圖譜的覆蓋情況;根據所鑒定蛋白質點在凝膠中的等電點、分子量值與預測的等電點、分子量作圖,分析二者之間的符合情況及可能的翻譯后修飾情況;根據基因組注釋所提供的蛋白質功能分類及定位情況,統計所鑒定的蛋白質在不同的功能分類及細胞內定位中的分布和覆蓋情況。
2 結果
2.1Hp外膜蛋白雙向電泳圖譜的建立 使用pH值為3~10的IPG干膠條,通過3次相對獨立的實驗提取Hp的外膜蛋白進行電泳,得到重復性較好的Hp外膜蛋白雙向電泳圖譜(圖1)。
2.2 免疫蛋白點的篩選和鑒定 先用麗春紅進行可逆染色對蛋白質點進行定位,然后將自制的用HpSS1株全菌灌胃提取的小鼠血清進行Western blot(圖2);從考馬斯亮蘭染色凝膠上找出顯色的點所對應的蛋白質點,切取該點經胰蛋白酶膠內酶切后,使用MALDI-TOF質譜成功鑒定了32個免疫反應蛋白點(表1)。這些蛋白點可以歸為以下幾類:與細胞泳動相關的蛋白(T1、T7、T8、T30);細胞加工蛋白(T3、T4、T9);酶類(T5、T11、T16、T17、T19、T20、T22、T25、T26、T29、T32);翻譯相關蛋白(T2、T6、T12、T13、T24);假想 蛋白(T15、T23、T27、T31);調控 蛋白(T21);金屬螯合相關蛋白(T28);細胞分裂相關蛋白(T10);胞壁合成相關蛋白(T18、T14)。
2.3 免疫結合蛋白與人血清鑒定到的抗原的比較將鑒定到的32個蛋白的全稱,或者蛋白TIGR號(如HP0072)作為搜索關鍵詞,在網站PubMed(www.pubmed.com,www.ncbi.nlm.nih.gov)中進行文獻檢索,經過比較分析共有17種作為抗原在Hp感染的人類血清中也可以檢測到。將這些抗原的名稱,在細胞中的位置及與人血清反應的特異性列于表2中。
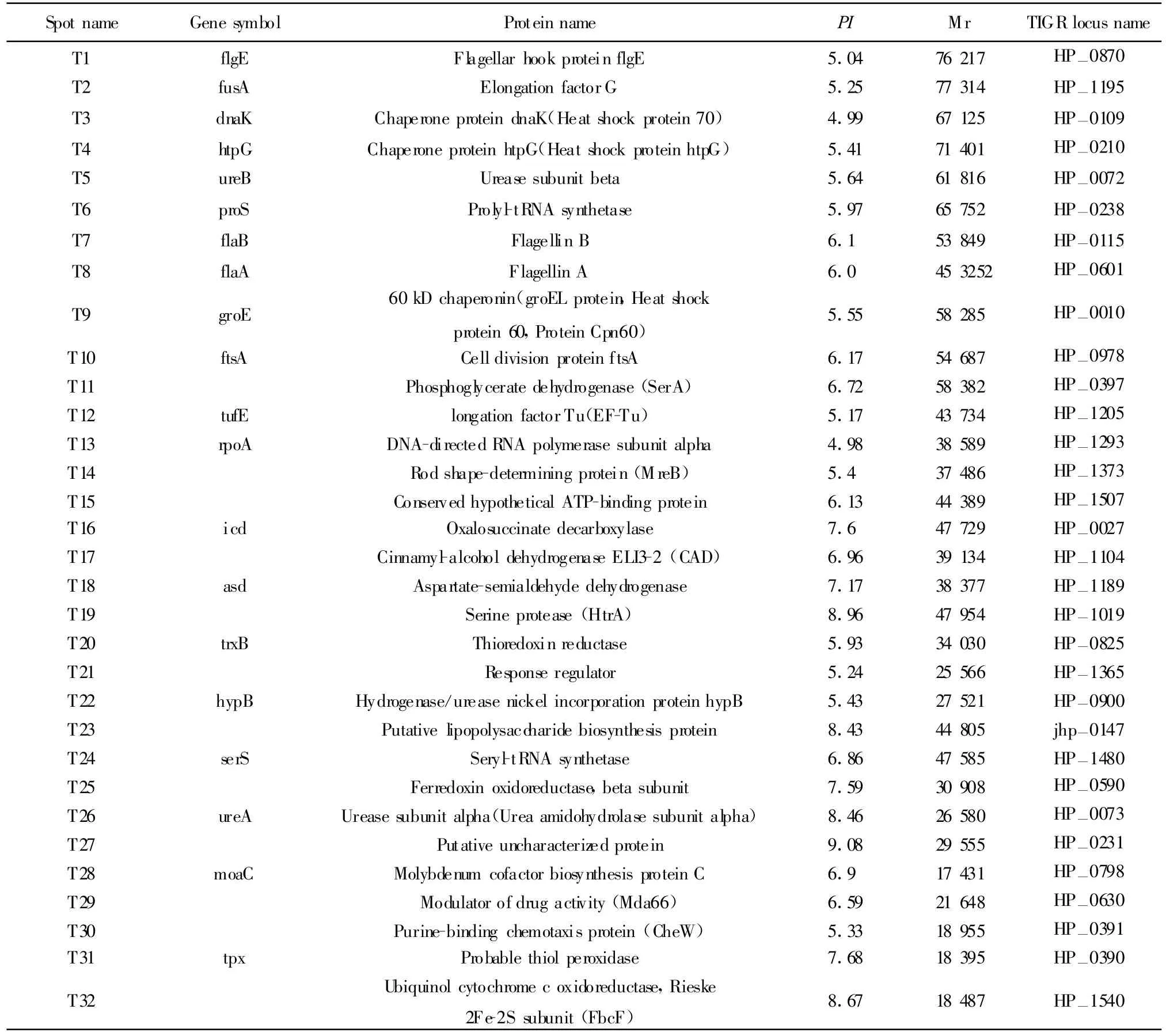
表1 質譜鑒定的免疫反應蛋白點Tab.1 Identification of immunogenic H.pylori p roteins by Immuno blotand M S
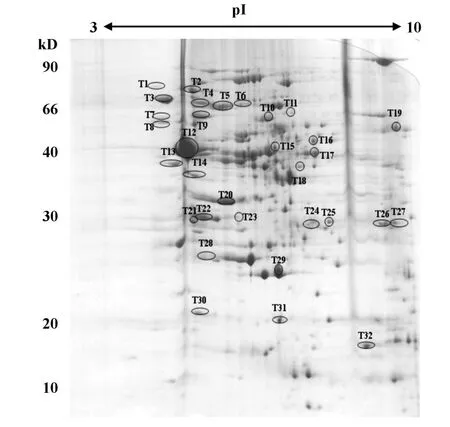
圖1 Hp SS1外膜蛋白雙向電泳圖Fig.1 2D map(pH 3 to10)of outer membrane proteins of Hp SS1
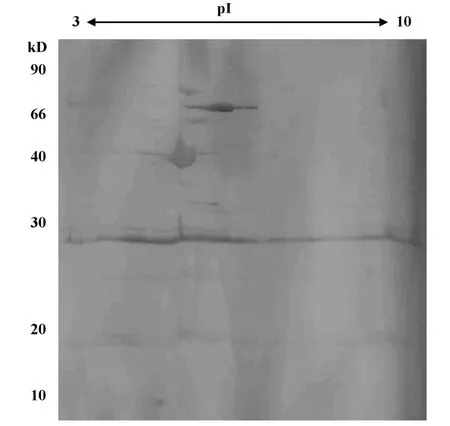
圖2 Hp SS1胞外膜蛋白的小鼠血清W estern blotFig.2 W estern blot of a 2D gel hybridized with serum from murinemodel
3 討論
雙向電泳的一塊PAGE膠上可以分辨100×100個點,結合Western blot方法可以大大提高抗原蛋白的檢出率,因此免疫蛋白質組學是尋找抗原的良好技術平臺。本研究運用該技術體系,在HpSS1株的外膜蛋白中成功鑒定了32個與小鼠血清進行免疫反應的蛋白點。
細菌的泳動和趨化能力與它的毒力密切相關。本研究鑒定出了與Hp鞭毛合成和細菌趨化作用相關的有4個蛋白:FlgE(HP0870)、FlaA(HP0601)、FlaB(HP0115)和CheW(HP0391)。FlaA和FlaB是位于細胞外的鞭絲的組成亞基。FlaA和FlaB是研究較多的Hp鞭毛抗原,它們可以在宿主體內引發較強的免疫反應[13,14]。FlgE位于外膜,它組成了細菌鞭毛鞭絲與基體動力裝置相連接的彈性結構,flgE基因的缺失會導致鞭毛鉤體無法形成,進而使細菌喪失泳動能力。CheW是假想嘌呤結合的趨化蛋白,它可以將趨化因子的受體信號跨膜傳送到鞭毛的動力系統[15]。cheW基因缺失的Hp的趨化能力喪失,并且在小鼠胃內引發的炎癥反應減弱[16]。
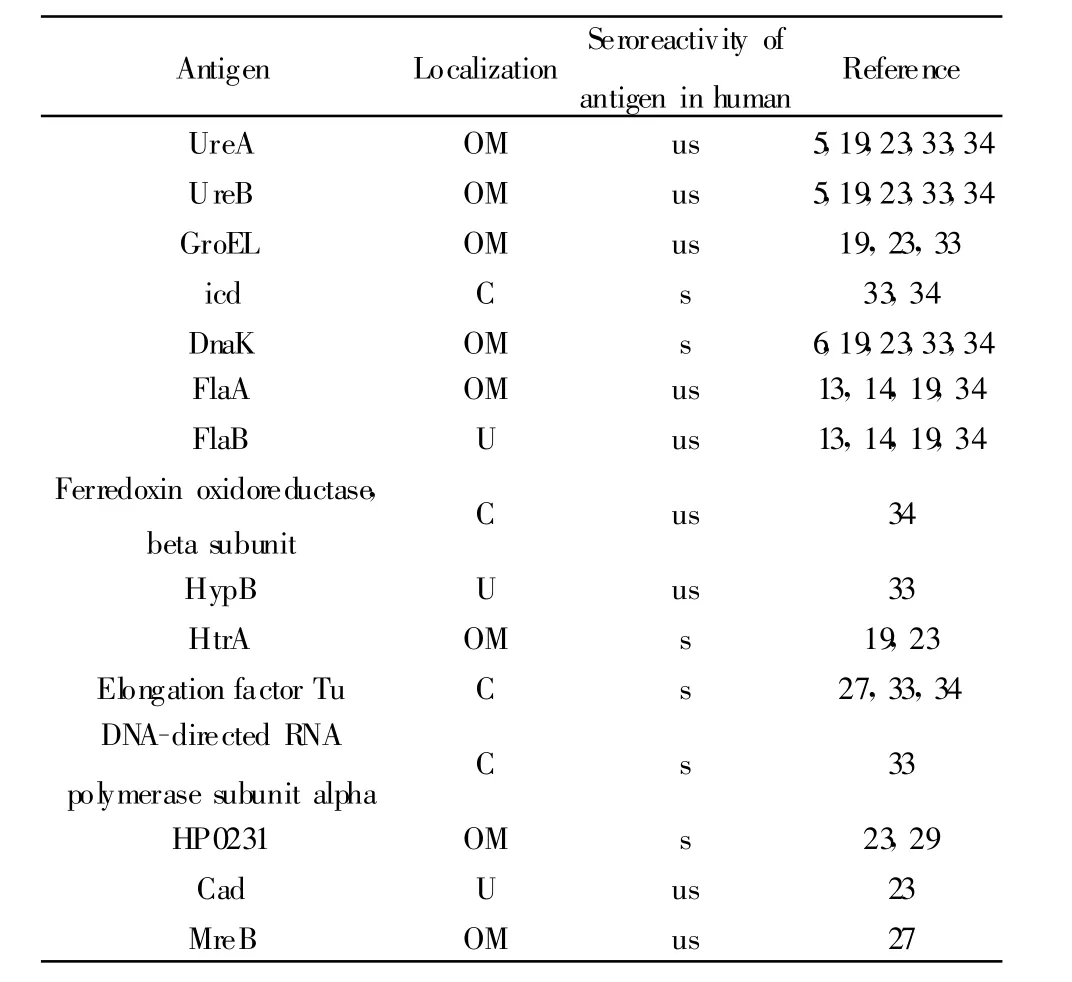
表2 小鼠血清中檢測到的幽門螺桿菌抗原的相關特性Tab.2 Characteristic of Helicobacter pylori antigen from murinemodle
在Hp中,熱休克蛋白(HSP)可以介導關鍵蛋白的分泌和折疊,使蛋白活性部位不會暴露于酸性環境中,從而對細菌起到保護作用。共鑒定到了3個具有免疫原性的屬于HSP家族的蛋白:DnaK(hsp70,HP0109)、HtpG(hsp90,HP0210)和 GroEL(hsp60,HP0010)。GroEL在調控尿素酶活性上起到一定作用[17],DnaK還可以起到粘附素的作用[18],對抗不利的外界環境介導Hp定植于胃粘膜。DnaK和GroEL都可以引發機體胃表皮慢性炎癥的發生,它們都可以作為Hp的備選保護性抗原[19]。
鑒定的蛋白中很多具有酶活性,大致可以把它們分為五類:(1)毒力相關酶類。尿素酶是Hp的重要毒力蛋白,它能夠中和胃酸,對細菌起到保護作用。尿素酶由UreA和UreB兩個亞單位組成,它們均為Hp保護性抗原,在實驗中也鑒定到了這兩種蛋白。而蛋白HypB(HP0900)是作為鎳特異性的金屬分子伴侶起作用的,它參與鎳離子的固定和將鎳離子轉移至尿素酶的金屬結合區[20]。(2)物質代謝相關酶類。其中磷酸甘油酸脫氫酶(SerA,HP0397)是細菌甘油代謝的重要酶。草酰琥珀酸脫羧酶(HP0027)參與了細菌的三羧酸循環。桂醇脫氫酶(CAD,HP1104)催化p-羥基桂醇類物質向其相應醇類物質的可逆轉換,HP1104編碼的CAD還具有歧化酶活性,它可以催化苯甲醛為苯甲醇或苯甲酸,從而降低細菌細胞內的醛類物質的濃度,利于細菌在氧應激條件下生存,在酸刺激條件下,Hp的CAD產物上升了24倍[21,22];并且有研究者在胃癌患者的血清中檢測到了HpCAD的抗體[23]。(3)氧化還原作用酶類。Hp在宿主胃粘膜上生存必然要應對外界環境中物理、化學或生物因素的刺激,應對于此,細菌自身具有氧化還原功能的酶類的分泌會提高,從而對有殺傷細菌作用的蛋白分子產生還原作用,保護細菌免受傷害。本實驗中的相關酶包括硫氧化還原蛋白還原酶(TrxB,HP0825),它能夠特異還原胰島素、粘液素、IgG和IgA內部的二硫鍵,另外研究證實TrxB能夠幫助Hp通過接觸性還原反應將胃粘膜層中連接粘蛋白單體之間的半胱氨酸連接區的二硫鍵打破,從而使Hp通過難以穿越的胃粘液層在胃粘膜層得以定植[24]。還有Mda66,它是一種NADPH醌還原酶蛋白,能夠將醌還原為苯二酚,細胞內的醌代謝對于細菌應對環境的氧化應激反應有著直接的影響,因此Mda66的還原形態在應對氧化應激反應中發揮了重要作用。有報道稱mda66的基因突變株對于氧化應激反應敏感并且能夠降低在鼠胃部中的定植能力[25]。除此之外,我們還鑒定到的具有氧化還原作用的酶包括鐵氧化還原蛋白氧化還原酶的β亞基(HP0590)和細胞色素c氧化還原酶(HP1504),它們都是細菌呼吸鏈的重要成分。(4)蛋白酶類。HP1019為假想的絲氨酸蛋白酶和分子伴侶H trA,實驗證明HtrA是一種具有蛋白水解活性的分泌酶,作為一種功能蛋白它既存在于細胞周質中起熱應激降解作用,也存在于胞外基質中起毒力因子的作用[26]。
我們還鑒定了與蛋白質翻譯相關的5種蛋白,它們是延伸因子 EF-G(FusA,HP1195)、EF-Tu(HP1205)、脯氨酸 tRNA合成酶(ProS,HP0238)、絲氨酸tRNA合成酶(SerS,HP1480)、DNA介導的RNA聚合酶α亞基(RpoA,HP1293)。其中EF-G、EF-Tu和RpoA都可以作為抗原蛋白與感染Hp病人的血清進行抗原抗體結合反應[19,27]。在我們鑒定到的蛋白中,有四種為假想蛋白,它們分別是假想的ATP結合保守蛋白(HP1507)、假想的脂多糖合成蛋白(Jhp0147)、假想的功能不明蛋白(HP0231)、假想的巰基過氧化物酶Tpx(HP0390)。其中Jhp0147編碼的酶證明具有葡萄糖轉移酶功能,與α-1,6-葡聚糖聚合體的合成有關,該聚合體是細菌核心LPS的一部分[28]。HP0231是近年來研究的比較多的一個蛋白,實驗證明它是一種保護性抗原,用重組表達的HP0231免疫小鼠然后用Hp攻毒,其保護效果接近Hp免疫實驗的金標準的4倍劑量[29]。Tpx蛋白體外實驗能夠還原H2O2,Hp的tpx突變株比野生株更易被過氧化物和超氧化物殺死,并且tpx突變株在小鼠胃內的定植能力降低[30]。
本實驗中我們鑒定到了二元調控系統HP1364/HP1365中的調控蛋白HP1365,該二元系統對于銅誘導的CrdA的轉錄和Hp對于銅離子的抗性是必須的,而且在小鼠模型中證明它們對于Hp在胃中的定植是必須的[31]。鑒定到的另一種與銅離子代謝有關的蛋白稱為銅離子輔助合成蛋白C(MoaC,HP0798),它參與銅離子的攝取。鑒定到的蛋白還有與細胞分裂相關的蛋白FtsA(HP0978),以及與胞壁合成相關的蛋白,這包括M reB(HP1373),它編碼假想的盤尼西林結合蛋白1A,并且參與細胞壁的合成[32];以及天冬氨酸半醛脫氫酶(Asd,HP1189),它是細菌DAP合成途徑的關鍵酶,而DAP是革蘭氏陰性菌細胞壁的主要成分。
經過與文獻比對,本研究鑒定到的蛋白中有17種蛋白與已有報道的與人血清有抗原抗體結合反應的Hp蛋白相一致。而其中之所以存在有胞內蛋白,是因為Hp具有自發裂解的特性,會使一些胞內蛋白釋放到胞外,從而使該蛋白也具有抗原性[35]。
本研究通過將HpSS1菌株的外膜蛋白中與細菌感染的小鼠血清呈陽性反應的蛋白用雙向電泳和免疫印跡的方法進行總體的篩選和鑒定,得到蛋白點經過文獻檢索發現很大一部分都與Hp的毒力和在宿主內的定植能力密切相關,再把得到的蛋白與已有報道的與Hp感染的人血清反應陽性抗原進行比較,結果發現兩者具有較高的相似性,說明小鼠感染的模型適于臨床應用前的Hp抗原候選蛋白的篩選,而且本實驗中所得到的一些蛋白點于未知的Hp的保護性抗原的選擇也有借鑒意義。
1 Peek RM Jr,BlaserM J.Helicobacter pyloriand gastrointestinal tractadenocarcinomas[J].Nat Rev Cancer,2002;2:28-37.
2 BuckleyM JM,Deltenre M.Therapy of H.pylori infection[J].Curr Opin Gastroenterol,1997;13:56-62.
3 Ferrero R L,Thiberge JM.TheGroEShomologofHelicobacter pyloriconfers protective immunity againstmucosal infection inmice[J].Proc Natl A-cad Sci USA,1995;92:6499-6503.
4 Marchetti M,Rossi M.Protection against Helicobacter pylori infection in miceby intragastric vaccinationwith H.pylori antigens isachieved using a non-toxicmutant of E.coli heat-labile enterotoxin(LT)as adjuvant[J].Vaccine,1998;16:33-37.
5 Pappo J,ThomasW D,Kabok Z.Effectoforal immunizationwith recombinant urease onmurineHelicobacter felisgastritis[J].Infect Immun,1995;63:1246-1252.
6 Radcliff F J,HazellS L,Kolesnikow Tetal.Catalase,a novel antigen for Helicobacter pylori vaccination[J].Infect Immun,1997;65:4668-4674.
7 Sutton P.Progress in vaccination against Helicobacter pylori[J].Vaccine,2001;19:2286-2290.
8 UttM,Nilsson I,LjunghAetal.Identification of novel immunogenic proteinsof Helicobacter pylori by proteome technology[J].JImmunolMethods,2002;259:1-10.
9 Tom B,Tirra S.2-D Electrophoresis:using immobilized pH gradients,Principles&Methods[M].Upsala:Amersham Pharmacia Biotech,Inc.,1998.
10 G?rg A,ObermaierC,Boguth Getal.The current stateof two-dimensional electrophoresis with immobilized pH gradients[J].Electrophoresis,2000;21(6):1037-1053.
11 G?rg A,Weisss W,Dunn M J.Current two-dimentional electrophoresis technology for proteomics[J].Proteomics,2004;4(12):1-21.
12 Cordwell S J.Acquisition and archiving of information for bacterial proteomics:from sample preparation to database[J].Methods:Enzymol,2002;358:207-227.
13 Tang RX,Luo D J,Sun AHetal.Diversity of Helicobacter pylori isolates in expression of antigens and induction of antibodies[J].World J Gastroenterol,2008;14(30):4816-4822.
14 Yan J,Liang XH,Mao Y Fetal.Construction of expression systems for flaA and flaB genes of Helicobacter pylori and determination of immunoreactivity and antigenicity of recombinant proteins[J].World JGastroenterol,2003;9(10):2240-2250.
15 Fredrick K L,Helmann JD.Dual chemotaxis signaling pathways in Bacillus subtilis:a sigmaD-dependentgene encodesa novel proteinwith both CheW and CheY homologous domain[J].J Bacteriol,1994;176:2727-2735.
16 Williams SM,Chen Y T,Andermann TMetal.Helicobacter pylori chemotaxismodulates inflammation and bacterium-gastric epithelium interactions in infectedmice[J].Infect Immun,2007;3747-3757
17 EvansD J,Evans D G,Engstrand Letal.Urease-associated heat shock protein ofHelicobacter pylori[J].Infect Immun,1992;60:2125-2127.
18 HuescaM,Borgia S,Hoffman Petal.Acidic pH changes receptor binding specificity of Helicobacter pylori:a binary adhesion model in which surface heat shock(stress)proteinsmediate sulfatide recognition in gastric colonization[J].Infect Immun,1996;64:2643-2648.
19 Lin Y F,Chen CY,TsaiM Hetal.Duodenalulcer-related antigens from Helicobacter pylori[J].MolCell Proteom ics,2007;6:1018-1026,
20 Stephane Benoit,Maier R J.Dependence ofHelicobacter pyloriurease activity on thenickel-Sequestering ability of theureE accessory protein[J].JBacteriol,2003;185:4787-4795
21 Ang S,Lee C Z,Peck Ketal.Acid-induced gene expression in Helicobacter pylori:study in genom ic scale bymicroarray[J].Infect Immun,2001;69:1679-1686.
22 Blanaid M,Dermot K,Jesus Fetal.Characterization of cinnamyl alcohol dehydrogenase ofHelicobacter pylori[J].FEBS,2005;272:1255-1264.
23 HaasG,Karaali G,Ebermayer Ketal.ImmunoproteomicsofHelicobacter pylori infection and relation to gastric disease[J].Proteomics,2002;2:313-324.
24 Henry JW,Fox A,Eidhin D Netal.The Thioredoxin system of Helicobacter pylori[J].JBiol Chem,2000;275(7):5081-5089.
25 Huang Z G,Duan G C,Fan Q Tetal.Mutation of cytotoxin-associated gene A affects expressions of antioxidant proteins of Helicobacter pylori[J].World JGastroenterol,2009;7:599-606.
26 Martin L,ChristianeW,Metzler Detal.Prediction of extracellular proteasesof thehuman pathogen helicobacter pylori reveals proteolytic activity of the Hp1018/19 Protein HtrA[J].PLoSONE,2008;3(10):e3510.
27 Lin Y F,Wu M S,Chang CCetal.Comparative immunoproteom icsof identification and characterization of virulence factors from helicobacter pylori related to gastric cancer[J].Mol Cell Proteomics,2006;5:1484-1496.
28 Susan M L,Eleonora A,Oksana Metal.Novelbiosynthetic functions of lipopolysaccharide rfaJ homologs from Helicobacter pylori[J].Glycobiology,2005;15(7):721-733.
29 Sabarth N,HurwitzR,Meyer T Fetal.Multiparameter selection ofHelicobacter pylori antigens identifies two novel antigenswith high protective efficacy[J].Infect Immun,2002;70:6499-6503.
30 WangG,O lczak A,Walton JPetal.Contribution of theHelicobacterpylori thiol peroxidase bacterioferritin com igratory protein to oxidative stress resistance and host colonization[J].Infect Immun,2005;73:378-384.
31 BarbaraW,KlausM,Frank N Setal.The Helicobacter pylori CrdRS two-component regulation system(HP1364/HP1365)is required for copper-mediated induction of the copper resistance determ inant CrdA[J].J Bacteriol,2005;187:4683-4688.
32 Kwon DH,DoreM P,Kim JJetal.High-levelβ-Lactam resistanceassociated with acquired multidrug resistance in Helicobacter pylor[J].Antimicrob AgentsChemother,2003;47:2169-2178.
33 Kimmel B,Bosserhoff A,Frank Retal.Identification of immunodom inant antigens from Helicobacter pylori and evaluation of their reactivities with sera from patientswith differentgastroduodenal pathologies[J].Infect Immun,2000;68:915-920.
34 M cAteeCP,Lim M Y,Fung Ketal.Identification ofpotentialdiagnostic and vaccine candidates of Helicobacter pylori by two-dimensional gel electrophoresis,sequence analysis,and serum profiling[J].Clin Diagn Lab Immunol,1998;5:537-542.
35 Phadnis SH,Parlow M H,Levy Metal.Surface localization of Helicobacter pylori urease and aheatshock protein homolog requiresbacterial autolysis[J].Infect Immun,1996;64:905-912.
[收稿2009-11-20]
(編輯 許四平)
Antigen screening and identification of Helicobacter pyloriouter membrane proteins from murine infection model
TAOHao-Xia,WANGPeng,YUANSheng-Ling,ZHANDe-Wen,WANGLing-Chun,WANGYan-Chun,LIUChun-Jie.BeijingInstituteofBiotechnology,InstituteofBiotechnology,MilitaryMedicalSciences,Beijing100071,China
Objective:To screen antigen of Helicobacter pylorioutermembrane proteins bymurine infectionmodel.Methods:Parallel two-dimensional gel electrophoresis(2D)of outermembrane p roteins extracted from Helicobacter pyloristrain SS1 was performed.Western blot of a duplicate2D gelhybridized with serum fromH.pylori-infectedmurinewas employed.ImmunogenicH.pyloriproteins identified in this way were digested ingel by trypsin and themass of generated peptidesweremeasured bymatrix assisted laser desorption ionization timeof flight mass spectrometry(MALDI-TOF-MS).The data obtained from peptidemass finger-printing(PMF)were searched using the internet availab le database.Results:32 proteinswere identified and they are in good agreementwith typical protectiveantigenswhich reacted with serum fromH.pylori-infected patients.Conclusion:The results suggest thatmurinemodel ofH.pylorimay be valid to screen antigens forhuman vaccination and the proteins identified in this paper are valuable for the selection ofH.pyloriprotective antigens aswell.
Helicobacterpylori;Animal diseasemodel;Two-dimensional gel electrophoresis;Immunology;Antigen screening
R391.2
A
1000-484X(2010)03-0195-06
①本文為“十一五”國家863重大項目(2006AA 02A219)
②共同并列第一作者
③通訊作者,E-mail:liucj@nic.bm i.ac.cn
陶好霞(1968年-),女,實驗師;王 (1976 年 -),女,博士,助理研究員。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
中老年保健(2021年3期)2021-08-22 06:50:04
昆明醫科大學學報(2021年1期)2021-02-07 01:06:36
現代臨床醫學(2021年1期)2021-01-26 00:56:02
中華養生保健(2020年4期)2020-11-16 01:31:40
中西醫結合肝病雜志(2020年2期)2020-10-27 02:18:50
學苑創造·A版(2020年9期)2020-10-13 09:41:02
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
現代檢驗醫學雜志(2014年4期)2014-02-02 02:44:59