調(diào)寧蛋白表達(dá)水平與慢性乙型肝炎患者肝臟微循環(huán)障礙的關(guān)系
2010-03-14 11:27:42代雪楓嚴(yán)家春李峻峰徐長(zhǎng)江袁蘇娜
實(shí)用肝臟病雜志 2010年2期
代雪楓 嚴(yán)家春 李峻峰 徐長(zhǎng)江 袁蘇娜
調(diào)寧蛋白(Calponin,CaP)是平滑肌所特有的重要調(diào)控蛋白。研究顯示,它具有調(diào)節(jié)平滑肌收縮、抑制細(xì)胞增殖及參與細(xì)胞骨架的維持等作用[1~3]。慢性乙型肝炎(CHB)的一個(gè)重要病理變化就是肝臟微循環(huán)障礙。基于肝臟組織血液供應(yīng)的特殊性,有必要對(duì)CaP在CHB患者肝臟血管改建中的作用進(jìn)行研究。我們對(duì)200例CHB患者肝活檢組織進(jìn)行了CaP檢測(cè),以探討它們之間的關(guān)系。
材料與方法
一、材料與分組 收集2004年1月至2007年12月我院200例CHB肝活檢組織,男143例,女57例,年齡15~57歲(平均年齡29.5±8.5歲)。按照國(guó)內(nèi)診斷標(biāo)準(zhǔn)[4],包括 CHB 輕度(G1~G2)、CHB 中度(G3)和CHB重度(G4)及肝硬化(LC)各50例。經(jīng)免疫組化檢測(cè),患者肝組織HBsAg皆陽(yáng)性。另選10例肝組織HBsAg陰性,肝組織學(xué)大致正常者作對(duì)照。
二、肝組織學(xué)檢查 采用16號(hào)肝活檢針抽取2條肝組織,其長(zhǎng)度≥1.5cm。其中一份用10%福爾馬林固定,常規(guī)制片,備HE及免疫組化染色;另一份按照電鏡技術(shù)制樣,備電鏡觀察。根據(jù)肝竇Ⅳ型膠原分布范圍,將肝竇毛細(xì)血管化定為輕(占肝小葉≥1%)、中(占肝小葉≥33%)、重型(占肝小葉或假小葉≥60%)。
三、肝組織CaP免疫組化檢測(cè) 采用S-P法檢測(cè)肝組織CaP的表達(dá)。抗CaP單克隆抗體及免疫組化試劑盒購(gòu)自北京中山生物技術(shù)有限公司。判斷標(biāo)準(zhǔn):陰性(-):無(wú)明顯陽(yáng)性細(xì)胞;陽(yáng)性(+):顯棕黃色,肝細(xì)胞質(zhì)膜散在表達(dá),肝竇及毛細(xì)血管灶性或斷續(xù)表達(dá),其表達(dá)范圍均占所觀察視野的10%~30%;強(qiáng)陽(yáng)性(++):顯深棕黃色或黃褐色,肝細(xì)胞、肝竇壁內(nèi)皮細(xì)胞、膽管細(xì)胞、成肌纖維細(xì)胞、血管纖維間隔及炎癥區(qū)的梭形細(xì)胞等均有表達(dá),表達(dá)范圍占所觀察視野的30%以上。
四、統(tǒng)計(jì)學(xué)處理 數(shù)據(jù)采用x2檢驗(yàn),P<0.05認(rèn)為有顯著性差異。
結(jié) 果
一、調(diào)寧蛋白在CHB肝組織的定位 在對(duì)照組,CaP僅在匯管區(qū)的血管及膽管壁有弱表達(dá),其余肝組織無(wú)表達(dá);而在CHB組,CaP在肝實(shí)質(zhì)及肝間質(zhì)細(xì)胞均有表達(dá)。在CHB肝組織,CaP表達(dá)主要分布于肝細(xì)胞(圖1)、小膽管或膽管樣細(xì)胞(圖2)、肝竇壁內(nèi)皮細(xì)胞(圖3)、肌纖維帶(圖4)、毛細(xì)血管增生區(qū)域(圖5)、貯脂細(xì)胞及成肌纖維細(xì)胞(圖6)。免疫組化染色提示,CaP表達(dá)變化有以下特點(diǎn):A:有肝竇擴(kuò)張或阻塞的內(nèi)皮細(xì)胞及貯脂細(xì)胞示陽(yáng)性表達(dá),纖維帶內(nèi)的小血管及其周邊的肝細(xì)胞、炎癥區(qū)域周圍的肝細(xì)胞、假小葉的肝細(xì)胞則示強(qiáng)陽(yáng)性表達(dá);B:纖維瘢痕組織中的膽管或膽管樣細(xì)胞及成肌纖維細(xì)胞示陽(yáng)性表達(dá);C:肝竇毛細(xì)血管化及肝竇擴(kuò)張的內(nèi)皮細(xì)胞、增生的血管內(nèi)皮細(xì)胞及其肌纖維帶則示強(qiáng)陽(yáng)性表達(dá)(圖4);D:異型增生的肝細(xì)胞示陽(yáng)性表達(dá)(圖7)。

圖1 肝細(xì)胞CaP強(qiáng)陽(yáng)性(SP×400)
圖2 膽管細(xì)胞CaP陽(yáng)性(SP×400)

圖3 肝竇及血管內(nèi)細(xì)胞CaP表達(dá)(SP×400)

圖4 肌纖維束CaP表達(dá)(SP×400)
圖5 毛細(xì)血管增生區(qū)CaP表達(dá)(SP×400)

圖6 貯脂及成肌纖維細(xì)胞CaP表達(dá)(SP×400)
圖7 異型增生肝細(xì)胞CaP表達(dá)(SP×400)
二、調(diào)寧蛋白表達(dá)與CHB肝內(nèi)病變的關(guān)系 本組資料提示,CaP表達(dá)與CHB肝內(nèi)病變的輕重程度有關(guān):在CHB輕度組,少數(shù)病例(24%)肝組織CaP表達(dá),與對(duì)照組相似,且強(qiáng)陽(yáng)性表達(dá)少見(6%);而在CHB重度組,肝組織中CaP表達(dá)陽(yáng)性占33%,強(qiáng)陽(yáng)性占67%(P<0.01);在LC組,除假小葉內(nèi)的肝細(xì)胞CaP示彌漫性強(qiáng)陽(yáng)性表達(dá)外,纖維組織內(nèi)的小膽管上皮細(xì)胞、擴(kuò)張的毛細(xì)血管內(nèi)皮細(xì)胞、成肌纖維細(xì)胞及平滑肌纖維帶(圖4)皆表達(dá)上調(diào),與上述組間有極顯著性差異(P<0.01)。肝組織CaP表達(dá)及組織定位與CHB肝內(nèi)病變的關(guān)系見表1。
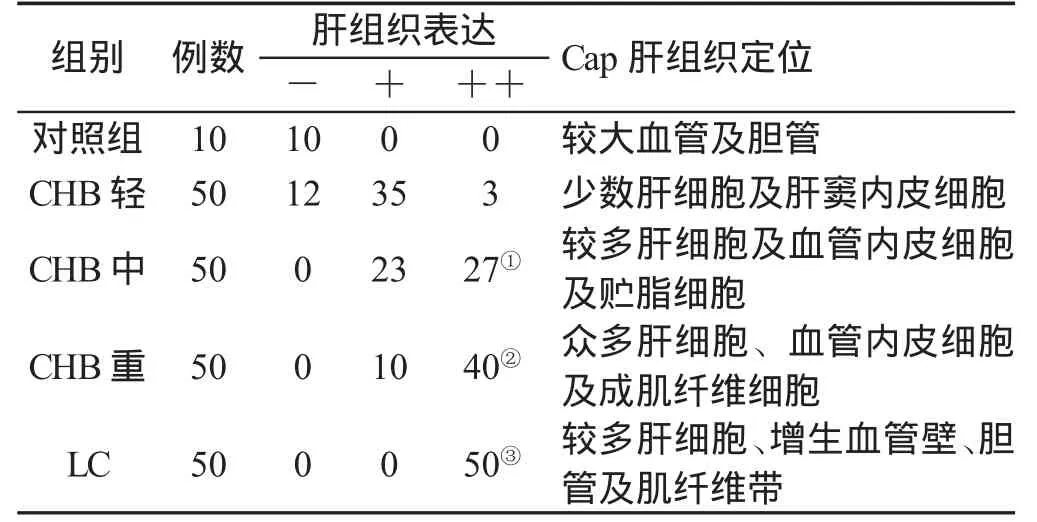
表1 CaP在肝組織的表達(dá)變化及定位
三、調(diào)寧蛋白表達(dá)與肝竇毛細(xì)血管化的關(guān)系 本組伴肝竇毛細(xì)血管化者161例(81%)。資料顯示,在CHB重度肝組織同時(shí)發(fā)生肝竇閉塞及擴(kuò)張改變,肝竇毛細(xì)血管化病變也較重,CaP表達(dá)范圍廣,顯色深;而在CHB輕度組,則肝細(xì)胞示濁腫,肝竇狹窄多見,肝竇擴(kuò)張罕見,肝竇毛細(xì)血管化也較輕,CaP表達(dá)范圍小,顯色弱,提示CaP表達(dá)變化與肝竇毛細(xì)血管化的輕重程度同步(表2)。
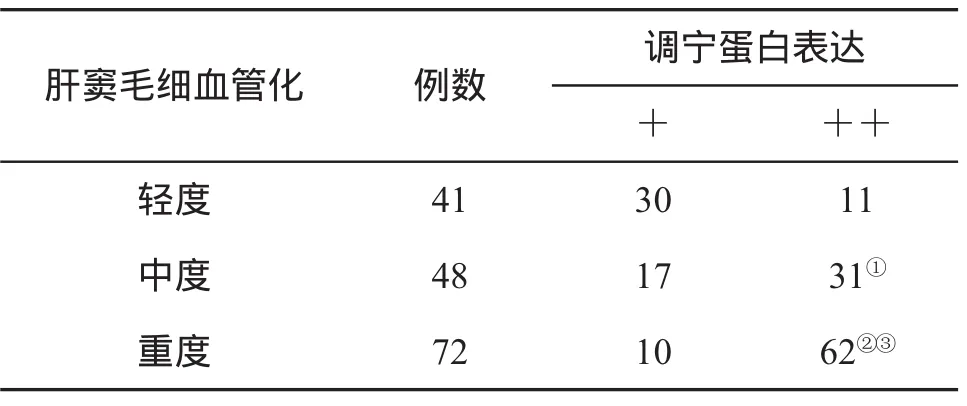
表2 調(diào)寧蛋白表達(dá)與CHB肝竇毛細(xì)血管化的關(guān)系
討 論
本研究發(fā)現(xiàn),①CaP在對(duì)照組肝組織定位于血管及膽管,其余組織無(wú)表達(dá);②在CHB肝組織,則肝細(xì)胞、貯脂細(xì)胞、成肌纖維細(xì)胞、肝竇內(nèi)皮及毛細(xì)血管化的內(nèi)皮細(xì)胞、小膽管上皮細(xì)胞、平滑肌纖維帶等組織均有CaP表達(dá);③CaP表達(dá)隨CHB肝組織的變性壞死、炎細(xì)胞浸潤(rùn)程度的加重而表達(dá)上調(diào);④伴隨肝竇擴(kuò)張、血管增生、肝竇毛細(xì)血管化及纖維化,CaP強(qiáng)陽(yáng)性表達(dá)率增加,提示CaP表達(dá)調(diào)節(jié)受CHB肝臟微循環(huán)障礙狀態(tài)的影響。
CaP主要生理功能是調(diào)節(jié)平滑肌的收縮和舒張[5,6]。正常生理情況下,平滑肌受到外界刺激時(shí),通過(guò)Ca2和磷脂激活蛋白激酶C,導(dǎo)致CaP磷酸化,不再抑制肌球蛋白ATP酶活性,平滑肌收縮;反之則平滑肌舒張[7,8]。本研究發(fā)現(xiàn),CaP表達(dá)變化與CHB肝微循環(huán)障礙有關(guān)。資料顯示,在肝內(nèi)病變較輕時(shí),匯管區(qū)血管炎癥雖較明顯,但血管破壞及阻塞的范圍較局限,肝界板完整或輕微破壞,肝細(xì)胞水腫及肝竇狹窄是局限的,中央靜脈完整,表明其有局限性血供輸入不良。而在肝內(nèi)病變較重時(shí),不僅肝實(shí)質(zhì)伴隨顯著的血管增生及肝竇毛細(xì)血管化,而且這些增生的血管可形成匯管與匯管、匯管與中央靜脈甚至跨小葉連通,即構(gòu)成所謂的橋形壞死或纖維化,導(dǎo)致肝小葉嚴(yán)重紊亂,在LC組肝細(xì)胞被血管間隔分隔為大小不同的團(tuán)塊(假小葉),因其無(wú)中央靜脈,加之肝細(xì)胞水腫引起肝竇閉塞,故導(dǎo)致肝小葉血液輸入和輸出障礙。在這樣的病理狀況下,可能反饋地引起CaP合成分泌增加,也可能引起Ca2+的變化,以改變ATP酶活性方式使CaP磷酸化或脫磷酸化,加速肝組織血液的輸入和輸出,以調(diào)整不利的病理環(huán)境。
本資料證實(shí),CaP在CHB肝組織合成分泌是增加的。CaP分泌上調(diào)應(yīng)增強(qiáng)血管的收縮[9,10],這與CHB輕度組肝竇閉鎖,毛細(xì)血管化的血管多閉合較一致。然而,在本組肝病重度組,CaP在肝組織表達(dá)雖然增加,但多數(shù)肝竇及血管則示擴(kuò)張改變。我們認(rèn)為,肝竇及血管擴(kuò)張表明肝內(nèi)壓力增加,系門脈高壓所致,而CaP除了可啟動(dòng)肌球蛋白輕鏈磷酸化,促進(jìn)肌動(dòng)蛋白與肌球蛋白結(jié)合及ATP酶活性,增加肌絲滑行及平滑肌收縮外[11,12],尚有抑制ATP酶活性,減少肌動(dòng)蛋白運(yùn)動(dòng)的功能,從而使平滑肌舒張,在門脈高壓下而引起肝竇毛細(xì)血管擴(kuò)張。
[1]高炬,方邦江,朱培庭,等.調(diào)寧蛋白在膽固醇結(jié)石形成中的表達(dá)及養(yǎng)肝柔肝中藥的干預(yù)作用[J].上海中醫(yī)藥大學(xué)學(xué)報(bào),2008,22(3):55-57.
[2]曹禹,李智,封瑞.肺動(dòng)脈高壓大鼠肺動(dòng)脈平滑肌上Calponin和TGFβ1的變化[J].中國(guó)藥理學(xué)通報(bào),2007,23(2):277-278.
[3]SUGENOYA Y,YOSHIMURA A,YAMAMUA H,et al.smooth-muscle calponin in mesangial cells:regulation of expression and a role in suppressing glomerulonrphritis[J].J AM Soe Nephol,2002,13:322-331.
[4]中華醫(yī)學(xué)會(huì)傳染病與寄生蟲病學(xué)會(huì)和肝病學(xué)會(huì).病毒性肝炎防治方案[J]. 中華傳染病雜志.2001,19(1):56-58.
[5]YANG JX,F(xiàn)ENG XH,ZHANG Y,et al.Influence of trace amount calponin on smooth muscle myosin in different States[J].Acta Biochimica Biophysica Sinica,2004,36(8):529-536.
[6]HYUN-DONG JE,SAMUDRA SG,TODD DA,et al.Calponin is required for agonist-induced signal transduction-evidence from an antisense approach in ferret smooth muscle[J].J Physiol,2001,37(2):567-577.
[7]FERJANI I,F(xiàn)ATTOUM A,MACIVER SK,et al.A direct interaction with calponin inhibits the actin-nucleating activity of gelsolin[J].Biochem J,2006,36(4):461-468.
[8]YANG JX,WANG XM,TANG ZY,et al.The characterization of Ca2+-calmodulin independent phosphorylation of myosin light chains by a fragment from myosin light chain kinase[J].Acta Biochim Biophys Sin,2003,35(9):793-800.
[9]COIRALT C,BLANC FX,CHEMLA D,et al.Biomecheanics and bio-energetics of smooth muscle contraction.Relation to bronchal hyperreactivity[J].Rey Mal Respir,2000,17(2pt2):549-554.
[10]LIN Y,SUN HJ,DAI SF,et al.The bidirectional regulation of filamin on the ATPase activity of smooth muscle myosin[J].Chin Med Sci J,2000,15(3):162-164.
[11]YANG JX,WANG XM,TANG ZY,et al.The characterization of myosin light chain phosphorylation by the constitutively active fragment of MLCK[J].Chin Med Sce J,2003,18(4):206-212.
[12]ZHOU Z,YIN J,DOU Z,et al.The calponin homology domain of vav 1 associates with calmodulin and is prerequisite to T cell antigen receptor induced calcium release in Jurkat T Lymphocytes[J].J Biol Chem,2007,282(32):23737-23744.
