短發(fā)夾狀RNA基因沉默補體受體C5aR及抑制LPS誘導(dǎo)的腎臟上皮細胞凋亡①
2010-09-07 01:33:24艷李尹注增張偉杰華中科技大學(xué)同濟醫(yī)學(xué)院附屬同濟醫(yī)院器官移植研究所教育部衛(wèi)生部器官移植重點實驗室武漢430030
中國免疫學(xué)雜志 2010年2期
陳 棟 張 艷李 明 尹注增 陳 剛 張偉杰 陳 實 (華中科技大學(xué)同濟醫(yī)學(xué)院附屬同濟醫(yī)院器官移植研究所 教育部/衛(wèi)生部器官移植重點實驗室,武漢430030)
·移植免疫學(xué)·
短發(fā)夾狀RNA基因沉默補體受體C5aR及抑制LPS誘導(dǎo)的腎臟上皮細胞凋亡①
陳 棟 張 艷②③李 明 尹注增 陳 剛 張偉杰 陳 實 (華中科技大學(xué)同濟醫(yī)學(xué)院附屬同濟醫(yī)院器官移植研究所 教育部/衛(wèi)生部器官移植重點實驗室,武漢430030)
目的:探討短發(fā)夾狀RNA基因沉默補體受體C5aR及抑制LPS誘導(dǎo)的腎臟上皮細胞凋亡的作用效果。方法:構(gòu)建針對大鼠補體受體C5aR基因編碼區(qū)的短發(fā)夾狀RNA(shRNA)真核表達載體質(zhì)粒pRNAT-U6.1-C5aR shRNA,采用電穿孔的方法轉(zhuǎn)染RK3E細胞,經(jīng)G418篩選后,形成穩(wěn)定的表達C5aR shRNA的細胞系。實驗分為3組,①正常對照組:未轉(zhuǎn)染的RK3E細胞;②陰性對照組:轉(zhuǎn)染空載體pRNAT-U6.1的RK3E細胞系;③實驗組:轉(zhuǎn)染C5aR shRNA的RK3E細胞系。經(jīng)脂多糖(LPS)孵育12小時后,流式細胞儀檢測各組細胞凋亡率,RT-PCR檢測mRNA的表達,γ計數(shù)儀測定125I標(biāo)記的C5a與RK3E的結(jié)合情況。結(jié)果:與正常對照組和陰性對照組相比,實驗組的細胞凋亡率顯著降低(P<0.01),C5aR mRNA水平顯著降低(P< 0.01),125I標(biāo)記的C5a與RK3E結(jié)合活性顯著下降。結(jié)論:針對C5aR的特異性短發(fā)夾RNA可以明顯引起靶基因的沉默,進而抑制LPS誘導(dǎo)的腎臟上皮細胞凋亡的發(fā)生。
RNA干擾;C5aR;基因沉默;凋亡
腎臟上皮細胞凋亡是急性腎功能衰竭等病理生理過程的重要機制之一,細胞凋亡的發(fā)生與補體系統(tǒng)的激活有關(guān),其中C5aR是補體系統(tǒng)的重要組成部分,表達于腎臟上皮細胞表面,參與補體系統(tǒng)的激活并介導(dǎo)靶細胞的凋亡[1]。RNA干擾(RNAi)是近年來發(fā)現(xiàn)的一種高效、特異性阻斷mRNA表達而導(dǎo)致轉(zhuǎn)錄后基因沉默的現(xiàn)象,從而不能表達相應(yīng)的特異性蛋白質(zhì),最終影響其生物學(xué)功能。這一干擾過程是由與目的基因有序列同源性的雙鏈RNA引起的。在哺乳動物中發(fā)現(xiàn)21~23核苷酸(nt)的雙鏈RNA介導(dǎo)的RNAi現(xiàn)象,開啟了應(yīng)用siRNA(Small interfering RNA)的治療應(yīng)用之門[2,3]。在本研究中,我們構(gòu)建了針對大鼠C5aR特異性的發(fā)卡結(jié)構(gòu)的RNAi質(zhì)粒表達載體,并轉(zhuǎn)染大鼠腎臟上皮細胞RK3E,建立穩(wěn)定的轉(zhuǎn)染細胞系,研究其基因沉默及抑制LPS誘導(dǎo)細胞凋亡的作用效果,為進一步研究如何減輕急性腎功能衰竭等病理過程奠定基礎(chǔ)。
1 材料與方法
1.1 材料
1.1.1 RNAi表達載體 pRNAT-U6.1/Neo siRNA表達載體購自美國Genescript公司。
1.1.2 細胞系 大鼠腎臟上皮細胞RK3E細胞由華中科技大學(xué)同濟醫(yī)學(xué)院附屬同濟醫(yī)院器官移植研究所保存。
1.1.3 試劑 DMEM培養(yǎng)基購自Sigma公司,胎牛血清購自Sigma公司,LB培養(yǎng)基為Promega公司產(chǎn)品,Trizol試劑購自Stratagene公司,C5aR的擴增引物由Invitrogen公司合成。
1.2 方法
1.2.1 C5aR基因shRNA的設(shè)計及載體構(gòu)建 選用pRNAT-U6.1/Neo siRNA表達載體作為空載體,其攜帶有cGFP和U6的啟動子,可以表達形成shRNA。根據(jù)GenBank提供的大鼠C5aR基因序列,使用美國Ambion公司在線設(shè)計軟件,通過基因序列比對,挑選出一條長為21個堿基的特異性寡核苷酸序列(5′-TG AAACTCTTGCTGTCCCTGC-3′),合成C5aR基因的發(fā)夾樣兩端配對的 siRNA寡核苷酸(Small hairpin RNA)(5′-G ATCCCGTG AAACTCTTGC TGTCCCTGCTTG ATATCCGGCAGGG ACAGCAAG AGTTTCATTTTTTCCAA A-3′),同時合成互補鏈5′-AGCTTTTGG AAAAAATG A AACTCTTGCTGTCCCTGCTCTCTTG AAGCAGGG ACAGC AAG AGTTTCACGG-3′),shRNA的寡核苷酸序列包括靶序列的正義鏈、反義鏈和環(huán)形區(qū)域。經(jīng)退火形成互補雙鏈,用于連接pRNAT-U6.1載體。
HindⅢ和BamHⅠ雙酶切RNAT-U6.1空載體成線性,T4連接酶將攜帶有C5aR特異性的RNAi核苷酸序列的互補雙鏈連接進入表達載體,然后轉(zhuǎn)化大腸桿菌DH5α,挑取單克隆菌落,小量擴增,收集菌液,使用DNA小量提取試劑盒提取質(zhì)粒,經(jīng)測序正確后,采用氯化銫離心法大量提取質(zhì)粒。
1.2.2 細胞培養(yǎng) 大鼠RK3E細胞用含10%胎牛血清的DMEM培養(yǎng)基在5%CO2、37℃下培養(yǎng)傳代,接種于75 cm2的細胞培養(yǎng)瓶。
1.2.3 細胞轉(zhuǎn)染 采用電穿孔的方法進行細胞轉(zhuǎn)染。收集RK3E細胞,2×106個細胞混合40μg的shRNA,在170 V,25 ms的條件下,應(yīng)用Bio-Rad電穿孔儀進行電穿孔,熒光顯微鏡下可見轉(zhuǎn)染的RK3E細胞呈綠色熒光,經(jīng)過G418篩選,形成穩(wěn)定的攜帶C5aR shRNA的RK3E細胞系。
1.2.4 脂多糖(LPS)誘導(dǎo)RK3E細胞凋亡率的檢測
實驗分為3組:(1)正常對照組:為正常的RK3E細胞;(2)陰性對照組:轉(zhuǎn)染空載體pRNAT-U6.1的RK3E細胞系;(3)實驗組:轉(zhuǎn)染 C5aR shRNA的RK3E細胞。
分別將上述3組細胞接種于六孔板中。分別加入LPS 100 ng/ml誘導(dǎo)RK3E細胞凋亡。LPS孵育12小時后收集細胞,磷酸鹽緩沖液(PBS)洗滌,調(diào)整細胞濃度為1×106ml-1,70%冷乙醇固定,-20℃過夜。檢測前PBS洗滌1次,加入含RNA酶(終濃度為0.25 mg/ml)的碘化丙啶(終濃度為50μg/ml)染色液,室溫孵育30分鐘,用流式細胞儀(美國FACSort-BD公司產(chǎn)品,使用CellQuest軟件)檢測細胞凋亡率。
1.2.5 半定量RT-PCR檢測RK3E細胞C5aR的表達 經(jīng)LPS刺激后,收集上述3組細胞,用Trizol試劑提取細胞總RNA,用3μg RNA合成cDNA。PCR條件:95℃5分鐘,94℃30秒,58℃30秒,72℃30秒,30個循環(huán),72℃延伸10分鐘。C5aR引物:上游引物為5′-GG ACCCCATAAGTAACG ACAGC-3′,下游引物為5′-CCACACCACCAGGGCATTTC-3′;內(nèi)參照磷酸甘油醛脫氫酶(G APDH)引物:上游引物為5′-TG ATG ACATCAAG AAGGTGGTG AA-3′,下游引物為5′-TCCTTGG AGGCCATGTAGGCCAT-3′,PCR產(chǎn)物經(jīng)1.5%的瓊脂糖凝膠電泳分析。使用Chemilmager 5500軟件進行圖像分析,測出擴增產(chǎn)物條帶的吸光度(A)值,并與同一樣本G APDH的A值進行比值定量分析,以此比值表示C5aR mRNA的相對含量。
1.2.6125I-rrC5a與RK3E的結(jié)合試驗 上述3組細胞接種于六孔板,并置于0℃冰上,用2 ml Hanks液孵育0.5小時,PBS液洗滌2次后,RK3E細胞與125I標(biāo)記的重組大鼠C5a(rrC5a)的PBS緩沖液(0.1%的胎牛血清)孵育20分鐘,隨后用PBS液洗滌4次后, 1%SDS裂解RK3E細胞,0.1%乙基苯基聚乙二醇洗滌后,采用γ計數(shù)儀測定125I標(biāo)記的C5a與RK3E的結(jié)合情況,數(shù)值用cpm值表示。
2 結(jié)果
2.1 成功構(gòu)建針對大鼠C5aR的shRNA表達載體pRNAT-U6.1-C5aR shRNA HindⅢ和BamHⅠ雙酶切pRNAT-U6.1空載體成線性,T4連接酶將攜帶有C5aR特異性的RNAi核苷酸序列的互補雙鏈連接進入表達載體,經(jīng)測序鑒定,證明我們設(shè)計的寡核苷酸序列已經(jīng)成功退火,并連接至pRNAT-U6.1。
2.2 C5aR shRNA轉(zhuǎn)染的RK3E穩(wěn)定細胞系 電穿孔的方法將C5aR shRNA轉(zhuǎn)染 RK3E細胞,由于pRNAT-U6.1攜帶有cGFP基因,可以表達綠色熒光蛋白,因此,轉(zhuǎn)染的RK3E細胞顯示綠色熒光,經(jīng)G418篩選,形成穩(wěn)定表達C5aR shRNA的細胞系,該細胞系在熒光顯微鏡下可見遍布視野的綠色熒光,而未轉(zhuǎn)染的RK3E細胞無綠色熒光,只見黑色背景視野(見圖1和圖2)。
2.3 LPS誘導(dǎo)的RK3E細胞凋亡率 流式細胞儀檢測結(jié)果顯示,在LPS的刺激12小時后,轉(zhuǎn)染C5aR shRNA的RK3E細胞凋亡率為24.81%±2.87%,而未轉(zhuǎn)染的RK3E細胞和轉(zhuǎn)染空載體pRNAT-U6.1的RK3E細胞凋亡率分別為 78.32%±3.62%和76.46%±5.54%,與未轉(zhuǎn)染的RK3E細胞和轉(zhuǎn)染空載體pRNAT-U6.1的RK3E細胞相比,轉(zhuǎn)染C5aR shRNA的RK3E的細胞凋亡率顯著下降,差異均有顯著的統(tǒng)計學(xué)意義(P<0.01)。見圖3。

圖1 未轉(zhuǎn)染的RK3E細胞Fig.1 Untransfected RK3E cells
圖2 C5aR shRNA轉(zhuǎn)染RK3E細胞Fig.2 C5aR shRNA transfected RK3E cells
2.4 C5aR mRNA水平的表達 LPS刺激12小時后,收集3組細胞,進行RT-PCR檢測mRNA水平的表達,結(jié)果顯示:經(jīng)LPS刺激后,轉(zhuǎn)染C5aR shRNA的RK3E細胞的mRNA相對含量為26.11%±3.16%,而未轉(zhuǎn)染的RK3E細胞和轉(zhuǎn)染空載體pRNAT-U6.1的RK3E細胞中mRNA相對含量為98.74%±2.27%和105.21%±2.57%。與未轉(zhuǎn)染的RK3E細胞和轉(zhuǎn)染空載體pRNAT-U6.1的 RK3E細胞相比,轉(zhuǎn)染C5aR shRNA的RK3E細胞mRNA表達水平顯著降低,差異均具有統(tǒng)計學(xué)意義(P<0.01),見圖4A、B。
2.5 各組RK3E細胞125I-rrC5a的結(jié)合活性 LPS刺激的條件下測定各組RK3E細胞中C5a的結(jié)合活性,結(jié)果顯示:與正常對照組和陰性對照組相比,經(jīng)LPS刺激后,125I-rrC5a與轉(zhuǎn)染C5aR shRNA的RK3E細胞的結(jié)合活性顯著降低(P<0.05),見圖5。

圖3 LPS刺激后RK3E細胞凋亡率Fig.3 Apoptosis rate of RK3Eafter LPS stimulated
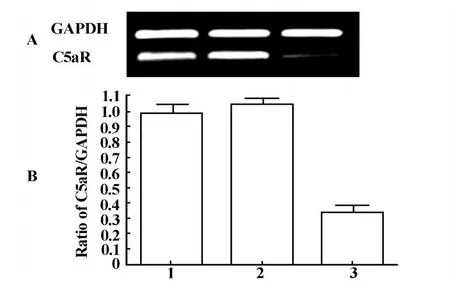
圖4 LPS刺激12小時后,RT-PCR檢測各組C5aR mRNA的表達(A,B)Fig.4 12 h after LPS stimulated,C5aR mRNA was detected by RT-PCR in different group(A,B)
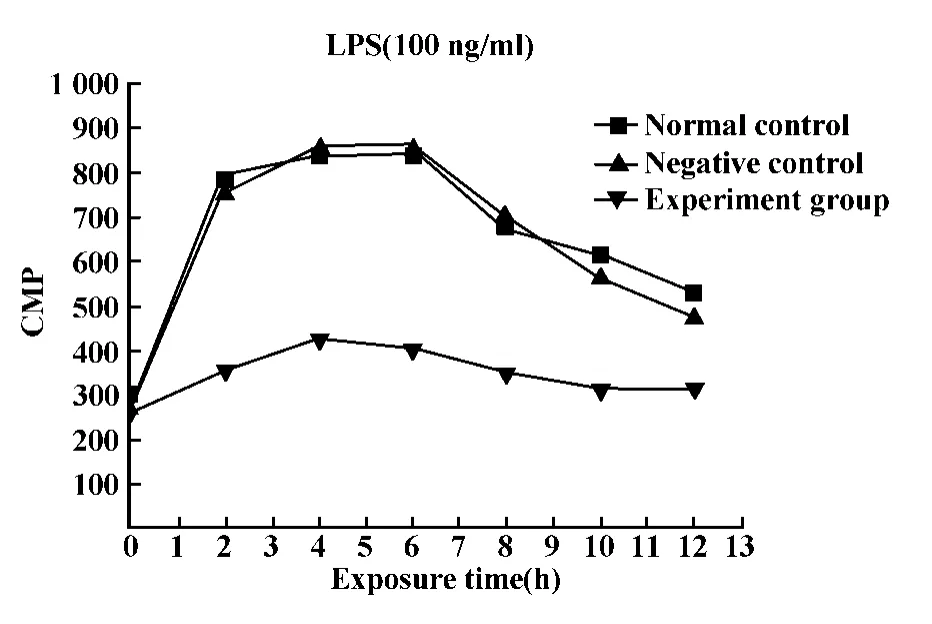
圖5 LPS刺激條件下125I-rrC5a與RK3E細胞的結(jié)合活性Fig.5 Binding of125I-rrC5a to RK3E stimulated with LPS
3 討論
補體活化參與缺血再灌注損傷等眾多病理生理過程,是關(guān)聯(lián)先天免疫和獲得性免疫的重要紐帶。腎臟上皮細胞表達眾多的補體受體C5aR,在腎臟上皮細胞的損傷和細胞凋亡的執(zhí)行過程中起關(guān)鍵作用[1,4,5]。因此,本研究中我們設(shè)計了針對大鼠補體受體C5aR作為抑制細胞凋亡的靶位點,進而探討通過短發(fā)夾狀RNA基因沉默C5aR及抑制LPS誘導(dǎo)的腎臟上皮細胞的凋亡的作用。
在多種生物體細胞中,外源或內(nèi)源性雙鏈RNA (Double-stranded RNA,dsRNA)可觸發(fā)細胞內(nèi)同源mRNA的特異性降解,從而使該基因表達沉默,這種現(xiàn)象稱為RNA干擾(RNA interference,RNAi)。RNAi屬于轉(zhuǎn)錄后水平的基因沉默機制,具有高度的序列特異性和抑制基因表達的高效性[6,7]。在本研究中我們根據(jù)大鼠C5aR的基因序列,設(shè)計發(fā)夾樣兩端配對的shRNA寡核苷酸,并連接到真核表達載體pRNAT-U6.1,構(gòu)建形成發(fā)夾狀siRNA的質(zhì)粒(shRNA)表達載體。然后進行電穿孔的方法轉(zhuǎn)染腎臟上皮細胞RK3E,經(jīng)G418篩選后得到穩(wěn)定的轉(zhuǎn)染細胞系,在熒光顯微鏡下可見遍布視野的綠色熒光,而未轉(zhuǎn)染的RK3E細胞只見黑色背景。該試驗表明,采用shRNA質(zhì)粒表達載體轉(zhuǎn)染哺乳動物細胞,就可以持續(xù)的轉(zhuǎn)錄出shRNA,通過篩選形成穩(wěn)定表達shRNA的細胞系,便于進一步研究基因沉默的作用效果。與siRNA介導(dǎo)的 RNAi作用相比,shRNA介導(dǎo)的RNAi作用能更長期,甚至穩(wěn)定地抑制目的基因的表達,更適合于對目的基因功能進行長期研究。
在腎臟C5aR主要表達在腎臟系膜細胞和近曲小管上皮細胞,遠曲小管和輸尿管、膀胱的移行上皮細胞也均有C5aR的表達。C5aR可能參與了腎臟上皮細胞多種細胞因子和補體成分的產(chǎn)生,誘導(dǎo)單核細胞釋放TNF-α、IL-1、IL-6和IL-8等細胞因子。在血液循環(huán)中較低的C5a濃度就可以誘導(dǎo)粒細胞應(yīng)答,而如果組織表達C5aR,則通過與C5a的結(jié)合,使組織局部的C5a濃度比血液循環(huán)要高很多,更有利于粒細胞向炎癥部位聚集,而且還可以促進內(nèi)皮細胞表達P選擇素,后者促進粒細胞粘附到血管壁,加速炎癥反應(yīng)[8,9]。給實驗動物注射脂多糖可以引起腎臟、肺和肝的C5aR mRNA的表達增多,同時C5aR的表達也上調(diào),說明其可能在組織炎癥反應(yīng)中扮演重要角色。本實驗中觀察到,經(jīng)LPS刺激12小時后,在未轉(zhuǎn)染的RK3E細胞、轉(zhuǎn)染空載體pRNAT-U6.1的RK3E細胞和轉(zhuǎn)染C5aR shRNA的RK3E細胞均出現(xiàn)C5aR mRNA及蛋白的表達,進而誘導(dǎo)腎臟上皮細胞出現(xiàn)凋亡。而與未轉(zhuǎn)染的RK3E細胞和轉(zhuǎn)染空載體的RK3E細胞相比,C5aR shRNA轉(zhuǎn)染RK3E細胞細胞凋亡率明顯下降,C5aR mRNA水平和蛋白活性顯著降低,該研究表明,一方面,LPS可以誘導(dǎo)腎臟上皮細胞RK3E出現(xiàn)凋亡,該細胞凋亡的發(fā)生與C5aR的表達增高有關(guān),在試驗中C5aR mRNA水平和蛋白均顯著升高。另一方面,C5aR shRNA對LPS誘導(dǎo)的RK3E細胞凋亡起到抑制作用,其主要機制是沉默RK3E細胞中的C5aR基因,試驗中C5aR mRNA水平較未轉(zhuǎn)染的RK3E細胞和轉(zhuǎn)染空載體的RK3E細胞顯著降低,說明了C5aR的基因沉默作用,C5aR基因沉默從而發(fā)揮抑制凋亡的作用。另外,我們在LPS的刺激性,測定了125I-rrC5a與未轉(zhuǎn)染的RK3E細胞、轉(zhuǎn)染空載體pRNAT-U6.1的RK3E細胞和轉(zhuǎn)染C5aR shRNA的RK3E細胞的結(jié)合活性,發(fā)現(xiàn)經(jīng)C5aR shRNA處理的RK3E細胞其結(jié)合活性顯著減低,提示經(jīng)RNA干擾處理的RK3E細胞表達C5aR顯著降低,進一步證實了采用RNA干擾技術(shù)可以有效的沉默腎臟上皮細胞表達C5aR。因此,我們設(shè)想,采用針對C5aR的shRNA來干預(yù)細胞凋亡過程,可以作為抑制腎臟缺血再灌注損傷等病理生理過程的一條有效途徑,并有可能為研制含有siRNA的藥物來抑制細胞凋亡的發(fā)生提供了思路[8,9]。
本文研究表明,通過攜帶C5aR的shRNA質(zhì)粒表達載體pRNAT-U6.1轉(zhuǎn)染哺乳動物細胞是可行的,經(jīng)過篩選可以形成穩(wěn)定的表達C5aR shRNA的細胞系,形成持續(xù)shRNA表達,C5aR shRNA可以顯著抑制靶基因達到基因沉默的效應(yīng)。LPS的刺激下, C5aR基因沉默可以明顯抑制腎臟上皮細胞的凋亡,該試驗為進一步研究抑制補體系統(tǒng)對腎臟缺血再灌注損傷的保護性作用等奠定條件。
1 Arumugam T V,Shiels I A,Strachan AJet al.A small molecule C5a receptor antagonist protects kidneys from ischemia/reperfusion injury in rats [J].K idney Int,2003;63(1):134-142.
2 張 艷,陳 棟,貢福盛 et al.RNAi介導(dǎo)Caspase-3基因及其抑制LPS誘導(dǎo)腎臟上皮細胞凋亡的研究[J].中國免疫學(xué)雜志,2008;24 (1):16-19.
3 Fire A,Xu S,Montgomery M Ket al.Pomnt and specific genetic interference by doublestranded RNA in Caenorhabditis elegans[J].Nature,1998; 391(6669):806-811.
4 de Vries B,K?hl J,Leclercq W ket al.Complement factor C5a mediates renal ischemia-reperfusion injury independent from neutrophils[J].J Immunol,2003;170(7):3883-3889.
5 Lewis A G,K?hl G,Ma Qet al.Pharmacological targeting of C5a receptors during organ preservation improves kidney graft survival[J].Clin Exp Immunol,2008;153(1):117-126.
6 Mao C P,Lin Y Y,Hung C Fet al.Immunological research using RNA interference technology[J].Immunology,2007;121(3):295-307.
7 杜 靜.RNA干擾的基礎(chǔ)研究和臨床應(yīng)用前景[J].國外醫(yī)學(xué)計劃生育/生殖健康分冊,2006;25(6):307-310.
8 Gueler F,Rong S,G winner Wet al.Complement 5a receptor inhibition improves renal allograft survival[J].J Am Soc Nephrol,2008;19(12): 2302-2312.
9 Welch T R,Frenzke M,Witte Det al.C5a is important in the tubulointerstitial component of experimental immune complex glomerulonephritis[J]. Clin Exp Immunol,2002;130(1):43-48.
[收稿2009-10-11]
(編輯 張曉舟)
Small hairpin RNA targeting rat C5aRcan inhibit the cell apoptosis induced by LPS in kidney epithelial cells
CHEN Dong,ZHANG Yan,LI Ming,YIN Zhu-Zeng,CHEN Gang,ZHANG Wei-Jie,CHEN Shi.Institute of Organ
Transplantation,Tongji Hospital,Tongji Medical College,Huazhong University of Science and Technology,Wuhan 430030,China
Objective:T o investigate RNA interference and apoptosis induced byLPS in kidney epithelial cells through silencing C5aR gene with small hairpin RNA(shRNA).Methods:We construct the eukaryotic expression vector of small hairpin RNA targeting rat C5aR gene, and transfected RK3E cell by electroporation,after G418 selection,so we got the stable cell line expressing C5aR shRNA.The experiment was designed into 3 groups:①normal control group,RK3E cells without transfection;②negative control group,RK3E cells transfected with blank vector;③experimental group,RK3E cells transfected with C5aR shRNA.After incubation withLPSfor 12 h,the ratio of apoptosiswas tested by flow cytometry,the level of mRNA was tested by RT-PCR,and bindingof125I-rrC5a to RK3E cells stimulated withLPSwere performed to examine the expression of C5aR in RK3E cells.Results:Compared with the normal control group and negative control group,in the experimental group the ratio of apoptosis was significantly decreased(P<0.01),and the expression of C5aR mRNA was significantly inhibited(P<0.01), and binding of125I-rrC5a to RK3E cells was significantly decreased also.Conclusion:Hairpin shRNA targeting C5aR gene can lead to obvious gene silence in vitro and inhibit the cell apoptosis induced by LPS in kidney epithelial cell.
RNA interference;C5aR;Gene silence;Apoptosis
R392.1 文獻標(biāo)識碼 A 文章編號 1000-484X(2010)02-0151-04
①本文為教育部留學(xué)回國人員科研啟動基金項目和國家自然科學(xué)基金(30600573)資助項目
②華中科技大學(xué)同濟醫(yī)學(xué)院附屬同濟醫(yī)院綜合醫(yī)療科,武漢430030
③通訊作者,E-mail:yzhang1977@yahoo.com.cn
陳 棟(1973年-),男,醫(yī)學(xué)博士,主要從事器官移植臨床及移植免疫方面的研究,E-mail:dchen0369@yahoo.com.cn。
