C57BL/6J小鼠脂肪組織總RNA提取及周脂素基因的克隆
2011-02-03 07:38:44衛兵艷劉田福郭永昌張引紅
中國比較醫學雜志 2011年8期
關鍵詞:實驗
衛兵艷,劉田福,郭永昌,張引紅
(山西醫科大學實驗動物中心,太原 030001)
脂肪組織不僅是機體的能量儲存庫,還是體內很大的內分泌器官;它可以合成或分泌多種具有廣泛生物活性的蛋白質或細胞因子,包括周脂素(perilipin)、脂聯素(adiponectin)、瘦素(leptin)、腫瘤壞死因子α(TNF-α)、白介素6(IL-6)[1-2]等這些因子與許多肥胖相關性疾病如胰島素抵抗、糖尿病、內皮功能障礙、高血壓病、動脈硬化癥等有關。體內能量代謝的核心反應是脂肪細胞的脂解。近年的研究表明[3],該反應主要由脂酶從胞液易位至細胞內脂滴的表面來調控的,該脂滴表面包被的蛋白中周脂素的磷酸化是HSL易位所必需的。由于周脂素在脂質分解中的重要作用,故成為近年來研究的熱點。
周脂素是一種新近在大鼠附睪脂肪組織中發現并命名的高磷酸化蛋白,特異表達于脂肪細胞和類固醇生成細胞脂質小滴表面[4]。新近研究表明,脂肪分解調控中的關鍵蛋白perilipin可能和2型糖尿病、肥胖、動脈粥樣硬化等多種代謝性疾病及心血管疾病有關[4-6]。但其作用機制尚不完全清楚。雖然周脂素的功能研究取得了一定進展,但關于周脂素的作用機制以及重組基因藥物產品開發等方面,還有待進一步研究。
以脂肪組織為材料提取高質量的總RNA主要存在兩方面的問題:第一,脂肪組織RNA含量較少,每毫克組織樣品所得總RNA的量小于0.05μg;第二,內源和外源性RNA酶對RNA的降解。因而所提RNA的得率較低。為獲得高質量的RNA制品,本實驗以脂肪組織為材料,經反復試驗,在商家提供的總RNA提取試劑的操作步驟基礎上優化和改進了某些提取過程,從而得到了符合實驗要求的高質量的總RNA。利用改進后的方法提取了C57BL/ 6J小鼠脂肪組織RNA。以此為模板進行RT-PCR,克隆了周脂素基因并對核酸序列進行分析,為下一步進行重組周脂素的大量表達及其生物學功能研究奠定基礎。
1 材料和方法
1.1 材料
1.1.1 實驗動物:C57BL/6J小鼠購于山西醫科大學實驗動物中心,生產許可證號:SCXK(晉2009-0001)。實驗過程中對動物處置符合2006年科學技術部發布的《關于善待實驗動物的指導性意見》[7]。
1.1.2 載體及宿主菌:受體菌E.coli JM109由本室保存;pMD18-T購自大連TaKaRa公司。
1.1.3 其他相關試劑:RNAisoTMPlus、限制性內切酶SpeⅠ和NotⅠ、DNA marker,均購于大連TaKaRa公司;質粒抽提試劑盒購于 O MEGA公司,LAmp二步法 R T-PCR試劑盒、DNA回收純化試劑盒、T4DNA連接酶、加A試劑盒均購于北京康為世紀生物科技有限公司。
1.2 方法
1.2.1 總RNA的提取:按照試劑盒說明書進行,部分操作步驟做相應的改進:(1)取新鮮的脂肪組織并將樣品的量加大到1 g,RNAisoTMPlus的量加大到20 m L。(2)第1次離心后上層含有大量油脂,用吸頭除去,保證完全去除油脂污染,然后再離心5 min。(3)進行兩次氯仿抽提,以去除DNA的污染。(4)用75%的乙醇洗RNA沉淀后,再用無水乙醇洗1次,縮短RNA沉淀晾干的時間,可避免RNA在空氣中暴露時間過長而增加外源RNase對RNA的降解。紫外分光光度計測定RNA的純度及含量,利用瓊脂糖凝膠電泳快速鑒定所提取的總RNA的質量。
1.2.2 引物設計、反轉錄-聚合酶鏈反應及其產物純化
1.2.2.1 引物設計與合成:根據GenBank中C57BL/6J小鼠周脂素的基因的序列,設計引物:序列如下(下劃線部分為酶切位點,斜體部分為保護性堿基。):5'端引物:5'-GGACTAGTCTTGGGCG TTTGCCTTACCT-3'(引入 S peⅠ酶切位點及保護性堿基),3'端引物:5'-AAGGAAAAAA GCGGCCGCCCTTGACGAGCAGCGACCTT-3'(引入NotⅠ酶切位點及保護性堿基)用于擴增周脂素全長編碼區。該引物由北京奧科公司合成。
1.2.2.2 RNA逆轉錄為cDNA:在0.2 m L的PCR管中加入下列成分:總 R NA(0.5μg/μL)4μL,Oligo d(T)18(100μmol/L)1μL,dNTP(2.5 mmol/L each)4μL,滅 R NA酶水4μL,混勻后置PCR儀中,65℃,5 min,迅速冰浴10 m in,短暫離心后繼續加入下列成分:5×RT buffer,5μL,RNA酶抑制劑(20 U/μL)1μL,0.1 mol/L DTT 2μL,混勻后置PCR儀中,37℃,孵育2 min后加入HiFi-MMLV逆轉錄酶 ( 200 U/μL)1μL,混勻后置PCR儀中,37℃50 min,終止反應,置 - 20℃冰箱保存備用。
1.2.2.3 周脂素基因的擴增:在0.2 mL的PCR管中加入下列成分:cDNA 4μL,10×LAmp PCR buffer 5μL,10μmol/L的正、反義引物各 2 μL,dNTP Mix(2.5 mmol/L each)4μL,5×RT-PCR enhancer 10μL,LAmp DNA polymerase 0.5μL加雙蒸水補足體積至50μL,混勻后置PCR儀中。PCR反應條件為:94℃預變性2 min,然后94℃變性30 s,60℃退火30 s,72℃延伸60 s,循環30次,72℃延伸5 m in,終止反應。取PCR擴增產物用1.2%瓊脂糖凝膠進行電泳,紫外燈下觀察電泳結果并照相。
1.2.2.4 PCR產物的純化回收:PCR擴增產物經1.2%瓊脂糖凝膠電泳分離,在紫外燈下切取含有目的基因的瓊脂糖凝膠塊,按膠回收試劑盒說明書中的方法,回收膠塊中的目的片段。
1.2.3 周脂素基因編碼區克隆、酶切及其DNA序列分析:
1.2.3.1 PCR產物加A:純化的目的基因7μL,dATP 1μL,10×A-tailing buffer 1μL,Tag DNA polymerase 1μL,混勻后置PCR儀中,72℃保溫30 m in。
1.2.3.2 PCR產物T-A克隆及鑒定:在10μL連接反應體系中,將加A的目的片斷與pMD18-T載體按摩爾數3∶1混合,加入T4DNA連接酶和連接緩沖液,22℃孵育1 h后,65℃10 m in滅活T4DNA連接酶,然后取上述連接產物10μL轉化大腸桿菌DH5α感受態細菌100μL,將轉化后的DH5α涂布于含氨芐青霉素(100μg/m L)及X-Gal(40 μg/m L)、IPTG(24μg/m L)的LB固體培養基平板上,37℃培養過夜,挑取單個白色菌落置于LB液體培養基中,擴大培養后提取質粒,經speⅠ/NotⅠ雙酶切鑒定含有目的基因的質粒由大連TaKaRa公司進行DNA測序,并將測序結果與GenBank中的已知基因序列及氨基酸序列進行比對。
1.2.3.3 C57BL/6J重組質粒的DNA序列分析:陽性克隆用通用引物測序,測序結果表明序列與GenBank比對達到100%一致(GenBank accession number:NM_175640.2)。
2 結果
2.1 C57BL/6J小鼠附睪脂肪組織總RNA的提取結果
提取的總RNA經紫外分光光度儀測定A值。A260/A280=1.89,比值在1.8~2.0之間;總RNA經1%瓊脂糖凝膠電泳可見28、18s 2條清晰的條帶,并且28s/18s≈2.0(圖1),表明提取的總RNA質量符合后續實驗的要求,可用于逆轉錄。

圖1 C57BL/6J脂肪組織總RNA瓊脂糖凝膠電泳Fig.1 Representative electrophoresis tracing of the total RNA from C57BL/6Jmouse adipose tissue
2.2 RT-PCR結果
提取C57BL/6J小鼠附睪脂肪組織的總RNA,經RT-PCR擴增后,1.2%瓊脂糖凝膠電泳可見與預期設計的周脂素基因片段大小一致的擴增條帶(包括兩側酶切位點和保護堿基)(圖2)。

圖2 反轉錄-聚合酶鏈反應產物瓊脂糖凝膠電泳Fig.2 Representative electrophoresis tracing of the RT-PCR products
2.3 PCR產物T-A克隆結果
經RT-PCR擴增的周脂素基因片段與pMD18-T載體連接構建成pMD18-T-plin重組載體,經speⅡNotⅠ雙酶切,1.2%瓊脂糖凝膠電泳可見2條帶,中1條帶與RT-PCR產物的電泳結果完全一致,另1條與pMD18-T載體片段(2 692 bp)的大小一致,見圖3。
2.4 周脂素重組質粒的DNA序列分析
陽性克隆用通用引物測序,測序結果表明序列與GenBank比對均為100%一致(GenBank accession number:D45371)。測序圖譜見圖4、5。表明pMD18-T載體中已含有周脂素基因片段。

圖3 周脂素重組質粒酶切鑒定結果Fig.3 Electrophoresis of the recombinant plasm id perilipin using double enzyme digestion
3 討論
由于脂肪組織富含脂類物質,單位體積細胞數較少,且單位細胞RNA含量也相對較少,按照RNAisoTMPlus試劑說明書提取脂肪組織RNA效果并不理想,所以如何有效地除去脂類的污染和最大限度地提高產量是在提取脂肪組織RNA過程中不可忽視的問題。在脂肪組織RNA提取過程中,對其中一些步驟作了適當的改進。本實驗主要做到以下幾點:①樣品量足,一般組織的RNA提取樣品量大概是100 mg,本實驗將樣品量加大到1 g進行提取從而保證了RNA的得率。②樣品組織裂解充分,短時間內液氮研磨至粉末狀加入超量的裂解液快速研磨至透明后劇烈震蕩3 min,保證了裂解液與組織的充分接觸。③除脂徹底,將樣品與裂解液反應后,離心5 min后,用吸頭除去了上層的脂類,取得的下層溶液應做同樣的2次離心以保證完全去除脂類污染。④去除內源和外源性RNA酶的影響,實驗過程中使用的實驗材料均用0.1%的DEPC水處理后,高壓滅菌,操作過程戴口罩帽子,盡量保持環境潔凈,液氮研磨充分,用75%的乙醇洗完RNA沉淀后,再用無水乙醇洗1次[8],以此減少內外源性RNA酶的影響。從而得到了高質量的RNA,為后續擴增周脂素基因做了充分保證。
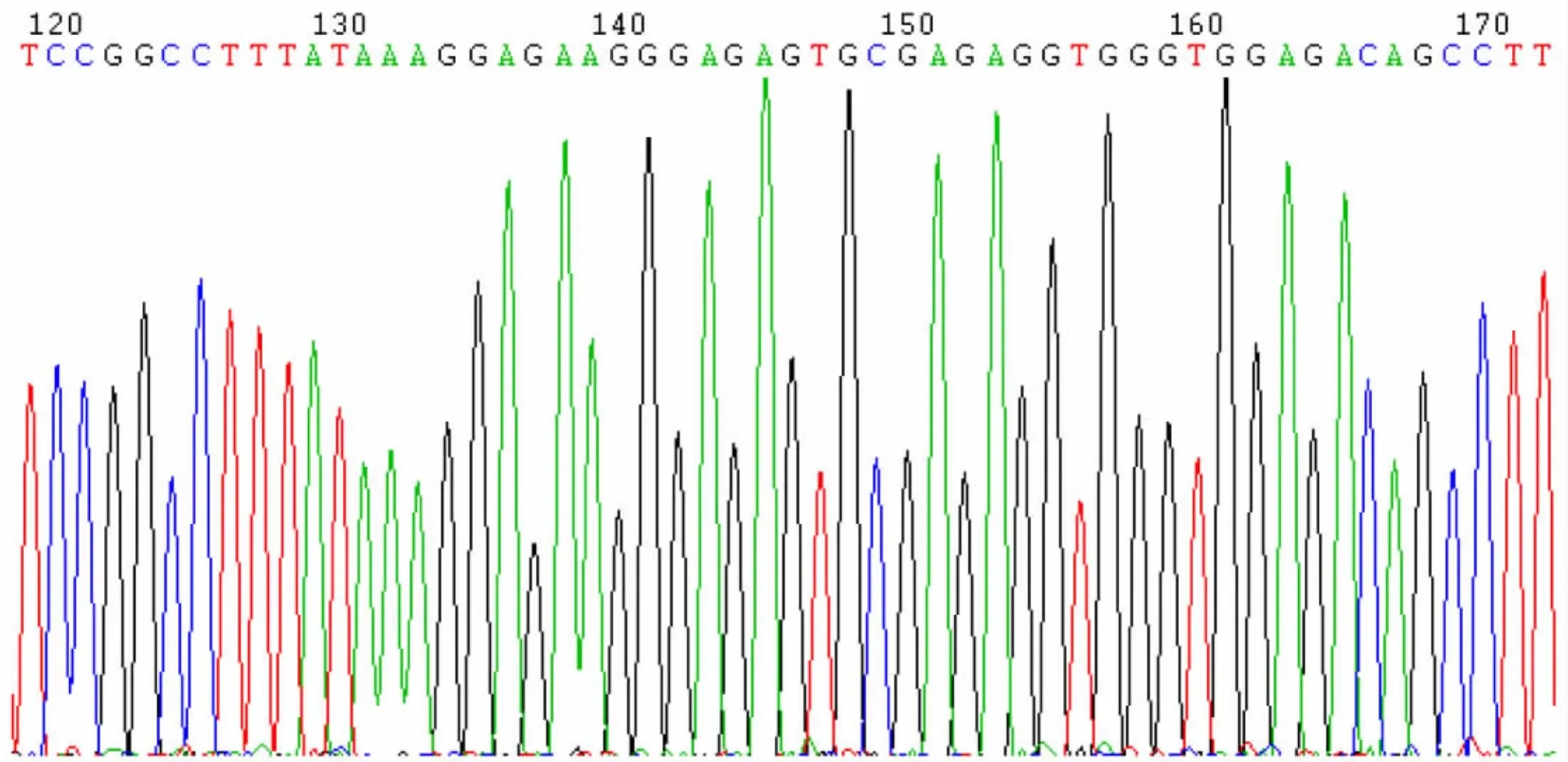
圖4 周脂素的測序圖譜Fig.4 Sequencing map of the plasm id perilipin

圖5 周脂素克隆序列比對部分結果Fig.5 Sequence analysis of the plasm id perilipin
周脂素由Plin基因編碼[9]。Perilipin氨基末端大約100個氨基酸的區域與ADRP的氨基末端區域極為相似,被稱為PAT結構域。鼠的perilipin基因定位于第7號染色體,人的基因定位于15q26,含有9個外顯子。研究發現,perilipin可能在脂肪分解調控中起到“分子開關”的作用,在脂肪水解和機體能量代謝過程中發揮著雙重作用[3],既可通過阻止脂肪酶接近脂滴降低基礎狀態下的脂解,又可促進激素刺激的脂肪分解,可以調控脂解率以及脂肪組織中甘油三酯的輸出。腫瘤壞死因子α(TNF-α)、過氧化物酶體增殖物激活受體γ(PPARγ)激動劑、瘦素(leptin)等均可以影響perilipin的表達[10-12]。由于周脂素與脂代謝的具體作用目前還不明確[13,14],其可能是聯系胰島素抵抗與代謝性疾病的重要環節,在脂質沉積及有序排列中發揮重要的調控作用[15-16],其在脂代謝中的關鍵作用可能與肥胖及相關疾病的發生有關[17]。
實驗經過提取脂肪總RNA、Rt-PCR、T-A克隆,得到周脂素編碼區重組質粒,測序結果表明,序列與Genbank比對均為100%一致(GenBank accession number:NM_175640.2)。本實驗得到沒有任何突變的基因,考慮與以下因素有關:①附睪脂肪組織總RNA的提取質量較高為進行反轉錄提供保證。②引物的GC比例設定合適(上游為55%,下游為60%),因而提高了基因擴增的特異性。③割膠純化時,操作速度很快,減少了紫外線對目的基因的損傷。紫外線可引起DNA損傷,應盡量減少其照射時間。④對純化的目的片段進行了加A反應,同時pMD18-T克隆載體是線性化的載體,其3’端有1個T伸出,顯著提高了重組效率。本實驗成功地克隆沒有任何突變的周脂素基因,為對其進行精確實驗提供了根本保障;為進一步真核表達周脂素蛋白,建立周脂素轉基因小鼠,分析周脂素的生物功能及作用機制提供了實驗基礎;為進一步理解體脂分布異常,胰島素抵抗開辟一新的方向,最終為探索該類藥物的開發提供嶄新的思路;可能也將為糖尿病等相關疾病的臨床工作提供一種新的防治策略。
[1]Crossno JT Jr,Majka SM,Grazia T,et al.Rosiglitazone promotes development of a novel adipocyte population from bone marrow-derived circulating progenitor cells[J].J Clin Inves,2006,16(12):3220-3228.
[2]Dalen KT,Dahl T,Holter E,et al.LSDP5 is a PAT p rotein specifically expressed in fatty acid oxidizing tissues[J].Biochim Biophys Acta,2007,1771(2):2102227.
[3]Moore HP,Silver RB,Mottillo EP,et al.Perilipin targets a novel pool of lipid droplets for lipolytic attack by hormonesensitive lipase[J].J Biol Chem,2005,280(52):43109 -43120.
[4]Greenberg AS,Egan JJ,Wek SA,et al.Perilipin,a major hormonally regulated adipocyte2 specific phosphoprotein associated with the periphery of lipid storage drop lets[J].J Biol Chem,1991,266(17):11341-11346.
[5]Souza SC,Vargas LM,Yamamoto MT,et a1.Overexpression of perilipin A and B blocks the ability of tumor necrosis factor alpha to increase lipolysis in 3T3-L1adipocytes[J].JBiol Chem,1998,273:24665-24669.
[6]Londos C,Sztalryd C,Tansey JT,et al.Role of PAT proteins in lipid metabolism.Biochimie,2005,87:45-49.
[7]中華人民共和國科學技術部.關于善待實驗動物的指導性意見[S].2006-09-30.
[8]曹廷兵,葉治家,鞏燕,等.一種快速鑒定RNA質量的方法[J].第三軍醫大學學報,2002,24(8):992-993.
[9]Martinez-Botas J,Anderson JB,TessierD,et al.Absence of perilipin Results in leanness and reverses obesity in Lepr(db/ db)mice[J].Nat Genet,2000,26:474-479.
[10]Fauny JD,Silber J,Zider A.Drosophila lipid storage droplet 2 gene(Lsd-2)is expressed and controls lipid storage in wing imaginal discs[J].Dev Dyn,2005,232(3):725-732.
[11]Londos C,Brasaem le DL,Schultz CJ,et al.On the control of lipolysis in adipocytes[J].Ann N Y Acad Sci,1999,892:155 -168.
[12]Arimura N,Horiba T,Imagana M,et al.The peroxisome proliferator 2 activated receptor gamma regulates expression of the perilipin gene in adipocytes[J].JBiol Chem,2004,279(11):10070-10076.
[13]Sztalryd C,Xu G,Dorward H,et al.Perilipin A is essential for the translocation of hormone-sensitive lipase during lipolytic activation[J].JCell Biol,2003,161:1093-1103.
[14]Clifford GM,Londos C,Fredric B.et al.Translocation of hormone-sensitive lipase and perilipin upon lipolytic stimulation of rat adipocytes[J].JBiol Chem,2000,275:5011-5015.
[15]Tansey JT,Sztalryd C,Gruia-Gray J,et al.Perilipin ablation Resultsin a lean mouse with aberrant adipocyte lipolysis,enhanced leptin production,and resistance to diet-induced obesity.Proc Natl Acad Sci U S A,2001,98(11):6494 -6499.
[16]Kawai T,Ng MC,Hayes MG,et al.Variation in the perilipin gene(PLIN)affects glucose and lipid metabolism in non-Hispanic white women with and without polycystic ovary syndrome.[J].Diabetes Res Clin Pract.2009,86(3):186 -92.
[17]Kern PA,Di Gregorio G,Lu T,et al.Perilipin expression in human adipose tissue is elevated with obesity[J].J Clin Endocrinol Metab,2004,89(3):1352-1358.
猜你喜歡
作文·小學低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學低年級(2024年2期)2024-04-29 00:00:00
作文·小學低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(2022年4期)2022-08-09 08:52:06
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
