基因Hath1表達對結腸癌細胞的抑制作用
2011-06-08 10:31:22趙麗麗劉忠張渝潔
中國醫藥生物技術 2011年3期
關鍵詞:結腸癌
趙麗麗,劉忠,張渝潔
Hath1 也稱作 ATOH1(atonal homolog 1),是一個堿性螺旋-環-螺旋結構(basic helix-loop-helix,bHLH)的轉錄因子。Hath1 基因與果蠅屬 atonal基因和鼠 Math1(mouse atonal homolog 1)基染色體同源,定位于人染色體 4q22,mRNA 長 1065 bp,編碼蛋白 38 kD[1]。結腸癌是常見的惡性腫瘤,發病率逐年上升,Hath1 基因能促進腸分泌細胞包括杯狀細胞(goblet)的終末分化,并誘導黏蛋白 2(mucin 2)的表達[2]。在結腸癌發生的漸進累積過程中,杯狀細胞逐漸減少甚至消失,黏蛋白也明顯減少[3]。在絕大多數結腸癌中 Wnt 信號一直處于活化狀態[4],使得 Hath1 的表達受到抑制。而Hath1 基因的表達可以促進結腸分泌細胞的分化,Hath1 基因表達的缺失限制了分泌型細胞的分化。因此嘗試在結腸癌中表達 Hath1 基因,研究 Hath1在結腸癌中是否可以發揮腫瘤抑制基因的作用。
1 材料和方法
1.1 材料
1.1.1 試劑 腫瘤細胞 HT-29、LS174T、SW480、SW620、IEC-6 購自上海生命科學院;Lipofectamine Reagent 轉染試劑盒購自美國 Invitrogen 公司;RT-PCR 試劑盒購自寶生物工程(大連)有限公司;MTS 購自美國 Promega 公司;trizol 購自上海生物工程公司;DNase I 購自大連寶生物公司;熒光染料超混合液購自美國 BioRad 公司;細胞 RNA抽提試劑盒購自上海華舜生物工程有限公司;pcDNA3.1(+) 大腸桿菌菌保(DH5 a)菌株由本公司保存。
1.1.2 儀器 FCAS440 流式細胞儀為美國 BD公司產品;Model 680 酶標儀為美國 BioRad 公司產品;7700 型實時熒光定量 PCR 儀為美國Applied Biosystems 公司產品。
1.2 方法
1.2.1 總 RNA 的提取 腫瘤細胞在 10cm 培養板中培養到 80%密度的時候,用 trizol 試劑提取細胞總 RNA。吸去細胞培養液,加入 3 ml trizol試劑,輕輕搖勻,室溫放置 5min 使細胞裂解完全,收集裂解液于離心管中,加 300 μl 氯仿劇烈搖勻,室溫放置 2min,4℃12 000×g離心 15min,取上層水相到另一離心管內,加入 750 μl 異丙醇搖勻,室溫放置 10min,4℃12 000×g離心 10min,棄去上清,用 1 ml 焦碳酸二乙酯(DEPC)–75%乙醇洗滌沉淀,晾干,溶解于 100 μl DEPC 水中。加11.3 μl 10×DNase I (脫氧核糖核酸酶)緩沖液,2 μl DNase I,42℃處理 25min,用分光光度計定量 RNA 濃度。組織總 RNA 的提取,在超聲破碎組織后以同法用 trizol(總 RNA 抽提試劑)試劑提取。
1.2.2 定量逆轉錄 PCR 法測 Hath1 的表達 取3 μg 總 RNA 于 PCR 管中,加 3 μl random primer p(dN)6(隨機引物)80 ng/μl,1.5 μl 10mmol/L dNTPs,加雙蒸水至 16 μl,于 PCR 儀中 65℃加熱 5min,冷卻至 4℃,加 6 μl 5×First Strand Buffer(合成第一鏈緩沖液),3 μl 50mmol/L MgCl2,3 μl 0.1 mol/L 二硫蘇糖醇(DTT),2 μl RNaseOUT(重組核糖核酸酶抑制劑),4℃5min,25℃2min,加 80 UnitsmmLV(莫洛尼鼠白血病病毒)逆轉錄酶,25℃10min,42℃50min 進行 cDNA 的第一鏈合成。75℃10min 失活逆轉錄酶,合成的cDNA 保存于 –20℃備用。
取 2 μl cDNA,加 10 μl 熒光染料超混合液,0.6 μl 10 μmol/L 引物,加水至 20 μl,在定量 PCR儀上檢測基因的表達水平。以 beta-actin 基因作為對照。引物序列如下:
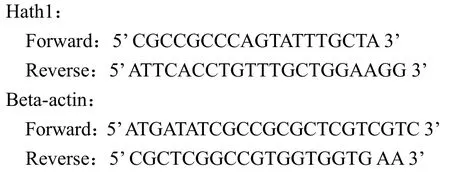
1.2.3 細胞培養及轉染 人腫瘤細胞 HT-29、LS174T、SW480、SW620 培養于 10%的胎牛血清的 RPMI 培養液中,在培養 48 h 后接種于6 孔板,按照 Lipofectamine 說明書進行脂質體轉染。
1.2.4 MTS 法測細胞增殖率 細胞增殖率用MTS 法測定。腫瘤細胞 HT-29、LS174T、SW480、SW620 在瞬時轉染 pcDNA3.1 或者 pcDNA3.1-Hath1 24 h 后,以每孔 1000 個細胞鋪于 96 孔板內,加 0.5%的血清于 37℃培養箱中培養 24 h。在測定前 1 h 加 MTS 到細胞中。具體方法參考試劑盒說明書。
1.2.5 IEC-6 穩定表達 Hath1 siRNA 細胞系的構建 RNA 干擾通過小發卡或短發卡 RNA(shRNA)實現。首先構建載有 Hath1 特異性 shRNA 的表達質粒載體 MSCV-LTRmiR30-PIG,將其轉染 LinX-1細胞包裝逆轉錄病毒載體,48 h 后收集含有逆轉錄病毒載體的細胞培養液,經 0.45 μm 的濾膜除去細胞及碎片,病毒凍存于 –80℃備用。
IEC-6 細胞以 200 μl 病毒液感染,感染的同時加 5 μg/ml 的聚凝胺(polybrene)提高病毒的感染效率。IEC-6 細胞以不含 shRNA 的病毒載體感染作為陰性對照。成功整合了病毒基因組的細胞會表達載體上的綠色熒光蛋白(GFP)。轉染了病毒的細胞用 2 μg/ml 的嘌呤霉素篩選 2 周。篩選后的具有藥物抗性的細胞采用實時熒光定量 PCR 和Western blot 方法分別從基因和蛋白質水平評價shRNA 對 Hath1 基因的干擾效應,并篩選出干擾效率最高的克隆。shRNA 的序列:AAACGACAAG AAGCTGTCCAAATAGTGAAGCCACAGATGTAT TTGGACAGCTTCTTGTCGTTG。
1.2.6 流式細胞儀檢測細胞凋亡百分率 細胞處理后,收集懸浮細胞于離心管,連同貼壁細胞視情況用胰酶消化后制成單細胞懸液同懸浮細胞合并。(700~1000)×g離心 5min,用 PBS 漂洗,重復離心,漂洗。用 70%乙醇固定,4℃過夜。離心,棄固定液,PBS 洗兩次并重懸細胞。加100 μl RNase A(0.5 mg/ml)至總濃度為50 μg/ml,37℃30min。再加入 400 μl 碘化丙啶(PI)染色液(100 μg/ml PI,1%Triton X-100,0.9%NaCl)至總濃度為50 μg/ml,混勻,避光保存 30min。上機測試,記錄激發波長 488 nm。以標準程序用FACScan 流式細胞儀檢測細胞內亞二倍體凋亡峰,使用 Mac V1.01 版本的 Cellquest 和 Modfit LT軟件分析。
1.3 統計學分析
應用 SPSS10.0 統計學軟件進行統計分析,兩組間比較采用組成設計t檢驗,P<0.05 為差異有統計學意義。
2 結果
2.1 Hath1 基因的表達
對 4株結腸癌細胞,一株大鼠小腸上皮細胞IEC-6 以及正常人小腸與結腸組織提取總 RNA,然后進行定量 PCR(圖1)。
結果顯示:在正常人的小腸與結腸組織中有著比較高水平的 Hath1 的表達,結腸中的表達比小腸高。此外,在大鼠小腸上皮細胞 IEC-6 中也有著較多 Hath1 的表達。而在 4株結腸癌細胞中Hath1 的表達都很低,尤其在轉移瘤 SW480 細胞中幾乎無表達。這些結果表明 Hath1 基因在腫瘤細胞中的表達低于正常組織,可能起著腫瘤抑制基因的作用。
2.2 Hath1 基因對結腸癌細胞增殖的影響
結腸癌細胞中的 Hath1 表達很低或是幾乎無表達,為研究 Hath1 對結腸癌細胞的作用,在 4株結腸癌腫瘤細胞中過表達人的 Hath1 基因,通過MTS 法測定細胞的增殖率(圖2)。
實驗結果表明,在所有 4株結腸癌細胞中Hath1 的表達都顯著地抑制了腫瘤細胞的增殖。在SW480 細胞中,細胞增殖率被抑制了 38.5%;在HT29 中細胞生長被抑制了 23.4%;而在 LS174T細胞中抑制率達到 55%。結果證明 Hath1 基因抑制了結腸癌細胞的增殖。
2.3 Hath1 基因的表達對結腸癌細胞凋亡的影響
為進一步闡明 Hath1 抑制結腸癌腫瘤細胞生長的機制,研究 Hath1 對結腸癌細胞 LS174T 的凋亡誘導作用(圖3)。實驗表明,LS174T 細胞過表達 Hath1 與對照組相比并無統計學差異。即Hath1 本身并不能有效地誘導腫瘤細胞的凋亡,其腫瘤生長抑制作用不是通過凋亡誘導實現的。但是,過表達 Hath1 的結腸癌細胞對化療藥物 5-FU引起的凋亡由對照組的 21.3%上升到 28.5%。即Hath1 的表達提高了腫瘤細胞對化療藥物的敏感性。
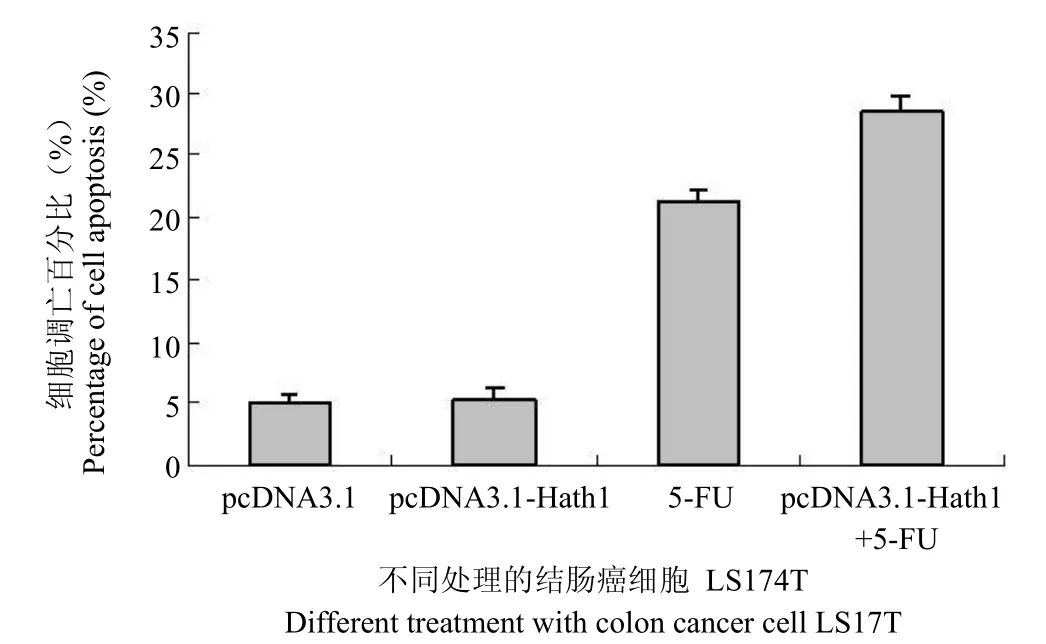
圖3 Hath1 基因對結腸癌細胞凋亡的影響Figure 3 The effect of Hath1 gene on the colon cancer cell apoptosis
2.4 正常細胞中下調 Hath1 的表達可提高細胞的生長率
在結腸癌細胞中的試驗表明提高 Hath1 的表達抑制了結腸癌細胞的生長。圖1 結果顯示正常細胞中有著較高水平的 Hath1 表達。針對大鼠中的 Hath1 同源基因設計的 siRNA 干擾試驗抑制大鼠 Hath1 同源基因的表達水平(圖4)。Hath1 基因被抑制的 IEC-6 細胞顯示出了顯著的細胞生長率的提高,從另一方面證明了 Hath1 起著腫瘤抑制基因的作用。
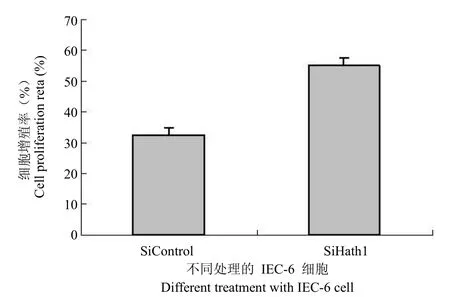
圖4 Hath1 siRNA對IEC-6細胞生長的影響Figure 4 The effect of Hath1 siRNA on the IEC-6 cell growth
3 討論
Hath1 基因作為一個轉錄因子,具有高度的同源性。Hath1 是 Ben-Arie 等[5]于 1996年克隆得到的果蠅 atonal 基因的同源基因。果蠅 atonal基因在果蠅神經系統的發育中起著關鍵的作用。Hath1 在小鼠中的同源基因 Math1 也在小鼠的消化系統的發育及分化過程中起著關鍵的作用。Math1缺失的轉基因小鼠在消化系統的發育中存在嚴重的缺陷。Math1 基因敲除的小鼠不能發育出所有三種小腸分泌型細胞[6],其中包括 goblet 細胞,但是吸收細胞 enterocytes 不受影響。Math1 與細胞增殖的標記 Ki-67 的共定位表明所有這些分泌型細胞都是來自一個表達 Math1 的前體細胞,Math1在分泌型細胞的發育中起著關鍵性的作用。其他研究也表明 Math1 基因在背部神經管的發育和細胞形態分化中有著重要的作用[7]。
所有這些研究都表明 Hath1 及其同源基因在個體的生長發育及多個組織的發生過程中起著重要的作用。其在小腸分泌型細胞的發生中起著的決定性作用已經得到了比較充分的研究,而在組織發育與細胞分化之外的作用一直沒有得到很好的研究。在結腸癌發生的漸進累積過程中,杯狀細胞逐漸減少[8],在結腸癌中,杯狀細胞明顯減少甚至消失。除發病率較低的黏液腺癌和印戒細胞癌外,大多數結腸癌中的黏蛋白都明顯減少[9-10],因此推測Hathl 基因與結腸癌的發病有關。
實驗闡明了這一在正常組織中高表達而在腫瘤細胞中低表達的因子具有對結腸癌細胞顯著的腫瘤抑制作用。過表達 Hath1 基因的結腸癌細胞都顯示出了顯著的生長抑制。在轉移性腫瘤 SW480中(38.5%)比原位性腫瘤 SW620 中(35.6%)的抑制作用略高。而在 LS174T 細胞中的抑制率達到了 55%。為此在 LS174T 細胞中進一步研究了這一抑制作用是否通過細胞凋亡發生。實驗結果表明Hath1 在LS174T 中不能有效地誘導腫瘤細胞發生凋亡,但是可以顯著地提高腫瘤細胞對化療藥物的敏感性。這為臨床研究提供了一些有意義的參考。
[1]Hutcheson DA, Vetter ML.The bHLH factors Xath5 and XneuroD can upregulate the expression of XBrn3d, a POU-homeodomain transcription factor.Dev Biol, 2001, 232(2):327-338.
[2]Vetter ML, Brown NL.The role of basic helix-loop-helix genes in vertebrate retinogenesis.Semin Cell Dev Bio1, 2001, 12(6):491-498.
[3]Boland CR, Montgomery CK, Kim YS.Alterations in human colonic mucin occurring with cellular differentiation and malignant transformation.Proc Natl Acad Sci U S A, 1982, 79(6):2051-2055.
[4]Bienz M, Clevers H.Linking colorectal cancer to Wnt signaling.Cell,2000, 103(2):311-320.
[5]Ben-Arie N, McCall AE, Berkman S, et al.Evolutionary conservation of sequence and expression of the bHLH protein Atonal suggests a conserved role in neurogenesis.Hum Mol Genet, 1996,5(9):1207-1216.
[6]Yang Q, Bermingham NA, Finegold MJ, et al.Requirement of Math1 for secretory cell lineage commitment in the mouse intestine.Science,2001, 294(5549):2155-2158.
[7]Gowan K, Helms AW, Hunsaker TL, et al.Crossinhibitory activities of Ngn1 and Math1 allow specification of distinct dorsal interneurons.Neuron, 2001, 31(2):219-232 .
[8]Velcich A, Yang W, Heyer J, et al.Colorectal cancer in mice genetically deficient in the mucin Muc2.Science, 2002, 295(5560):1726-1729.
[9]Yang W, Velcich A, Lozonschi I, et al.Inactivation of p21 WAFI/cip1 enhances intestinal tumor formation in Muc2-/-mice.Am J Pathol,2005, 166(4):1239-1246.
[10]Akazawa C, Ishibashi M, Shimizu C, et al.A mammalian helix-loop-helix factor structurally related to the product of Drosophila proneural gene atonal is a positive transcriptional regulator expressed in the developing nervous system.J Biol Chem, 1995,270(15):8730-8738.
猜你喜歡
天津醫科大學學報(2019年6期)2019-08-13 07:04:32
西南國防醫藥(2016年7期)2016-12-01 06:01:15
腹腔鏡外科雜志(2016年10期)2016-06-01 12:10:08
中國衛生標準管理(2015年6期)2016-01-14 05:17:12
中國衛生標準管理(2015年1期)2016-01-14 03:41:26
中國當代醫藥(2015年26期)2015-03-01 02:06:57
西南軍醫(2015年6期)2015-01-23 01:25:50
河南醫學研究(2014年3期)2014-02-27 14:51:48
沈陽醫學院學報(2014年1期)2014-02-16 06:19:24
中華介入放射學電子雜志(2014年1期)2014-02-02 05:24:06
