α、β-微管蛋白在乳腺癌變不同階段的表達及意義
2011-08-14 05:17:36應榮彪馮俊李建軍瞿海江姚俊
中國癌癥雜志 2011年8期
應榮彪 馮俊 李建軍 瞿海江 姚俊
臺州市腫瘤醫院,復旦大學附屬腫瘤醫院臺州分院腫瘤外科,浙江 臺州 317502
微管是細胞骨架的主要組成部分, 由α、β-微管蛋白異二聚體組成[1],是紫杉烷類抗腫瘤的作用靶點,微管蛋白可以通過對這一靶點的調節影響紫杉醇的抗腫瘤活性。微管在細胞分裂中具有極其重要的作用, 現已成為抗腫瘤藥物研究的重要靶點之一。目前,紫杉烷類是治療乳腺癌最重要的化療藥物之一,通過促進微管蛋白聚合,最終導致乳腺腫瘤細胞凋亡。然而紫杉烷類的耐藥性常常限制了治療效果,使病情難以長期控制。故揭示α、β-微管蛋白在乳腺癌變過程中表達水平的變化顯得尤為重要,為通過改變其表達水平改善乳腺癌紫杉烷類化療耐藥性提供理論基礎。
1 資料和方法
1.1 一般資料
選取臺州市腫瘤醫院2004年8月—2009年12月間具有完整的臨床病理資料和存檔的石蠟包埋組織標本,按世界衛生組織(WHO)乳腺病理組織學分類(2003年新版)標準[2],經雙盲病理診斷,乳腺非典型增生(atypical ductal hyperplasia,ADH)30例,從同期病例中隨機抽樣,導管內癌(ductal carcinoma in situ,DCIS)30例和浸潤性癌(invasive ductal carcinoma,IDC)30例,另從乳腺纖維腺瘤行區段切除術的標本中取瘤旁正常乳腺組織30例。
1.2 試劑與儀器
鼠抗人α-tubulin單克隆抗體IgG(ZM-0438),鼠抗人β-tubulin單克隆抗體IgG(sc-58880)為Santa Cruz公司產品;超敏即用型鼠二步法(非生物素)免疫組織化學檢測試劑盒(PV-9002)為美國GBI公司產品;使用全自動免疫組化機(美國Biogenex AS60302)進行免疫組化實驗。
免疫組織化學檢測:按照免疫組織化學二步法檢測試劑盒操作說明進行(一抗稀釋度:α-微管蛋白1∶100,β-微管蛋白1∶100)。以PBS液代替第一抗體做陰性對照,以已知乳腺癌切片做陽性對照。
1.3 免疫組織化學結果評價標準
α、β-微管蛋白染色陽性定義為:細胞質內呈均勻著色的棕黃色顆粒,背景清晰,無非特異性著色。染色結果采用雙盲法閱片判斷,依據Tanaka的定量計分法[3]:選取5個有代表性的不同區域,400倍鏡下計算陽性細胞百分率并評定顯色強度,依照陽性細胞百分率≤5%、>5%~25%、>25%~50%、>50%~75%和>75%分別得0、1、2、3、4分;依照顯色強度未著色、淺黃色、棕黃色、棕褐色分別得0、1、2、3分,兩項得分相乘,并取5個高倍視野得分的平均值。按照分數分為4個等級:0~1為(-),2~4為(+),5~8為(++),9分以上為(+++)。
1.4 統計學處理
本實驗免疫組化檢測所得結果為等級資料,采用SPSS 13.0統計分析軟件,以秩和檢驗、Spearman等級相關分析為統計學方法。P<0.05為差異有統計學意義。
2 結 果
2.1 α、β-微管蛋白表達和定位
可于光鏡下觀察到細胞質內呈均勻著色的棕黃色顆粒,細胞核、間質纖維細胞、淋巴細胞和單核吞噬細胞等均無陽性著色(圖1,2),各組樣本α、β-微管蛋白免疫組織化學染色評分情況(表1),正常乳腺組陽性率最低,ADH組、DCIS組至IDC組陽性率逐漸上升,其中IDC組最高。
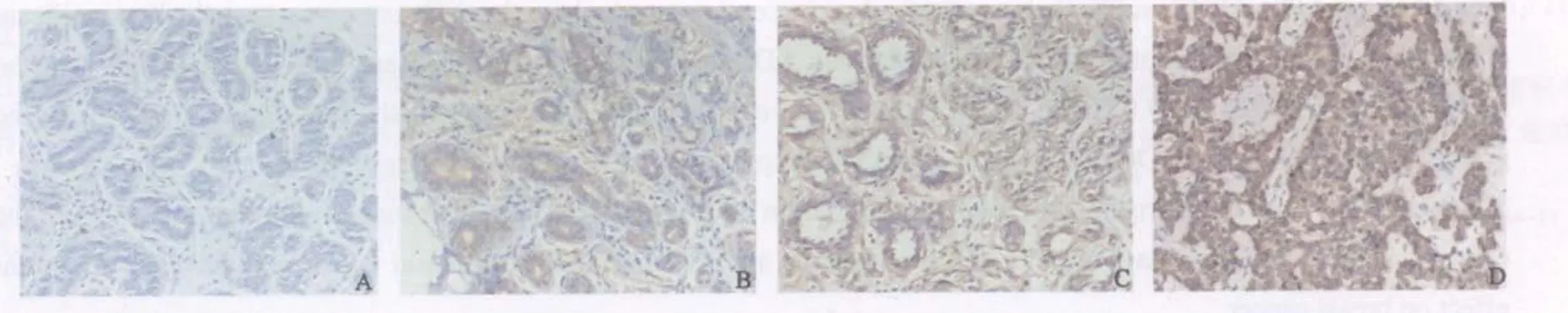
圖1 α-微管蛋白分別在正常組織(A)、ADH(B)、DCIS(C)及IDC(D)中的表達Fig.1 Expression of α-tubulin in normal(A), ADH(B), DCIS(C) and IDC(D) tissue of breast(PV, ×200)
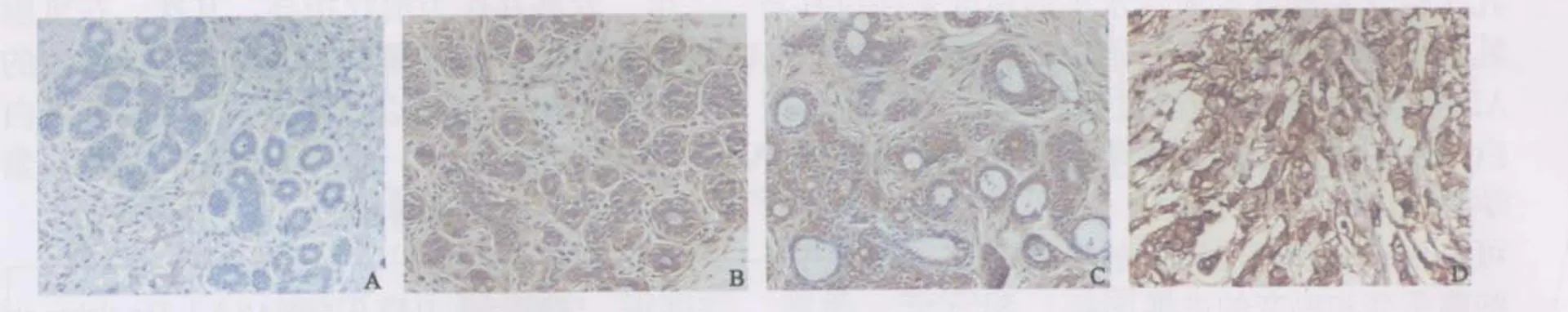
圖2 β-微管蛋白分別在正常組織(A)、ADH(B)、DCIS(C)及IDC(D)中的表達Fig.2 Expression of β-tubulin in normal(A), ADH(B), DCIS(C) and IDC(D) tissue of breast(PV, ×200)

表1 α、β-微管蛋白在不同乳腺組織中的表達情況Tab.1 Expression of α, β-tubulin in different breast tissues
2.2 α、β-微管蛋白表達的組間相關性
α-微管蛋白在乳腺正常組織、ADH、DCIS及IDC病例中的表達經等級分組資料的秩和檢驗(Krauskal-Wallis檢驗),上述4組樣本間表達的差異有統計學意義(χ2=32.864,P=0.000),再通過Mann-Whitney檢驗對各組進行兩兩比較:各組間差異均有統計學意義(P<0.05,表2)。
β-微管蛋白在乳腺正常組織、ADH、DCIS及IDC病例中的表達差異有統計學意義(χ2=26.28,P=0.000),各組進行兩兩比較:除ADH組和DCIS組間、DCIS組和IDC組間差異無統計學意義(P>0.05),其他各組間差異均有統計學意義(P<0.05,表2)。
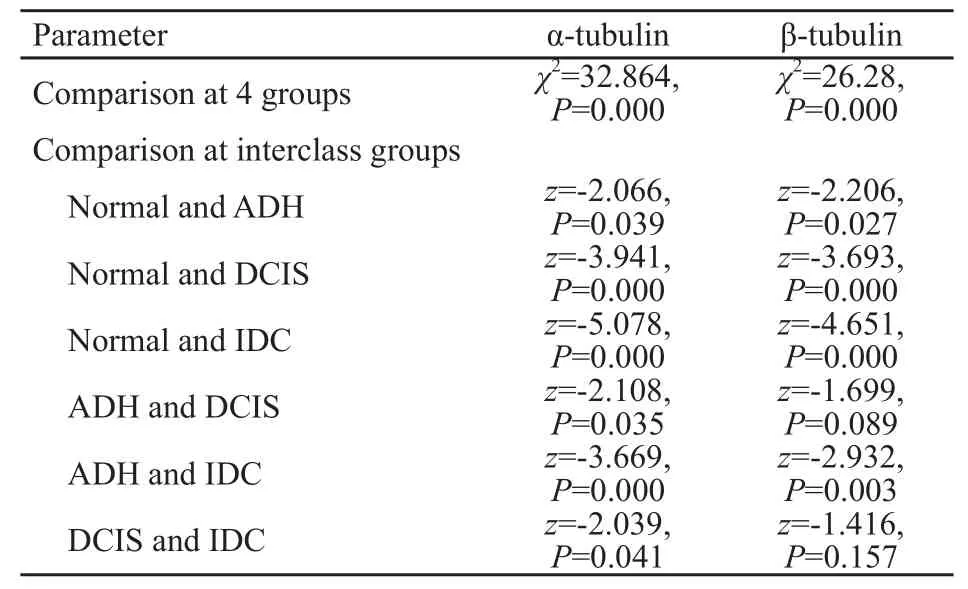
表2 不同乳腺組織間α、β-微管蛋白表達的比較Tab.2 Comparison of α, β-tubulin expression in different breast tissues
2.3 α、β-微管蛋白表達的組內相關性
分別在各組內對α、β-微管蛋白的表達進行Spearman相關性分析,結果顯示,在ADH、DCIS及IDC各組內,α、β-微管蛋白表達呈正相關(P<0.05),而正常乳腺組織則無相關性(表3)。
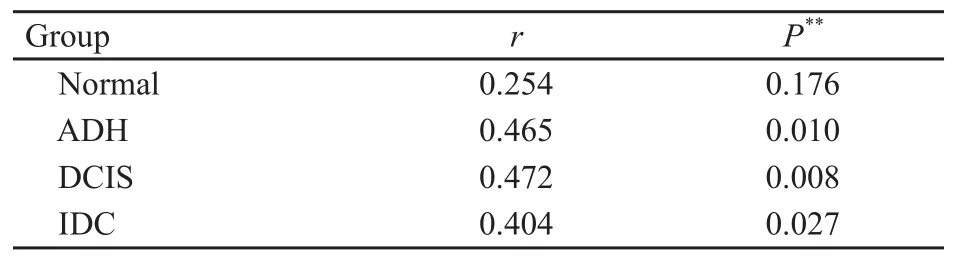
表3 α、β-微管蛋白在不同組織中表達的相關性Tab.3 Correlation of α, β-tubulin expression in different breast tissues
3 討 論
紫杉烷類是乳腺癌最重要的化療藥物之一[4],可通過促使α、β-微管蛋白異二聚體裝配成微管,并阻止其解聚而使微管束不能與微管組織中心相互連接,由此抑制紡錘體的正常形成,將細胞周期阻斷于G2/M期,導致有絲分裂異常或停止,使癌細胞無法繼續分裂而死亡[5]。最近的研究表明,乳腺癌患者對紫杉烷類耐藥性的出現,至少部分是由乳腺腫瘤細胞中α、β-微管蛋白的表達或修飾水平發生改變所引起的。
α、β-微管蛋白表達異常與腫瘤的惡性生物學行為密切相關。Giarnieri等[6]研究發現,α、β-微管蛋白的表達與直腸癌的發生、發展有關。本實驗從α、β-微管蛋白在乳腺癌變不同階段表達水平的角度來探討其與乳腺癌的關系。結果顯示,在乳腺正常細胞、ADH、DCIS及IDC發展過程中,α、β-微管蛋白陽性表達的細胞數逐漸上升,差異具有統計學意義(P<0.05)。提示α、β-微管蛋白的異常可出現在乳腺癌浸潤前的早期階段,并促進了細胞惡變和癌變的進展。
經進一步分析發現,在ADH及DCIS中,α、β-微管蛋白標志的陽性細胞數也明顯高于正常組織組,差異均有統計學意義。提示在ADH和DCIS中均已有α、β-微管蛋白的過度表達,可以推測在乳腺惡性腫瘤形成的關鍵性步驟中,隨著α、β-微管蛋白表達量逐漸增加,解聚動力加快,與微管蛋白池交換蛋白數量增加,引起中心體拷貝數增加,多紡錘體形成。中心體異常可導致染色體組的不穩定性和非整倍體的形成,導致抑癌基因的缺失或癌基因的獲得,改變細胞表型進而促進腫瘤發生、發展[7-8]。
本研究通過檢測乳腺癌細胞的中心體α、β-微管蛋白的改變,闡述了乳腺癌發展的不同階段組織中α、β-微管蛋白表達水平的變化,并對乳腺癌的發展模式進行了討論。這些研究成果從細胞遺傳學角度進一步闡明乳腺癌的發生、發展機制,提示α、β-微管蛋白可以作為乳腺癌腫瘤標志物,用于揭示乳腺癌發生、發展并作為治療指導。其次,在乳腺癌的治療過程中如果配合α、β-微管蛋白的表達水平檢測,并輔以針對α、β-微管蛋白表達調節的聯合治療,有可能提高乳腺癌對紫杉烷類的敏感性,改善預后。
[1] LEWIS S A, TIAN G, COWAN N J. The alpha- and betatubulin folding pathways[J].Trends Cell Biol, 1997, 7(12):479-484.
[2] TAVASSOLI F A, DEVILEE P. The WHO classification of tumors of the breast and female genital organs[M]. Lyon:IARC Press, 2003: 110-112.
[3] TANAKA K, IWAMOTO S, GON G, et al. Expression of survivin and its relationship to loss of apoptosis in breast carcinomas[J]. Clin Cancer Res, 2000, 6(1): 127-134.
[4] MCGROGAN B T, GILMARTIN B, CARNEY D N, et al.Taxanes, microtubules and chemoresistant breast cancer[J].Biochim Biophys Acta, 2008, 1785(2): 96-132.
[5] EL-KAREH A W, LABES R E, SECOMB T W. Cell cycle checkpoint models for cellular pharmacology of paclitaxel and platinum drugs[J]. AAPS J, 2008, 10(1): 15-34.
[6] GIARNIERI E, DE FRANCESCO G P, CARICO E, et al. Alpha- and beta-tubulin expression in rectal cancer development[J]. Anticancer Res, 2005, 25(5): 3237-3241.
[7] STORCHOVA Z, PELLMAN D. From polyploidy to aneuploidy, genome instability and cancer[J]. Nat Rev Mol Cell Biol, 2004, 5(1): 45-54.
[8] SALISBURY J L, D'ASSORO A B, LINGLE W L. Centrosome amplification and the origin of chromosomal instability in breast cancer[J]. J Mammary Gland Biol Neoplasia, 2004,9(3): 275-283.
猜你喜歡
中老年保健(2022年6期)2022-08-19 01:41:48
音樂探索(2022年2期)2022-05-30 21:01:37
現代臨床醫學(2022年1期)2022-02-12 02:04:58
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國生殖健康(2019年2期)2019-08-23 08:11:42
中國特種設備安全(2018年11期)2019-01-08 02:08:32
中國生殖健康(2019年6期)2019-01-06 09:20:12
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
祝您健康(2018年5期)2018-05-16 17:10:16
母子健康(2015年1期)2015-02-28 11:22:02
