典型草原不同圍封年限植被-土壤系統碳氮貯量的變化
2011-10-22 03:49:24敖伊敏焦燕徐柱
生態環境學報 2011年10期
關鍵詞:植物
敖伊敏,焦燕*,徐柱
1. 內蒙古師范大學化學與環境科學學院,內蒙古 呼和浩特 010022;2. 中國農業科學院草原研究所,內蒙古 呼和浩特 010010
在陸地生態系統中,土壤有機碳和氮在全球碳氮循環中起著重要作用[1-2]。研究顯示,土壤含碳量的多少很大程度上依賴于地表植被和土地利用狀況[3],且土壤有機碳含量的變化會影響植物對水分和營養元素的吸收,進而影響生產量[4]。就全球來看,土壤有機碳總儲量大約為1500 Pg[1],其中草原土壤的碳儲量大約占全球土壤碳儲量的30%[5],占草地生態系統碳總貯量的92%[6]。氮元素是調節陸地生態系統生產量、結構和功能的關鍵性元素[7],能夠限制群落初級和次級生產量[4]。草地生態系統作為我國乃至世界上最大的陸地生態系統,是陸地碳氮循環的一個重要部分[8-11],草地生態系統退化與恢復必然會直接影響到其本身碳氮貯量以及整個陸地生態系統的碳氮循環過程。典型草原是中國北方重要的生態屏障和牧民賴以生存的基本生產資料,在維持生態平衡、調節氣候、涵養水源、保持水土、防風固沙等方面起到重要作用,對維護生態安全,促進牧區經濟發展有著十分重要的意義。該類草原占內蒙古天然草地總面積的1/3(2163×107hm2),由于人口與家畜數量的劇增,目前典型草原面臨著生物多樣性減少,生產能力下降,草原大面積退化等嚴重問題,已使該區80%左右的草原處于不同程度的退化之中[12]。隨著國家對生態恢復與草地保護的關注,相關部門開展了大量的生態建設工作,其中對退化草地的恢復措施常用的方法有圍欄封育(自然恢復)、農業措施(包括松土、輕耙、淺耕翻、補播等)和建立人工草地等。其中圍封作為退化草地恢復治理的手段之一,可以通過自然力的作用使退化草地的植被與土壤得到良好的恢復,并使得退化草地出現了不同的恢復演替階段[13]。蘇永中等對科爾沁退化沙地草地研究表明,對已嚴重退化的草地進行禁牧圍封使植被得以恢復,輸入土壤的凋落物增加,使土壤有機碳得以積累[14]。閆玉春等內蒙古典型草原區研究表明羊草草原退化變型—冷蒿草原經過18年圍封,其植被和土壤特征得到明顯恢復[9]。Greenwood對威爾士南部冷溫帶草原研究表明,圍封2.5年后由于消除家畜的踐踏,表層土壤物理性質得到改善,相對于放牧樣地土壤非飽和水傳導率增加,土壤容重減小[15]。在埃塞俄比亞最北部的丘陵地帶(坡地)研究表明,圍封不僅可以有效地恢復植被,而且也能改善土壤養分,減少土壤侵蝕,草地土壤有機質、全氮均顯著高于放牧地[16]。自由放牧都會導致生態系統總碳氮貯量的降低,而圍封禁牧則可以明顯提高典型草原生態系統的總碳氮貯量[13-17]。
綜上所述,國內外學者對圍封草地恢復狀況的研究較多,但主要集中在植被特征的研究上,對碳氮貯量相關研究較少。已有的研究由于區域、氣候差異等原因使得研究結果也不盡相同。而在同一研究區域,相同氣候條件下,比較不同圍封年限的草地植物和土壤系統的碳氮貯量研究還較少。因此本文選取內蒙古典型草原生長季圍封恢復措施下的天然草地為研究對象,選取自由放牧草地作為對照,通過對生長季草地植被和土壤特征的測定,分析了在不同圍封復年限下植被和土壤碳氮貯量的變化,揭示圍封恢復措施對植物-土壤系統碳氮的平衡影響,為草地資源的合理利用與有效管理提供依據。
1 研究區域及研究方法
1.1 研究地概況
研究區域位于內蒙古錫林郭勒盟南部太仆寺旗 典 型 草 原 , 地 處 北 緯 41°35′~ 42°10′, 東 經114°51′~115°49′之間,氣候類型為中溫帶半干旱大陸性氣候,冬季寒冷干燥,夏季溫暖濕潤。年平均氣溫1.6 ℃,無霜期138 d,年平均降水407 mm,主要集中在6、7、8月,雨熱同期明顯,年平均日照2937.4 h。土壤為淡栗鈣土。草原類型為干旱與半干旱典型草原,以羊草(Leymus chinensis)、克氏針茅(Stipa krylovii)為優勢種,位于草群上層,下層植被為糙隱子草(Cleistogenes squarrosa)、洽草(Koeleria cristata)、冷蒿(Artemisia frigida)、星毛委陵菜(Potentilla acaulis),伴生種有根莖冰草(Agropyron michnoi)、寸草苔(Carex duriuscula)、扁蓿豆(Melilotoidesruthenica)、麻花頭(Serratula centauroides)、茵陳蒿(A. capillaris)、二裂葉委陵菜(P. bifurca)、阿爾泰狗娃花(Heteropappus altaicus)、矮韭(Allium anisopodium)、豬毛菜(Salsola collina)、歧花鳶尾(Iris dichotoma)等。在嚴重過牧地段植被群落演替為以冷蒿+星毛委陵菜為主,小半灌木冷蒿取代了羊草、克氏針茅,禾草類植物有所下降,常見伴生種無明顯變化[17]。
1.2 采樣及樣品處理方法
于2010年8月植物生長高峰期進行采樣。
1.2.1 植物樣品的采集
在每一樣地內隨機選取4個50 cm×50 cm的樣方,將植物齊地面刈割,65 ℃烘干,測定地上現存量干質量。用水洗分離法獲得根系樣品,65 ℃烘干,測定根系生物量干質量。然后將植物樣品和根系均粉碎過1 mm分樣篩,用于植物和根系有機碳和全氮的測定。
1.2.2 土壤樣品的采集
采用GPS定位S形布點法采集土壤樣品。為了避免地形等因素的影響,研究樣區選在平坦地形按照鄰近原則進行布置。每一圍封年限下的3塊樣地作為3個重復,每個樣地S形采樣,10個采樣點,每個采樣點用100 cm長土鉆分層采樣,分別采取0~10、10~20、20~30 cm土層樣,同時用環刀采集原狀土壤樣品用于測定土壤容重(圍封 25年樣地土壤容重未測)。土壤樣品帶回實驗室后風干,過1 mm篩,用于測定土壤化學性質。
1.3 樣品的測定與分析方法
采用環刀法測定土壤容重,用重鉻酸鉀氧化—外加熱法測定土壤及植物組分有機碳含量,用半微量凱氏法測定土壤全氮含量。
1.4 數據分析
用Excel進行數據處理并繪圖,通過SPSS 13.0軟件采用單因素方差分析(ANOVA) (P<0.05)。
2 結果與分析
2.1 圍封年限對植被碳氮貯量的影響
2.1.1 不同圍封年限草地植被生物量的變化
不同圍封年限之間草地地上植物現存量(F=141.11,P=0.000)存在顯著差異,根系生物量(F=2.45,P=0.114)差異性不顯著,但均隨圍封年限增加呈逐漸增加趨勢,最大值均出現在圍封 14年草地,之后有降低的趨勢(圖1)。與自由放牧草地相比,圍封 8、11、14、21和 25年樣地,地上植物現存量分別增加 5.7%、137.7%、293.7%、83.6%,和 121.5%;地下根系生物量分別增加41.1%、121.6%、171.4%、51.9%和124.3%。
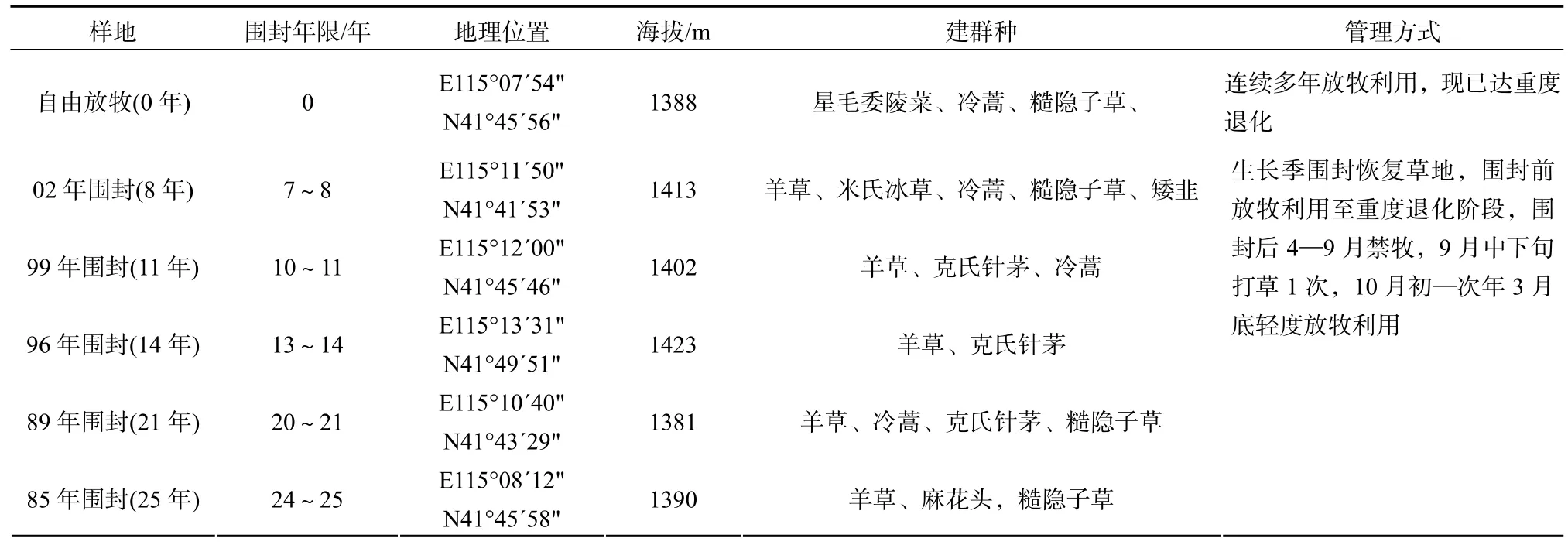
表1 研究樣地描述Table 1 Description of the experiment sites
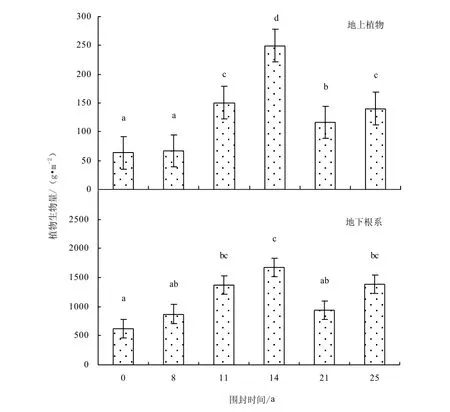
圖1 不同圍封年限草地植物生物量Fig.1 Plant biomass in different years of exclosure
2.1.2 不同圍封年限草地植被碳氮含量的變化
由圖2-3可知,不同圍封年限草地之間地上植物有機碳含量(F=3.51,P=0.033)和地下根系的有機碳含量(F=6.54,P=0.007)均存在顯著差異,植被的碳含量隨圍封年限的增加呈增加趨勢,在圍封14年達到最大值,之后隨圍封年限的繼續增加出現降低趨勢。氮含量隨圍封年限的增加呈降低趨勢,在圍封 14年樣地出現最小值,之后出現增加趨勢,與碳含量變化趨勢相反。不同圍封年限草地之間地上植物氮含量(F=22.66,P=0.000)和地下根系氮含量之間(F=6.78,P=0.006)均存在顯著差異。與自由放牧地相比,圍封8、11、14、21、25年地上植物碳質量分數增加6.7%、19.1%、33.1%、17.1%、17.3%,氮質量分數減少1.8%、14.2%、46.7%、2.5%、27.4%;地下根系碳質量分數增加11.1%、13.2%、17.1%、-10.1%、-3.9%,氮質量分數減少 23.2%、14.8%、42.3%、20.7%、4.7%。
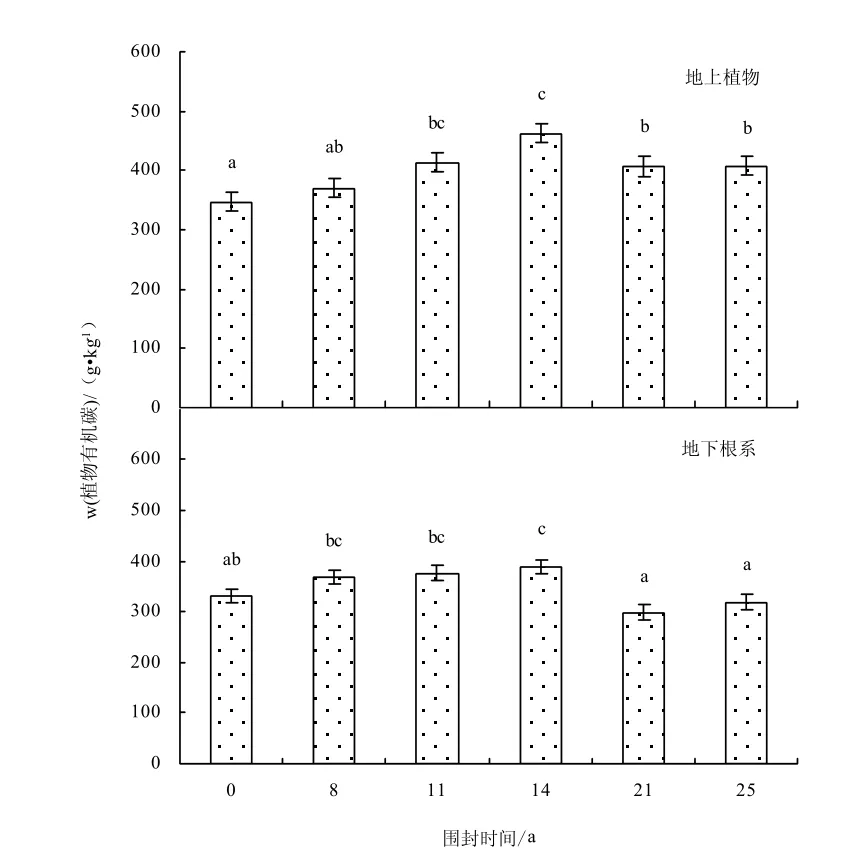
圖2 不同圍封年限草地植物有機碳含量Fig.2 Plant organic carbon content in different years of exclosure
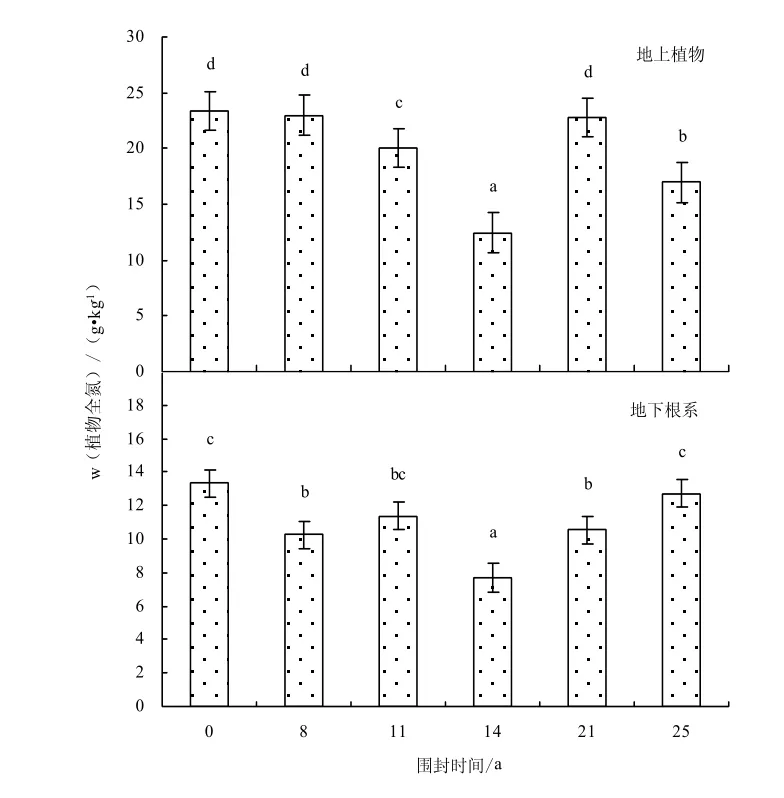
圖3 不同圍封年限草地植物全氮含量Fig.3 Plant total nitrogen content in different years of exclosure
2.1.3 不同恢復演替階段草地植被碳氮貯量的變化
由于地上植物和地下根系碳氮貯量是由其碳氮含量和生物量決定的,所以地下根系的碳氮貯量高于地上植物的碳氮貯量(如圖4-5)。地上植物碳貯量(F=145.07,P=0.000)和氮貯量(F=36.81,P=0.000)在不同圍封年限草地之間均存在顯著差異;而地下植物碳貯量(F=3.46,P=0.051)和氮貯量(F=2.87,P=0.080)在不同圍封年限草地之間不存在顯著性差異。植被的碳貯量隨圍封年限的增加而增加,最大值出現在圍封 14年樣地,其后出現降低趨勢;氮貯量也隨圍封年限的增加而增加,但其最大值出現在圍封 25年樣地。其原因是氮素作為植物的養分元素其流失速度要比碳大,所以恢復對氮素的累積時間要比碳累積多[13]。與自由放牧地相比,圍封8、11、14、21、25年地上植物碳貯量增加13.4%、183.5%、425.0%、115.1%、160.3%,氮貯量增加4.1%、103.9%、108.9%、78.9%、60.8%;地下根系碳貯量增加 56.7%、153.7%、215.6%、36.5%、114.4%,氮貯量增加11.9%、90.1%、52.9%、20.8%、112.9%。
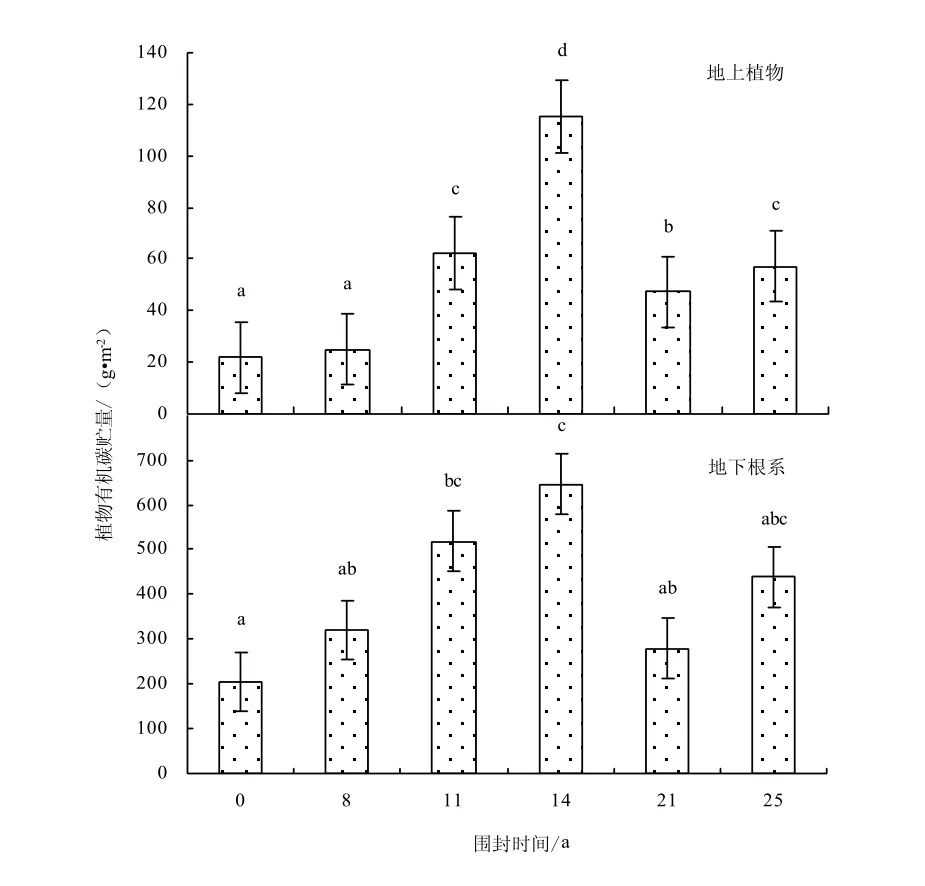
圖4 不同圍封年限草地植物有機碳貯量Fig.4 Plant organic carbon storage in different years of exclosure
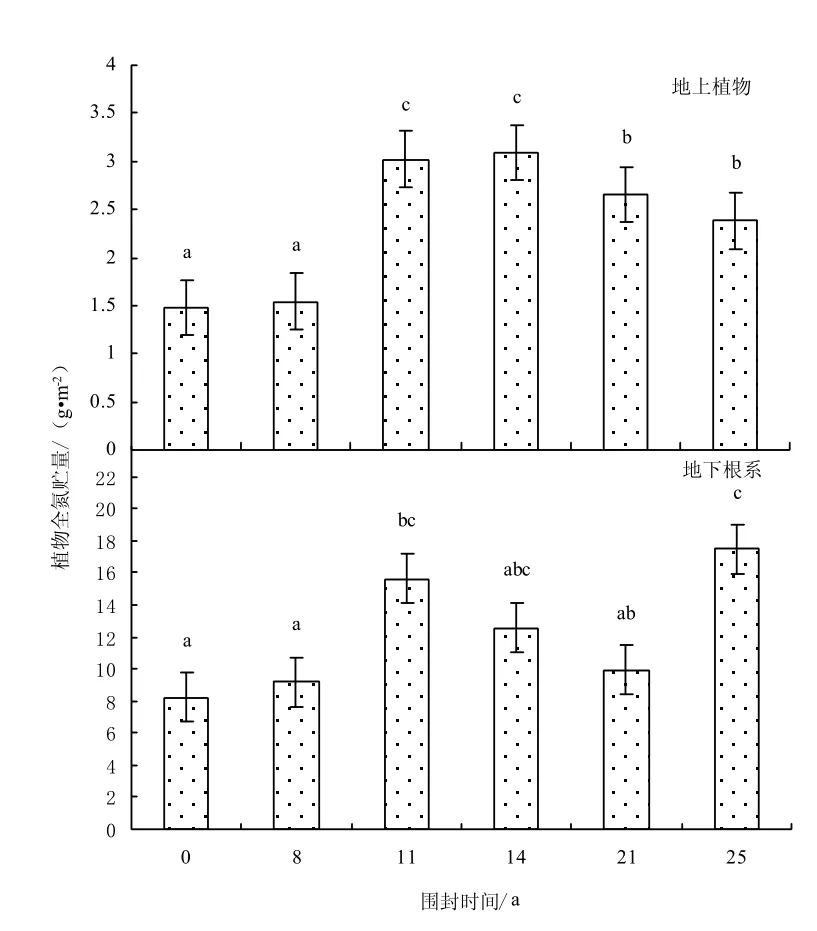
圖5 不同圍封年限草地植物全氮貯量Fig.5 Plant total nitrogen storage in different years of exclosure
2.2 圍封年限對土壤碳氮貯量的影響
2.2.1 不同圍封年限草地土壤容重的變化
不同圍封年限典型草原土壤容重的變異特征如圖6所示,與放牧草地相比,重度退化草地采用生長季圍封措施后土壤容重隨圍封年限的增加而下降,圍封 14年達到最低,之后又呈上升趨勢。就不同土層來看,同一樣地容重均隨土層加深而增加,原因為植物根系的土層分布不同[18]。土壤容重在不同圍封年限草地的不同土層間均具有顯著性差異(F=4.26,P=0.050)。與自由放牧地相比,圍封8、11、14年,0~10 cm土層土壤容重下降0.42%、2.36%、13.77%,圍封21年增加1.87%;10~20 cm土層圍封8、11、14、21年容重分別下降2.53%、8.95%、15.60%、7.61%;20~30 cm土層容重下降3.49%、6.91%、12.81%、10.71%。
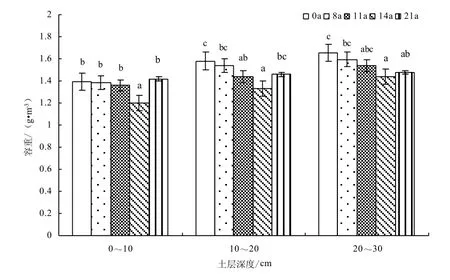
圖6 不同圍封年限草地土壤容重變化Fig.6 Changes of soil bulk density in different years of exclosure
2.2.2 不同圍封年限草地土壤有機碳和全氮含量變化
不同圍封年限典型草原土壤有機碳、全氮的變異特征如圖7-8所示,自由放牧草地采用生長季圍封措施后,圍封樣地土壤有機碳、全氮隨圍封年限的增加呈增加趨勢,圍封 14年達到最大值,隨圍封年限的繼續增加出現下降趨勢。不同圍封年限草地之間土壤有機碳含量(F=4.19,P=0.042)和土壤全氮含量(F=4.68,P=0.031)均存在顯著差異。與自由放牧草地相比,圍封8、11、14、21、25年,土壤 0~10 cm土層有機碳質量分數分別增加19.57%、23.57%、77.12%、21.70%、38.65%,全氮質量分數增加24.36%、37.11%、62.27%、30.18%、-10.32%;10~20 cm土層有機碳質量分數增加1.38%、18.83%、128.34%、43.52%、28.03%,全氮質量分數增加0.75%、31.50%、119.25%、120.07%、-18.32%;20~30 cm土層有機碳質量分數增加5.46%、6.27%、96.09%、22.32%、28.88%,全氮質量分數增加36.18%、70.55%、120.07%、90.65%、-36.32%;0~30 cm土層有機碳質量分數增加10.19%、17.43%、97.90%、28.52%、32.76%,全氮質量分數增加19.10%、42.20%、92.82%、53.48%、-18.31%。
2.2.3 不同圍封年限草地土壤碳氮貯量變化
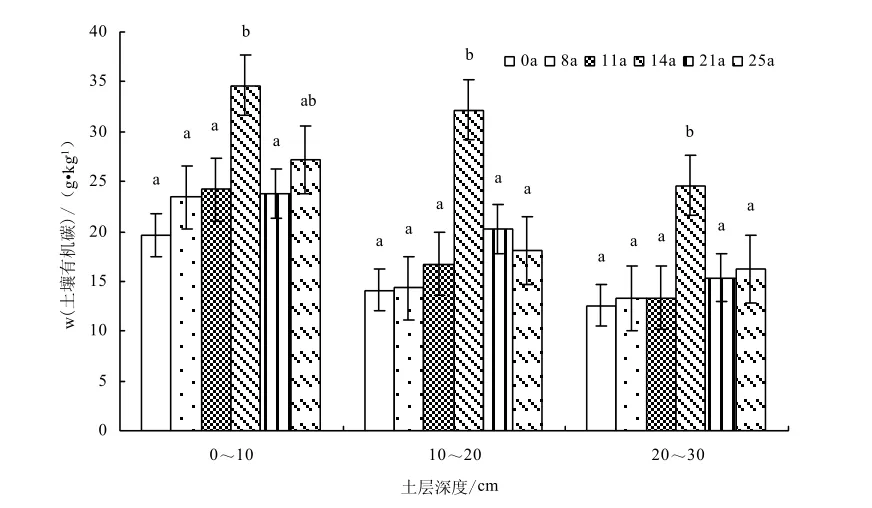
圖7 不同圍封年限草地土壤有機碳含量變化Fig.7 Changes of soil organic carbon content in different years of exclosure
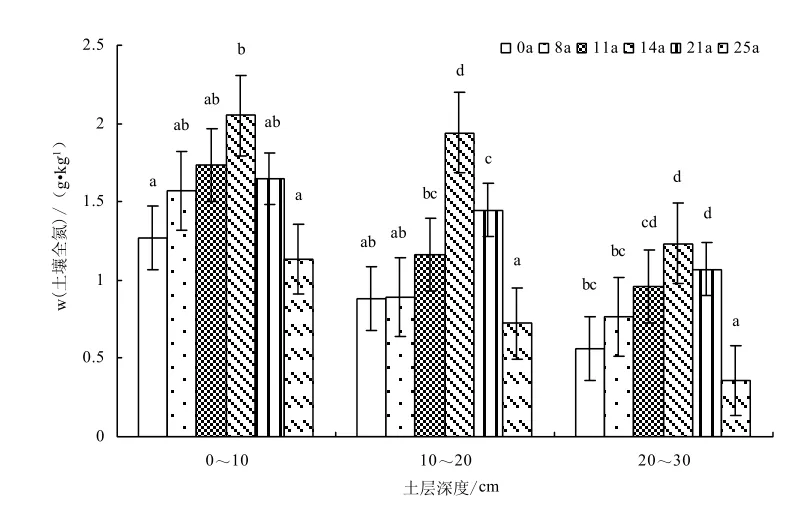
圖8 不同圍封年限草地土壤全氮含量變化Fig.8 Changes of soil total nitrogen in different years of exclosure
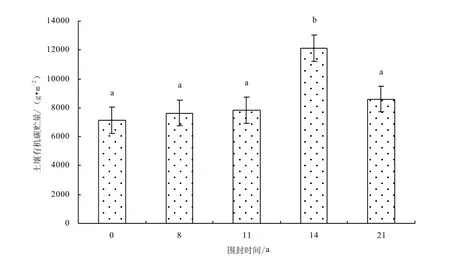
圖9 0~30cm土層不同圍封年限草地土壤有機碳貯量Fig.9 Soil organic carbon stroage in different years of exclosure in 0~30cm layer
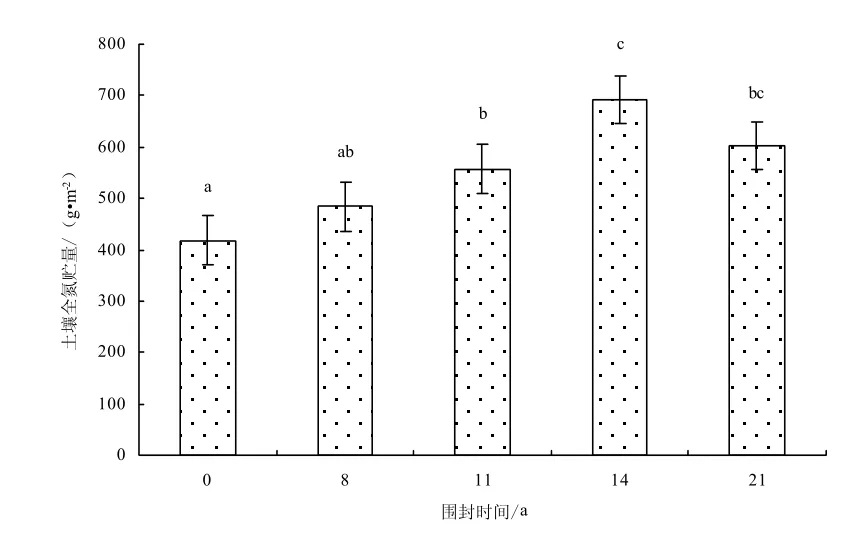
圖10 0~30cm土層不同圍封年限草地土壤全氮貯量Fig.10 Soil total nitrogen storage in different years of exclosure in 0~30cm layer
土壤碳氮貯量由土壤碳氮含量和土壤容重決定,因此土壤碳氮貯量與土壤碳氮含量呈現相同的變化趨勢:自由放牧樣地碳氮貯量最低,而季節性圍封恢復使退化草地土壤碳氮貯量提高,而且圍封時間越長土壤碳氮貯量越大,恢復14年達到最大,隨著圍封時間的繼續增加,土壤碳氮貯量有減小的趨勢(圖 9-10)。由于土壤容中的變化而縮小了差異,在0~30 cm土壤中,不同圍封年限草地之間土壤碳貯量(F=4.91,P=0.032)和氮貯量(F=5.55,P=0.027)差異性均顯著。與自由放牧地相比,圍封8、11、14、21年樣地0~30 cm土壤碳貯量分別增加 7.2%、9.8%、69.9%、20.7%,氮貯量分別增加15.7%、33.2%、65.4%、44.1%。各恢復樣地0~10 cm土壤碳貯量占0~30 cm土壤碳貯量的38.2%、41.4%、41.3%、34.2%、39.3%和氮貯量占42.2%、43.9%、42.0%、35.4%、38.8%。說明圍封措施對碳氮貯量的影響主要集中于土壤的表層,對深層土壤影響較小。
2.3 不同圍封年限草地植物-土壤系統碳氮貯量總量及其分配
本研究植物-土壤系統碳氮總量包括地上植物、地下根系和土壤碳氮貯量之和。處于不同圍封年限的草地植物-土壤(0~30 cm)系統碳氮總量比自由放牧地高,碳總量分別高出8.6%、14.3%、75.0%、21.4%和氮總量分別高出 15.5%、34.5%、65.4%、43.8%。在自由放牧地和圍封草地中,其碳氮貯量在植被—土壤系統中的分配比例相當,絕大部分都分配在土壤中,平均在96%左右,而地上植物組分碳氮貯量所占比例最小,在6個樣地均不足1%,植物組分中碳氮貯量主要集中在地下,圍封恢復草地和持續放牧草地0~30 cm根系碳氮貯量分別是其地上植物組分碳截存的5~12倍左右。由表2可知,圍封時間越長,地上植物和地下根系在碳氮總量中所占比重越大,土壤所占比例越來越小,其原因在于生長季圍封促進了草原植物的生長和根系的累積[19],而對促進土壤碳氮貯量增加的作用則需要較長的時間才能顯現出來。
3 討論
(1)地上生物量的增加是草地恢復演替過程的顯著特征,而根系是植物吸收和貯藏營養物質的主要器官,地下根系的多少直接影響到貯藏營養物質的數量,同時也反映了根系生存環境的優劣狀況[10]。所以生物量是評價生態系統結構與功能的重要參數,對研究系統營養物質分配和營養物質循環具有重要意義[13]。本研究結果表明,與自由放牧地相比,重度退化草地采取生長季圍封措施后,限制了家畜對草地植被的啃食和踐踏,使得處于不同圍封年限的草地地上植物和地下根系生物量明顯增加,并且與圍封時間成正相關。由于碳是組成植物體的結構性物質,氮是生物體的功能性物質,其在植物體的含量直接關系草地生態系統功能的正常發揮[13]。本研究結果表明:地上植物和地下根系中有機碳含量隨圍封年限的增加而增加,而全氮含量隨圍封年限的增加而降低。這是由于家畜采食牧草后,牧草再次生長又促進氮素向地上植物幼嫩器官重新分配,從而導致放牧草地植物氮含量顯著高于圍封樣地[13]。由于植物和根系的碳氮貯量取決于碳氮含量和生物量的大小,使得圍封樣地地上植被和地下根系的碳氮儲量均顯著高于自由放牧地,且與恢復時間成正比。通過地上植物和地下根系生物量與與其碳氮儲量間相關分析得知:地上植物和地下根系生物量與其碳氮貯量呈顯著正相關關系(P<0.01),其相關系數分別為 0.992、0.831和0.967、0.809。說明不同圍封年限草地植物碳氮貯量差異均與其各自生物量差異引起的。
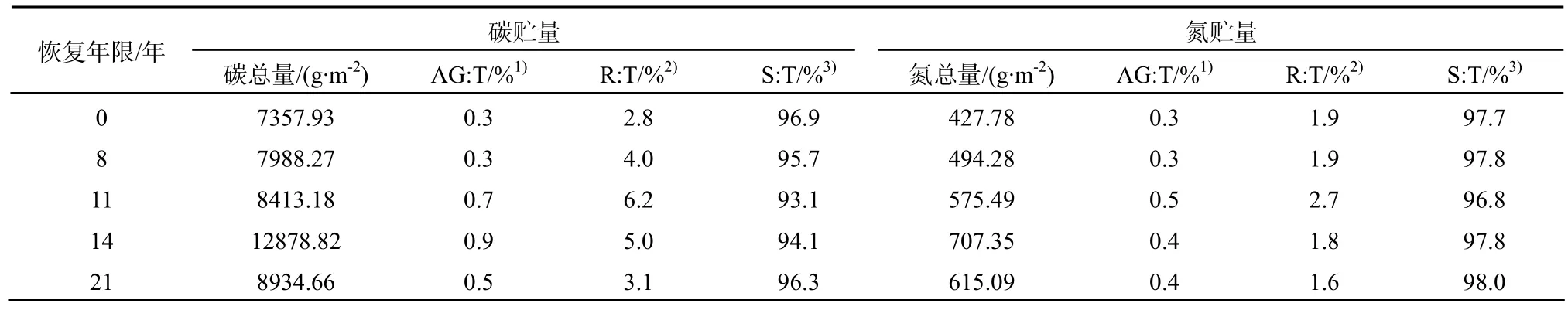
表2 不同圍封年限草地植被-土壤系統碳氮總量及其分配Table 2 The total carbon and nitrogen storages and its distribution of plant-soil system in different years of exclosure
(2)重度退化草地采用生長季圍封恢復措施后,由于限制了放牧家畜踐踏,土壤的結構能力和抗風能力增強,沙化程度的減少,地下根系生物量增加,使得土壤疏松,孔隙增多,容重減小。土壤有機碳、全氮含量是土壤營養狀況的主要標志,也是植物生長所需營養物質的主要來源[20]。本研究表明,隨著圍封時間的增加,土壤有機碳、全氮含量高于自由放牧地,并且與圍封年限增加而增加,而且有機碳和全氮含量呈顯著正相關關系(P<0.01),其相關系數為0.660。說明季節性圍封,其他時間輕度放牧利用的恢復措施能促進家畜糞便的歸還及地表凋落物的分解,有助于有機碳和氮素等積累及土壤肥力的增加。不同圍封年限的草地土壤碳氮儲量的變化和土壤有機質和全氮含量變化一致,均高于自由放牧地,且隨圍封年限的增加而增大。其原因是放牧導致了草地植被的破環,使得草原根系量以及枯落物的積累量都明顯減少,最終導致土壤碳氮儲量的下降。而重度退化草地采取季節性圍封恢復措施后,草地的植被得到了很好的恢復和保護,地上植物和根系生物量與自由放牧地相比明顯提高,從而使草地淺層土壤碳氮貯量有明顯的增加[19]。
(3)處于不同圍封時間的草地,植物-土壤碳氮總量在各部分的分布有著相似的規律,都以土壤碳氮貯量占有最大的比重,一般在93.1%~98%之間;其次為根系,其所占比重在1.6%~6.2%之間;地上植物碳氮貯量所占比重較小,一般低于 1%。許多研究都表明,草原生態系統碳庫中,土壤碳庫占主導地位[21-22]。土壤碳占主導地位對于草地生態系統的碳匯功能來說至關重要。草地生態系統以草本植物為主,不像木本植物那樣會有很高的生物量的積累,并且在生物體中停留的時間較長,所以草原植物本身的貯碳能力較低,作為碳庫的作用較小;而將大部分的碳貯存于地下,由于典型草原獨特的地理位置,地下部分分解較為緩慢,這樣碳在土壤中停留的時間更加長久,其作為碳庫的作用也更為明顯[23]。
(4)通過本研究結果表明,重度退化草地采用生長季圍封恢復措施后,由于控制了家畜的放牧時間與強度,從而保證了退化草地的恢復演替。隨著圍封時間的增加草地植被發育良性化,地上植物和地下根系量明顯增加,土壤容重降低,植物和土壤中碳氮貯量增加,群落結構和生存環境明顯改善,說明生長季(4—9月)圍封收獲干草,其它時間輕度放牧利用的季節性圍封管理措施,一方面可保證退化草地在一定程度上得到恢復,同時也可克服草地圍封恢復過程中不利用或連年放牧等不合理利用問題,季節性圍封禁牧是可行的草地管理方法之一。據王煒等[24]、劉鐘齡等[25]、呂世達等[26]研究報道,重度退化草地采用全年圍封禁牧的管理方式后,8年左右可達到較好的恢復效果,群落植被生產力可基本接近原生群落。但本研究表明恢復年限達到14年左右各項指標達到了最大值,說明草地恢復14年左右可達到較好的恢復效果。說明季節性圍封草地恢復速率比全年圍封草地恢復速率要慢,分析原因為采用季節性圍封措施后,草地在恢復過程中仍存在割草及未圍封季節放牧利用等干擾,導致草地恢復年限比全年圍封禁牧延長。包青海等[27]研究指出,干旱半干旱典型草原合理的輪割制度為割1年休1年或割2年休1年,連年刈割容易造成草地生物量下降,草質變劣,密度下降,草群矮化,引起草地退化。本研究顯示,草地恢復年限達到21到25年時,由于連續多年打草利用草地的各項指標又會出現降低的趨勢,出現了逆行演替,這與劉忠寬等[28-29]研究結果一致,即草地休牧時間過長,植被灌叢化趨勢明顯,可食化程度降低,均勻性降低,草地發生2次逆行演替,所以為保證草地植物可食性及均勻性,退化草地圍封一定時期后提倡進行適度放牧或刈割利用。因此在草地管理中不僅要根據草地退化現狀堅持管理的原則性,同時也要根據草地群落的長勢、年度氣候條件及經濟發展、生態保護及社會需求等靈活掌握[8]。
4 結論
(1)重度退化草地采取生長季圍封恢復措施后,與自由放牧地相比,植被特征比得到了良好的改善,植被生物量及其碳氮貯量均隨圍封年限的增加呈增加趨勢。
(2)與自由放牧地相比,圍封草地土壤容重降低,土壤表層有機碳、全氮含量及其貯量增高,均隨圍封年限的增加呈增加趨勢。
(3)植物-土壤系統碳氮總量在各部分的分布相似的規律,都以土壤碳庫占有最大的比重,其次為根系,植物地上部分所占比重較小,且隨恢復時間時間的增加,地上植物和根系在碳氮總量中所占比重逐漸增大。
植被和土壤碳氮含量在恢復 14年左右時均達到最大值,隨著圍封時間的繼續增長,其植被和土壤特征均有下降趨勢,出現了退化演替,因此恢復一定時期后提倡進行適度放牧或刈割利用,以使草原良性循環。
[1] Batjes N H, Sombroek W G. Possibilities for carbon sequestration in tropical and subtropical soils[J]. Global Change Biology, 1997(3):161-173.
[2] Domaar J F, Sylver Soliak, Walter D Willms. Distribution of nitrogen fractions in grazed and ungrazed fesue grassland Ah horizons [J].Journal of Range Management, 1990, 43 (1):6-9.
[3] Arrouays D, W Deslais, Badeau. The carbon content of topsoil and its geographical distribution in France [J]. Use and Management, 2001,17: 7-11.
[4] Priess J A, G H J de Koning, A Veldkamp. Assessment of interactions between land use change and carbon and nutrient fluxes in Ecuador [J].Agriculture Ecosystem and Environment, 2001, 85:269-279.
[5] Parton W J, M O Scurlock, D S Ojima, et al. Observations and modeling of biomass and soil organic matter dynamics for the grassland biome worldwide [J]. Global Biogeochemical Cycles, 1993,7 (4):785-809.
[6] Houghton R A. Changes in the storage of terrestrial carbon since 1850.In: R Lal, J Kimble, E Levine, and BA Stewart [J]. Soils and global change Inc Boca Raton, Florida: CRC Press, 1995, 45-65.
[7] Nasholm, Torgny, Alf Ekblad, Annika Nordin, Reiner Giesler, Mona Hogberg, Peter Hogberg. Boreal forest plants take up organic nitrogen[J]. Nature, 1998, 392: 914-916.
[8] Conant R T, Paustian K. Potential soil sequest ration in over grazed grassland ecosystems [J]. Global Biogeochemical Cycles, 2002, 16 (4):1143-1151.
[9] 閆玉春,唐海萍. 圍封下內蒙古典型草原區退化草原群落的恢復及其對碳截存的貢獻[J]. 自然科學進展, 2008, 18(5): 546-551.YAN Yuchun, TANG Haiping. Restoration of degraded steppe and its contribution for carbon sequestration with exclosure in the typical steppe of Inner Mongolia in north China[J]. Progress in Natural Science, 2008, 18(5): 546-551.
[10] Reeder J D, Schuman G E. Influence of livestock grazing on C sequest ration in semiarid mixed grass and short-grass rangelands[J].Environmental Pollution, 2002, 116: 457-463.
[11] 李博. 中國北方草地退化及其防治對策[M]. 中國農業科學,1997,30(6): 1-9.LI Bo. The Rangeland Degradation in North China and Its Preventive Strategy [M]. Scientia Agricultura Sinica, 1997, 30(6): 1-9.
[12] 單貴蓮. 內蒙古錫林郭勒典型草原恢復演替研究與健康評價[D].中國農業科學院,2009.SHAN Guilian. Study on Restoration Succession and Health Evaluation of Typical Steppe in Xilinguole, Inner Mongolia[D].Chinese Academy of Agricultural Sciences, 2009.
[13] 董曉玉,傅華,李旭東,等. 放牧與圍封對黃土高原典型草原植物生物量及其碳氮磷貯量的影響[J]. 草業學報,2010,19(2): 175-182.DONG Xiaoyu, FU Hua, LI Xudong, et al. Effects on plant biomass and CNP contents of plants in grazed and fenced steppe grasslands of the Loess Plateaus[J]. Acta Prataculturae Sinica, 2010, 19(2):175-182.
[14] 蘇永中, 趙哈林. 持續放牧和圍封對科爾沁退化沙地草地碳截存的影響[J]. 環境科學, 2003, 24(4): 23-28.SU Yongzhong, ZHAO Halin. The effect s of continuance range and exclosure on Keerqin sandiness grassland carbon capture[J].Environment Science, 2003, 24 (4):232-281.
[15] 何念鵬, 韓興國, 于貴瑞. 長期封育對不同類型草地碳貯量及其固持速率的影響[J]. 生態學報,2011,31(15): 4270-4276.HE nianpeng, HAN xingguo, YU guirui. Carbon and nitrogen sequestration rate in long-term fenced grasslands in Inner Mongolia,China[J]. Acta Ecological Sinica, 2011, 31(15):4270-4276.
[16] Wolde M, Veldkamp E, Mitiku H, et al. Effectiveness of exclosures to restore degraded soils as a result of overgrazing in Tigray, Ethiopia[J].J Arid Environ, 2007, 69: 270-284.
[17] 單貴蓮,徐柱,寧發,等. 圍封年限對典型草原植被與土壤特征的影響[J]. 草業學報,2009, 18(2):3-10.SHAN Guilian, XU Zhu, NING Fa, et al. Influence of seasonal exclosure on plant and soil characteristics in typical steppe [J]. Acta Prataculturae Sinica, 2009, 18(2):3-10.
[18] 寧發,徐柱,單貴蓮. 干擾方式對典型草原土壤理化性質的影響[J].中國草地學報, 2008,30(4):46-50.NING Fa, XU Zhu, SHAN Guilian. Effects of Disturbance Ways on Soil Physical and Chemical Properties[J]. Chinese journal of grassland,2008, 30(4):46-50.
[19] 許中旗,李文華,許晴,等. 人為干擾對典型草原土壤碳密度及生態系統碳貯量的影響[J]. 自然資源學報,2009, 24(4): 621-629.XU Zhongqi, LI Wenhua, XU Qing. The Impacts of Human Disturbances on Soil Carbon Density and Ecosystem Storage of Carbon of Typical Steppes[J]. Journal of Natural Resources, 2009,24(4): 621-629.
[20] 安淵,徐柱,閆志堅. 不同退化梯度草地植物和土壤差異[J]. 中國草地, 1999(4):31-36.AN Yuan, XU Zhu, YAN Zhijian. Diversity of grassland plant and soil along different degradation gradient [J]. Grassland of China, 1999(4):31-36.
[21] 王艷芬,陳佐忠. 人類活動對錫林郭勒地區主要草原土壤有機碳分布的影響[J]. 植物生態學報,1998, 22(6): 545-551.WANG Yanfen, CHEN Zuozhong. Distribution of soil organic carbon in the major grasslands of Xilinguole[J]. Inner Mongolia China Acta Phytoecologica Sinica, 1998, 22(6): 545-551.
[22] 齊玉春. 內蒙古溫帶草地生態系統生物地球化學循環中主要溫室氣體通量與碳平衡[D]. 北京: 中國科學院地理科學與資源研究所,2003.QI Yuchun. The Experiment Study on Main Greenhouse Gases Flux and the Carbon Budget during the Biogeochemical Cycle in the Temperate Grassland Ecosystem in Inner Mongolia[D]. Beijing:Institute of Geographic Sciences and Natural Resources Research,2003.
[23] 李凌浩. 土地利用變化對草原生態系統土壤碳貯量的影響[J].植物生態學報, 1998, 22(4): 300-302.LI Linghao. Effect of land use change on soil carbon storage in grassland ecosystem[J]. Acta Phytoecologica Sinica, 1998, 22(4):300-302.
[24] 王煒,劉鐘齡,郝敦元,等. 內蒙古草原退化群落恢復演替的研究Ⅱ恢復演替時間進程的分析[J]. 植物生態學報, 1996, 20 (5):449-459.WANG Wei, LIU Zhongling, HAO Dunyuan, et al. Research on the restoring Succession of the Degenerated Grassland in Inner MongoliaⅡ. Basic Characteristics and Driving Force for Restoration of the Degenerated Grassland[J]. Acta Phytoecologica Sinica, 1996, 20(5):449-459.
[25] 劉鐘齡,王煒,梁存柱,等. 內蒙古草原退化與恢復演替機理的探討[J]. 干旱區資源與環境, 2002,16(1):84-91.LIU Zhongling, WANG Wei, LIANG Cunzhu, et al. Probes on the Degeneration and Recovery Succession Mechanisms of Inner Mongolia Steppe[J]. Journal of Arid Land Resources and Environment,2002, 16(1): 84-91.
[26] 呂世達, 劉鐘齡, 陳佐忠, 等. 內蒙古半干旱草原土壤-植被-大氣相互作用綜合研究[J]. 氣象學報, 2005,63(5):571-593.Lü Shida, LIU Zhongling, CHEN Zuozhong, et al. Study on soil-vegetation-atmosphere interaction in inner-mongolia semi-arid grassland [J]. Acta Meteorologica Sinica, 2005, 63(5):571-593.
[27] 包青海, 寶音陶格濤, 閻巧玲, 等. 羊草草原割草處理群落特征比較研究[J]. 內蒙古大學學報, 2003,34(1):74-78.BAO Qinghai, BAOYIN Taogetao, YAN Qiaoling, et al. The Comparison Study on Community Characteristics under Mowing Treatments in the Steppe of Leymus chinensis[J]. Acta scientiarum naturalium universitatis neimongol, 2003, 34(1):74-78.
[28] 劉忠寬,汪詩平,陳佐忠,等.不同放牧強度草原休牧后土壤養分和植物群落變化特征[J]. 生態學報, 2006,26(6):2048-2056.LIU Zhongkuan, WANG Shiping, CHEN Zuozhong, et al. Properties of soil nutrients and plant community after rest grazing in Inner Mongolia steppe, China[J]. Acta Ecological Sinica, 2006, 26(6):2048-2056.
[29] 劉忠寬,智建飛,李英杰,等. 休牧后土壤養分空間異質性和植物群落多樣性[J]. 河北農業科學, 2004, 8(4):1-8.LIU Zhongkuan, ZHI Jianfei, LI Yingjie, et al. Spatial Heterogeneity of Soil Nutrients and α Diversity of Plant Community During Rest Grazing[J]. Journal of Hebei agricultural sciences, 2004, 8(4):1-8.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
