不同毒力瘧原蟲感染早期根治性治療對再感染細胞免疫應答的影響
2011-11-13 07:29:18劉英杰潘艷艷曹雅明
中國人獸共患病學報 2011年6期
劉英杰,潘艷艷,李 瑩,馮 輝,劉 軍,曹雅明
2.中國醫科大學免疫學教研室,沈陽 110001
瘧疾流行主要由惡性瘧原蟲和間日瘧原蟲感染所致,但兩個蟲種的毒力存在明顯差異,其中前者毒力較強,是導致重癥瘧疾發生的主要蟲種。瘧疾再感染的發生在流行區普遍存在,但惡性瘧再感染的發生風險顯著高于間日瘧[1],表明原蟲毒力差異與再感染的免疫保護存在密切相關性。細胞免疫應答是抵御胞內寄生病原體再感染的主要因素[2],然而毒力不同的瘧原蟲感染早期根治性治療后,宿主在再感染過程中細胞免疫應答是否存在差異,目前尚未見明確報道。
致死型約氏瘧原蟲(Plasmodium yoelii17XL,Py17XL)和非致死型約氏瘧原蟲(Plasmodium yoelii17XNL,Py17XNL)分別與惡性瘧原蟲和間日瘧原蟲具有相似的生物學特性[3]。為此,我們對Py17XL和Py17XNL感染早期DBA/2小鼠進行根治性治療,然后用同種瘧原蟲進行再感染,通過對再感染前后細胞免疫應答強度的檢測,探討了毒力不同瘧原蟲感染早期的根治性治療對再感染細胞免疫應答的影響。
1 材料與方法
1.1 小鼠、瘧原蟲和實驗感染 雌性,6~8 w齡DBA/2小鼠,分別經腹腔接種1×106Py17XL和Py17XNL寄生的紅細胞。尾靜脈采血,吉姆薩薄血膜染色法計數紅細胞感染率。
1.2 藥物治療和小鼠再感染 初次感染后3 d,經口給藥(氯喹90 mg/kg,青蒿琥脂片60 mg/kg),每天一次,連續3 d。待初次感染后90 d,小鼠用同種瘧原蟲再次攻擊感染。
1.3 脾細胞培養 無菌取出再感染前(0 d)和再感染后第1、3、5 d小鼠脾臟,常規制備脾細胞懸液,調脾細胞終濃度為1×107/mL,于24孔培養板上加入細胞懸液,500 μ L/孔,一式 3 組 ,培養 48 h,收集上清,-80℃保存,待細胞因子檢測。
1.4 脾T細胞中活化性T細胞百分率的檢測 在預先加入FcγⅢ/Ⅱ封閉抗體的染色管中加入脾細胞懸液0.1 mL,再加入抗CD4-FITC和抗CD69-PE mAb進行表面染色,洗滌兩次并懸于500 μ L PBS中,流式細胞儀檢測。
1.5 細胞因子檢測 用ELISA試劑盒按說明書的實驗步驟對小鼠脾細胞培養上清中IFN-γ、TNF-α、IL-4和IL-10的含量進行檢測,酶標儀檢測450 nm處OD值。以試劑盒提供的標準品繪制標準曲線,計算細胞因子含量。
1.6 統計學處理 應用SPSS11.5統計學分析軟件,Student′s-t檢驗比較各組均值的顯著性差異,P<0.05為差異有統計學意義(結果為3次結果的均值)。
2 結 果
2.1 再感染后不同時間的原蟲血癥水平 如圖1所示,兩組感染早期根治性治療小鼠在再感染過程中均出現了短暫的低水平蟲體血癥。在再感染后每一檢測時間點,不同毒力蟲株感染鼠的蟲體血癥水平基本相似,并均于攻擊后第6 d左右自愈;而未治療初次感染小鼠的蟲體血癥水平持續升高,表明毒力不同瘧原蟲初次感染早期根治性治療的小鼠對同種瘧原蟲的再感染具有相似的抵抗力。
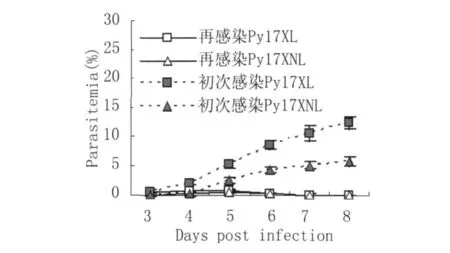
圖1 Py17XL和Py17XNL初次感染和再感染后的蟲體血癥水平Fig.1 The levels of parasitemia after primary infection and reinfection with Py17XL and Py17XNL
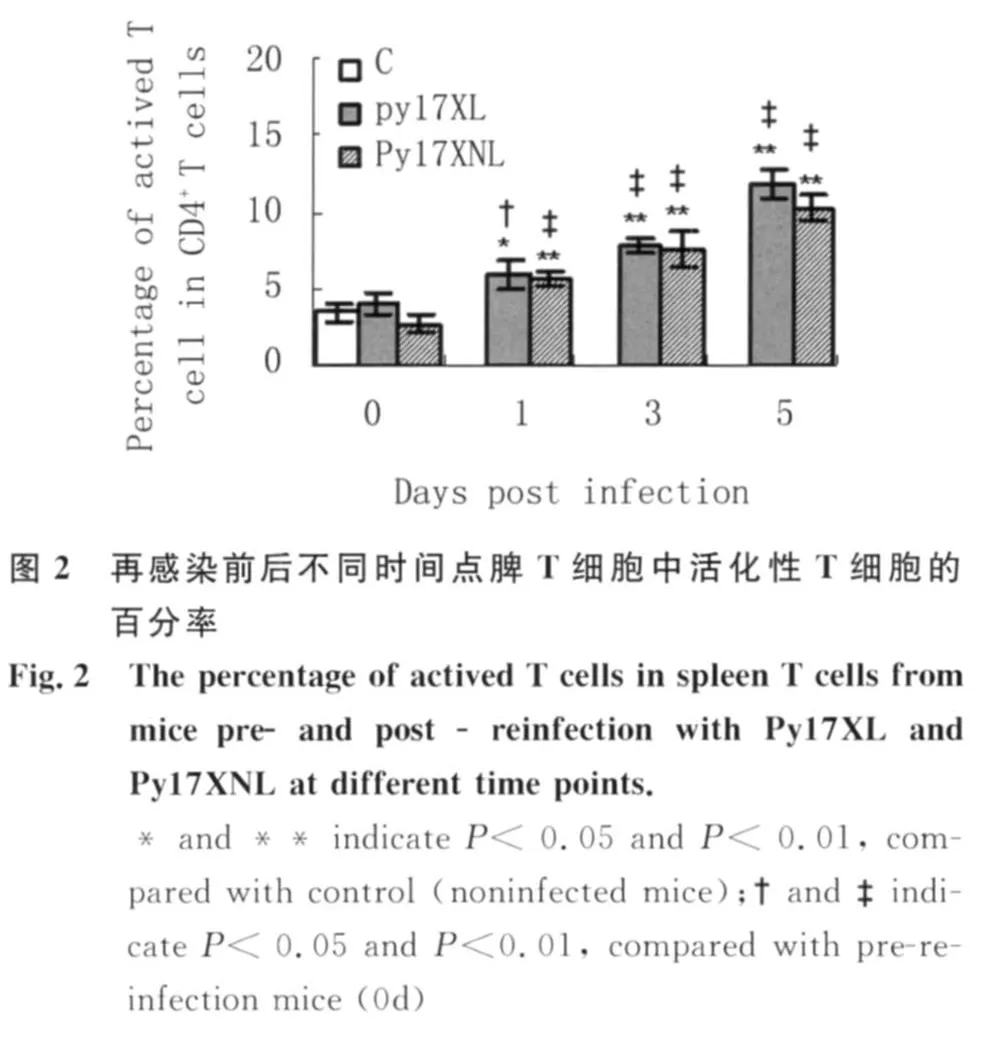
2.2 再感染前后不同時間點脾T細胞中活化性T細胞的百分率 如圖2所示,與正常或再感染前小鼠相比,再感染后第1 d兩組小鼠脾T細胞中活化性T細胞百分率均明顯增加,表明再感染前小鼠體內存在一定數量的記憶性T細胞,當再次感染時這些細胞迅速活化。而此后其進一步升高,則提示除記憶性T細胞活化外,可能又有一些初始T細胞逐漸活化。但在每一檢測時間點,不同毒力蟲株感染鼠活化性T細胞的百分率基本相似。
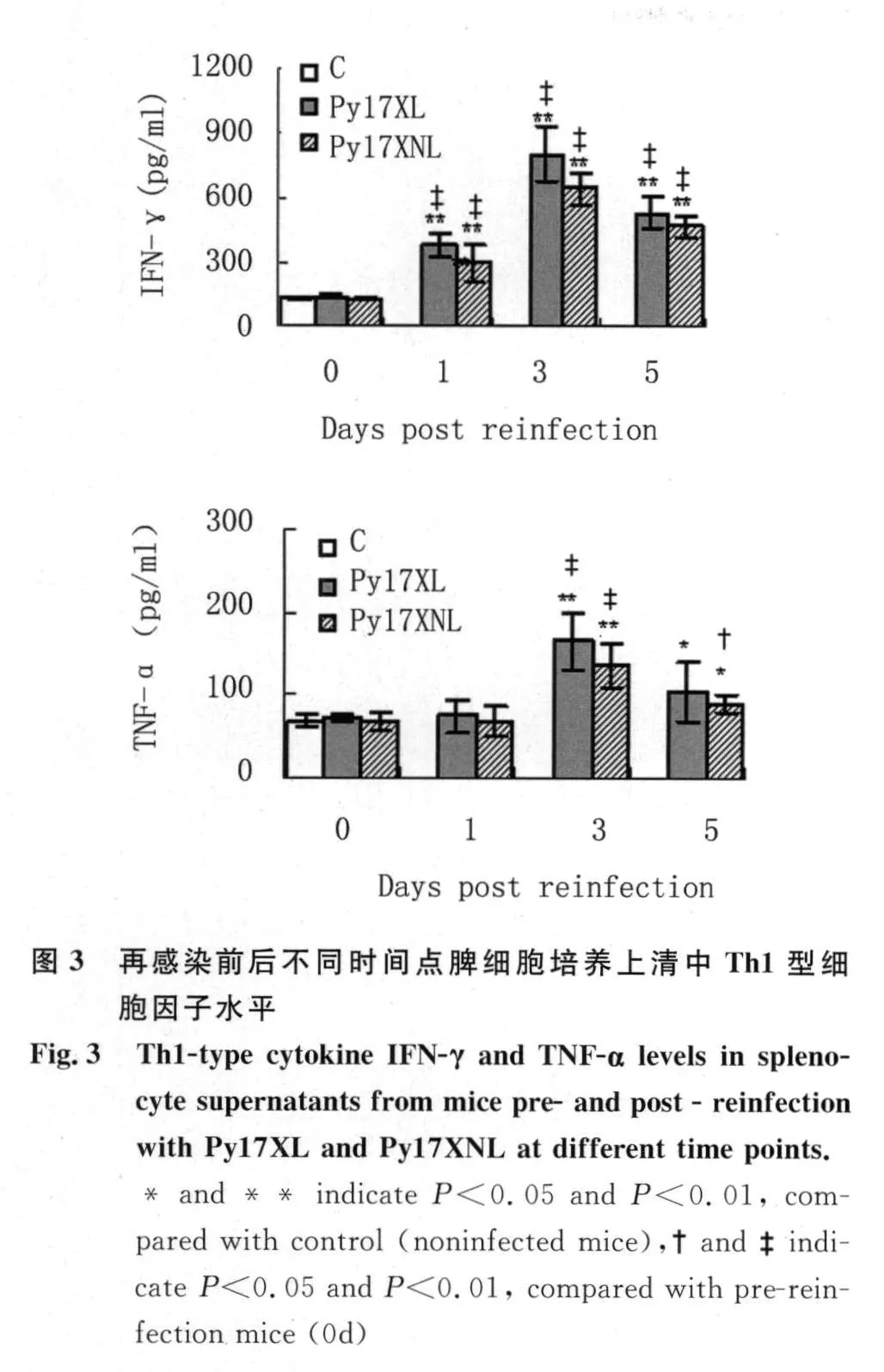
2.3 再感染前后不同時間點脾細胞培養上清中Th1型細胞因子水平 如圖3所示,與正常或再感染前小鼠相比,再感染后第1 d兩組小鼠的IFN-γ水平均出現了有意義的升高,并于第3 d達到峰值水平;與此同時,TNF-α也開始顯著升高。在每一檢測時間點,不同毒力蟲株感染鼠間的IFN-γ和TNF-α水平沒有顯著差異。
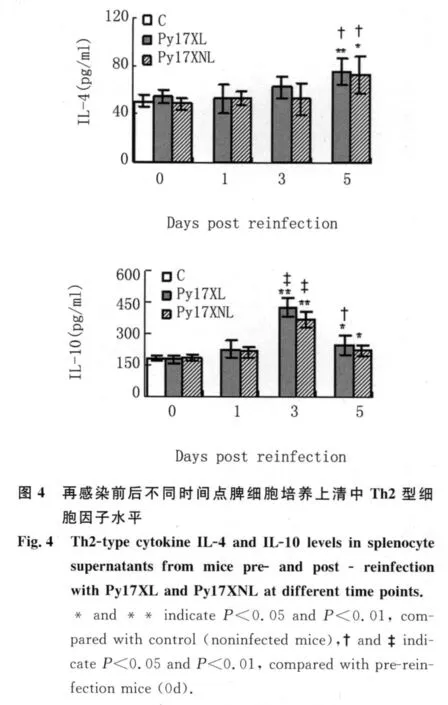
2.4 再感染前后不同時間點脾細胞培養上清中Th2型細胞因子水平 如圖4所示,與正常或再感染前小鼠相比,兩組小鼠的IL-4和IL-10分別在再感染后第5 d和3 d出現了有意義的升高。盡管IL-10在再感染后第5 d出現一定程度的下降,但在每一檢測時間點,不同毒力蟲株感染鼠間的IL-4和IL-10水平并沒有顯著差異。
3 討 論
流行病學調查顯示,惡性瘧再感染的發生風險高于間日瘧[1],盡管惡性瘧再感染后的蟲荷水平有所下降,但多數感染者仍具有較高的蟲荷水平,并有明顯的發熱等臨床表現[4],而間日瘧再感染后的蟲荷水平較低,且沒有明顯的臨床癥狀[5]。有研究證實,CD4+T細胞介導的細胞免疫在抵御瘧疾再感染過程中發揮重要作用[6],間日瘧感染者痊愈后一年左右,其外周血中淋巴細胞可對間日瘧原蟲抗原產生明顯的應答反應(分泌高水平IFN-γ),但惡性瘧感染者卻未能產生此應答反應[7]。這些結果充分提示,再感染的T細胞應答水平與原蟲毒力差異存在密切相關性。
本實驗結果顯示,不同毒力蟲株感染鼠早期根治性治療后對同種瘧原蟲再感染具有相似的抵抗力;其再感染后均可迅速建立以IFN-γ分泌升高為主的高強度Th1應答,而后又產生了適度的以IL-4和IL-10升高為主的Th2應答,并且在每一相同檢測時間點,不同毒力蟲株感染鼠間無論是蟲體血癥水平還是活化性T百分率以及Th1和Th2細胞因子水平均未出現顯著差異。有研究證明,強毒株瘧原蟲感染可損傷或破壞宿主的免疫器官,進而產生免疫抑制效應[3];抗原劑量可改變宿主的免疫應答類型和強度[8-9]。據此我們認為,當 Py17XL和Py17XNL感染的紅細胞在外周血出現時即進行根治性治療可使小鼠受到相同的蟲荷抗原刺激,同時也抑制了強毒株Py17XL對小鼠免疫系統的破壞作用,因此兩組小鼠在再感染時獲得了相似的免疫保護。
綜上所述,不同毒力瘧原蟲感染早期根治性治療后,宿主在同種瘧原蟲再感染時可產生模式和強度相似的細胞免疫應答,強毒株原蟲感染也能獲得與弱毒株相同的可抵御再感染的能力。
[1]Michon P,Cole-Tobian JL,Dabod E,et al.T he risk of malarial infections and disease in Papua New Guinean children[J].Am J Trop Med Hyg,2007,76(6):997-1008.
[2]Grakoui A,Shoukry NH,Woollard DJ,et al.HCV persistence and immune evasion in the absence of memory T cell help[J].Science,2003,302(5645):659-662.
[3]Schaecher K,Kumar S,Yadava A,et al.Genome-wide expression profiling in malaria infection reveals transcriptional changes associated with lethal and nonlethal outcomes[J].Infect Immun,2005,73(9):6091-6100.
[4]Collins WE,Jeffery GM.A retrospective examination of secondary sporozoite-and trophozoite-induced infections withPlasmodium f alciparum:development of parasitologic and clinical immunity following secondary infection[J].Am J Trop Med Hyg,1999,61(1 Suppl):20-35.
[5]Collins WE,Jeffery GM,Roberts JM.A retrospective examination of reinfection of humans withPlasmodium vivax[J].Am J Trop Med Hyg,2004,70(6):642-644.
[6]Praba-Egge AD,Montenegro S,Cogswell FB,et al.Cy tokine responses during acute simianPlasmodium cy nomolgiandPlasmodium knowlesiinfections[J].Am J T rop Med Hyg,2002,67(6):586-596.
[7]Zevering Y,Khamboonruang C,Rung ruengthanakit K,et al.Life-spans of human T-cell responses to determinants from the circumsporozoite proteins ofPlasmodium f alciparumandPlasmodium vivax[J].Proc Natl Acad Sci USA,1994,91(13):6118-6122.
[8]Pombo DJ,Lawrence G,Hirunpetcharat C,et al.Immunity to malaria after administration of ultra-low doses of red cells infected withPlasmodiumf alciparum[J].Lancet,2002,360(9333):610-617.
[9]Freitas do Rosário AP,Muxel SM,Rodr í guez-M á laga SM,et al.Gradual decline in malaria-specific memory T cell responses leads to failure to maintain long-term protective immunity toPlasmodium chabaudi ASdespite persistence of B cell memory and circulating antibody[J].J Immunol,2008,181(12):8344-8355.
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
音樂探索(2022年2期)2022-05-30 21:01:37
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
學苑創造·A版(2020年9期)2020-10-13 09:41:02
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
人大建設(2019年12期)2019-05-21 02:55:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00
