一株嗜熱菌的分離、鑒定及海藻糖合成酶基因的克隆
2011-11-24 07:06:52王慧榮韋宇拓黃日波
湖南師范大學(xué)自然科學(xué)學(xué)報(bào) 2011年6期
關(guān)鍵詞:生長
王慧榮 韋宇拓,黃日波
(1.浙江省環(huán)境保護(hù)科學(xué)設(shè)計(jì)研究院,中國 杭州 310007;2. 廣西大學(xué)生命科學(xué)與技術(shù)學(xué)院,中國 南寧 530005;3.廣西科學(xué)院,中國 南寧 530003)
自從在水生棲熱菌里發(fā)現(xiàn)了DNA聚合酶,促進(jìn)了生物學(xué)發(fā)展和嗜熱酶的商業(yè)開發(fā)和應(yīng)用以后,人們不斷尋找新的嗜熱菌,不僅為了研究嗜熱菌的嗜熱機(jī)理,更多的是尋找有應(yīng)用價(jià)值的嗜熱酶.由于嗜熱酶有熱穩(wěn)定性和在高溫下有最佳活力的特點(diǎn),在造紙工業(yè)、環(huán)護(hù)、能源利用、煙草業(yè)、石油開采、液體燃料的生產(chǎn)、生物轉(zhuǎn)化及抗生素生產(chǎn)等領(lǐng)域都有廣泛的應(yīng)用[1-2].嗜熱酶的廣泛應(yīng)用促使人們不斷尋找新的嗜熱菌.海藻糖(trehalose)是細(xì)胞在不良環(huán)境中所產(chǎn)生的一種重要的抗逆應(yīng)激物,在環(huán)境脅迫條件下對(duì)生命體和生物活性物質(zhì)具有保護(hù)功能,在食品工業(yè)、農(nóng)業(yè)、醫(yī)藥工業(yè)、分子生物學(xué)等領(lǐng)域有廣泛的應(yīng)用前景[3].雖然目前,已從ThermusaquaticusATCC33923、ThermusthermophilusHB8等多種嗜熱菌中克隆了海藻糖合成酶編碼基因(treS),并且進(jìn)行了異源表達(dá)[4-5].但是嗜熱菌來源的海藻糖合成酶分子量一般較大,表達(dá)量不高,需要解決密碼子偏愛性等問題以提高在大腸桿菌中的表達(dá)量.由于海藻糖合成酶三維晶體結(jié)構(gòu)的研究還未見報(bào)道,缺乏足夠的數(shù)據(jù)詳細(xì)闡述它的催化機(jī)理,通過空間分子結(jié)構(gòu)的改造來改善酶學(xué)性質(zhì)的研究有一定的困難,目前亟需大量的關(guān)于海藻糖合成酶的序列、結(jié)構(gòu)和生物學(xué)特性等相關(guān)數(shù)據(jù).
本文從廣西熱泉中分離到一株嗜熱菌,初步鑒定它為Thermusthermophilus,對(duì)該菌的海藻糖合成酶基因進(jìn)行了初步探索,為構(gòu)建高產(chǎn)海藻糖合成酶的工程菌提供理論基礎(chǔ)及重要的實(shí)驗(yàn)依據(jù);另一方面,也進(jìn)一步豐富不同微生物中該酶基因序列和結(jié)構(gòu)方面的信息.
1 材料和方法
1.1 樣品采集與處理
從廣西賀州市西雞鎮(zhèn)80 ℃,pH為6.7,含鹽量2 g/L的熱泉中采集水樣和泥樣.水樣以12 000 r/min離心10 min,收集菌體;泥樣用泉水徹底懸浮后,以500 r/min離心2 min,去除大顆粒的雜質(zhì),然后再用處理水樣的方式處理上清液富集菌體.
1.2 菌株、載體及試劑
大腸桿菌JM109由本實(shí)驗(yàn)室保存,PCR產(chǎn)物純化試劑盒和膠回收試劑盒購自上海華舜生物工程有限公司,限制性內(nèi)切酶,T4DNA連接酶,pMD 18-T vector購自寶生物工程(大連)有限公司,其他試劑均為國產(chǎn)分析純.
1.3 培養(yǎng)基
分離培養(yǎng)基[6]:NaCl 1 g,MgCl2·6H2O 1 g,MgSO4·7H2O 0.15 g,CaCl20.23 g,KCl 0.5 g,KH2PO40.42 g,(NH4)2SO40.1 g,NaBr 0.05 g,SrCl2·6H2O 0.02 g,NaHCO31 g,Yeast extract 1 g,Tryptone 1 g,肉汁胨0.5 g,F(xiàn)eCl3(100 g/L) 10 mL,1 000×Vitamin Solution 10 mL,Trace element solution 1 mL,NaOH調(diào)pH值至7.0,1×105Pa 滅菌30 min.Vitamin Solution用0.45 μm濾膜過濾,固體培養(yǎng)基加20 g瓊脂.
1.4 嗜熱菌的分離與培養(yǎng)
取處理好的樣品直接涂布于固體培養(yǎng)基上,分別置于75 ℃和80 ℃培養(yǎng)箱中培養(yǎng),待菌落長出后,挑取單個(gè)菌落進(jìn)行劃線培養(yǎng),分別置于85 ℃,86 ℃,87 ℃下培養(yǎng)24~48 h.篩選能在最高溫度下生長的菌,從而實(shí)現(xiàn)菌株的分離與純化.為防止溫度過高在培養(yǎng)過程中平板脫水干裂,在培養(yǎng)時(shí)將平板倒置放在一個(gè)10 L的玻璃器皿中,里面放置1杯水,每天定期打開玻璃器皿通氧.
1.5 形態(tài)觀察和培養(yǎng)條件
1.5.1 形態(tài)觀察 固體培養(yǎng)基上培養(yǎng)了24~48 h的單菌落,用電子顯微鏡觀察細(xì)胞形態(tài),革蘭氏染色光學(xué)顯微鏡觀察.
1.5.2 培養(yǎng)溫度 接種一個(gè)單菌落到10 mL液體培養(yǎng)基中,培養(yǎng)使其OD600達(dá)到0.6~0.8時(shí),以2%的接種量接種到100 mL,pH為7.0的液體培養(yǎng)基中,在220 r/min條件下,分別置于55、60、65、70、75、80、85 ℃下,每隔2 h取1.5 mL菌液測定OD600.
1.5.3 培養(yǎng)pH值 同樣以2%的接種量接種到100 mL, pH值分別為5.5,6.0,6.5,7.0,7.5,8.0,8.5,9.0的液體培養(yǎng)基中,70 ℃,220 r/min條件下,每隔2 h取1.5 mL菌液測定OD600.
1.5.4 NaCl濃度的測定 以2%的接種量接種到100 mL NaCl質(zhì)量濃度分別為0,4,8,10,12 g/L,pH值為7.0的液體培養(yǎng)基中,在75 ℃,220 r/min條件下培養(yǎng).每隔2 h取1.5 mL菌液測定OD600.
1.6 G+C 物質(zhì)的量濃度和抗生素敏感性的測定
用溶解溫度(Tm)法[7],參照文獻(xiàn)[8] 的濃度進(jìn)行測定.
1.7 16S rRNA基因的PCR擴(kuò)增、序列測定和系統(tǒng)發(fā)育分析
取對(duì)數(shù)期的菌液,離心后收集菌體,CTAB法提取總DNA.依照細(xì)菌16S rRNA基因擴(kuò)增的通用引物:F27(5-AGAGTTTGATCCTGGCTCAG-3),R1522(5-AAGGAGGTGATCCAGCCGCA-3),以TTH總DNA為模板,PCR反應(yīng)條件:94 ℃變性4 min,94 ℃ 1 min,60 ℃ 1 min,72 ℃ 1.5 min,共25個(gè)循環(huán),72 ℃延伸6 min.PCR產(chǎn)物純化后連接到 pMD 18-T vector,經(jīng)過抽提質(zhì)粒和酶切驗(yàn)證酶連接產(chǎn)物正確,委托上海基康生物技術(shù)有限公司進(jìn)行測序.將得到的16S rRNA序列與GenBank中的核酸數(shù)據(jù)進(jìn)行BLAST比對(duì),選取與其同源性達(dá)到98%的相關(guān)序列.采用軟件ClustalX對(duì)所獲得的核苷酸序列進(jìn)行序列比對(duì),用軟件MEGA4.1計(jì)算出序列的系統(tǒng)進(jìn)化距離,采用鄰位相連法構(gòu)建系統(tǒng)進(jìn)化樹.
1.8 treS基因的擴(kuò)增
根據(jù)GenBank中已公布的模式菌株ThermusthermophilusHB8的treS基因序列設(shè)計(jì)引物,引物1:5′- TGGTAAACCCGCTCCCACAG -3′,引物2:5′- TGCCTGGGCACGGAAAAGAA-3′.以TTH總DNA為模板,PCR條件為94 ℃ 5 min; 94 ℃ 30 s, 58 ℃ 30 s, 72 ℃ 2 min, 30個(gè)循環(huán)后72 ℃延伸10 min.PCR產(chǎn)物經(jīng)瓊脂糖凝膠電泳驗(yàn)證后進(jìn)行膠回收,與pMD 18-T vector 4℃過夜連接,將連接產(chǎn)物轉(zhuǎn)化E.coliJM109,藍(lán)白斑篩選,篩選陽性克隆,菌落PCR檢測目的條帶,提取陽性菌的質(zhì)粒.酶切鑒定,測序由上海基康生物技術(shù)有限公司完成.

圖1 菌株TTH的電鏡照片(×10 000)
2 結(jié)果
2.1 形態(tài)和培養(yǎng)特征
菌株TTH為革蘭氏陰性的桿狀細(xì)胞,大小為(0.4~0.6)μm×(3.0~4.5)μm,無鞭毛、芽孢.培養(yǎng)48 h的菌落呈圓形,中間稍有凸起,邊緣整齊,表面光滑,有光澤,淺黃色,不透明.電鏡照片見圖1.
2.2 溫度對(duì)TTH生長的影響
菌株TTH的生長溫度范圍是55 ℃~85 ℃,低于55 ℃,高于85 ℃都不生長,在55 ℃,85 ℃下生長極度貧乏,最適生長溫度是70 ℃左右,培養(yǎng)約12 h進(jìn)入穩(wěn)定期.結(jié)果見圖2.
2.3 pH對(duì)TTH生長的影響
菌株TTH生長pH值的范圍是5.0~9.0,在5.0,5.5和9.0下生長十分緩慢,低于5.0,高于9.0條件下不生長.最適的生長pH值為6.5~7.5,結(jié)果見圖3.

圖2 溫度對(duì)菌株TTH生長的影響
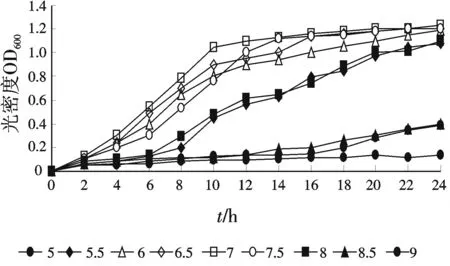
圖3 pH對(duì)菌株TTH生長的影響

圖4 NaCl質(zhì)量濃度對(duì)菌株TTH生長的影響
2.4 NaCl對(duì)TTH生長的影響
TTH菌株生長的NaCl質(zhì)量濃度范圍是0~12 g/L,在12 g/L NaCl濃度下生長貧乏,大于12 g/L NaCl不生長,最適的NaCl濃度是0~4 g/L.結(jié)果見圖4.
2.5 G+C物質(zhì)的量分?jǐn)?shù)和抗生素敏感性
菌株TTH的G+C物質(zhì)的量分?jǐn)?shù)為67.6%.參照文獻(xiàn)[9],加氨芐青霉素、氯霉素到液體培養(yǎng)基中,使其終濃度為10 mg/L,70 ℃,24 h未見生長.在加了鏈霉素、卡那霉素的培養(yǎng)基中,鏈霉素、卡那霉素的終濃度在50 mg/L時(shí),菌株都可以生長.四環(huán)素10 mg/L時(shí)不抑制生長,100 mg/L時(shí)可完全抑制生長.
2.6 以16S rRNA序列為基礎(chǔ)的系統(tǒng)發(fā)育分析
菌株TTH 16S rRNA全序列總共1 478個(gè)堿基,在GenBank登錄號(hào)為EU616794.1.將其序列在GenBank中進(jìn)行BLAST比較,發(fā)現(xiàn)菌株TTH與多株棲熱菌屬(Thermus)的16S rRNA序列同源性高達(dá)98%,依據(jù)此結(jié)果,菌株可歸屬于棲熱菌屬(Thermus).選擇BLAST比對(duì)結(jié)果中相似度最高的5條16S rRNA,利用ClustalX 1.8和 MEGA4.1軟件,采用鄰位相連法構(gòu)建系統(tǒng)發(fā)育樹.圖5表明菌株TTH與Thermusthermophilus的3株菌在同一族上,其中與ThermusthermophilusXM、Thermusthermophilusit-1、Thermusthermophilus這3株菌的同源性高達(dá)98%,結(jié)合形態(tài)學(xué)觀察、生理生化特征指標(biāo)測定及16S rRNA序列分析結(jié)果,確定菌株TTH是棲熱菌屬中的Thermusthemophilus.

圖5 菌株TTH的系統(tǒng)發(fā)育進(jìn)化樹

1: treS gene; M: DL2000 Marker圖6 PCR產(chǎn)物瓊脂糖凝膠電泳
2.7 treS基因的擴(kuò)增
以TTH的總DNA為模板,采用引物1和引物2進(jìn)行PCR擴(kuò)增treS基因,對(duì)PCR產(chǎn)物進(jìn)行瓊脂糖凝膠電泳分析,結(jié)果如圖6所示.
從圖6中可以看出PCR擴(kuò)增出了約1 200 bp的片段,將PCR產(chǎn)物與pMD 18-T vector連接后,轉(zhuǎn)化E.coliJM109,藍(lán)白斑篩選,菌落PCR和酶切驗(yàn)證正確后,對(duì)treS基因進(jìn)行測序,其結(jié)構(gòu)基因全長為1 250 bp,GenBank accession:GQ175175,測序結(jié)果在GenBank中Blast進(jìn)行比對(duì),發(fā)現(xiàn)它與菌株ThermusthermophilusRQ-1,Thermuscaldophilus,ThermusthermophilusHB8,Thermusthermophilus的treS基因的同源性最高可達(dá)87%,作者克隆的該菌的treS基因有可能是海藻糖合成酶的同源基因,treS基因的獲得為TTH菌株完整的treS基因的克隆奠定了基礎(chǔ).
3 討論
早在1971年日本人就從當(dāng)?shù)?5 ℃~85 ℃的熱泉中分離到了Thermusthemophilus,從此對(duì)該菌進(jìn)行了大量的研究,但是已知的Thermusthemophilus大多從海洋熱泉中分離得到,能夠耐鹽[10].本文從陸地?zé)崛蟹蛛x的TTH菌株生長的NaCl質(zhì)量濃度范圍是0~12 g/L,在12 g/L NaCl濃度下生長貧乏,NaCl濃度大于12 g/L時(shí)不生長,最適的NaCl濃度是0~4 g/L.在Thermusthemophilus里有好氧菌和厭氧菌,菌株TTH是好氧菌,文獻(xiàn)中HB27只能在好氧條件下生長,HB8不僅可在好氧條件下生長,還能在硝酸鹽存在的條件下厭氧生長.與TTH硝酸鹽還原反應(yīng)陰性相一致,可見在自然界中存在多種不同的Thermusthemophilus菌株[11].由此可以看出嗜熱微生物資源相當(dāng)豐富,還有很大的探索空間,篩選出更多的嗜熱菌不僅可以豐富嗜熱微生物資源,還為研究嗜熱菌機(jī)理提供了實(shí)驗(yàn)素材.
本實(shí)驗(yàn)室通過DNA Shuffling技術(shù)尋求新的耐高溫海藻糖合成酶基因的研究已經(jīng)多年,從已報(bào)道的Thermusthermophilus不同菌株中克隆了海藻糖合成酶基因[12],但是從嗜熱菌TTH中克隆的treS基因與這些菌中存在的海藻糖合成酶基因還是有差異的,且測定的序列與已報(bào)道的ThermusthermophilusRQ-1,Thermuscaldophilus,ThermusthermophilusHB8,Thermusthermophilus的treS基因的同源性最高達(dá)87%,由此作者認(rèn)為Thermusthermophilus的treS基因序列既有保守性,又有在不同環(huán)境條件脅迫下產(chǎn)生的進(jìn)化差異性.對(duì)菌株TTH的treS基因序列進(jìn)行進(jìn)一步的分析,將會(huì)為理解該酶的作用機(jī)制,以及該酶在不同細(xì)菌中的進(jìn)化演變情況提供有價(jià)值的參考.
參考文獻(xiàn):
[1] LI J B, XU L M, YANG F. Expression and characterization of recombinant thermostable alkaline phosphatase from a novel thermophilic bacteriumThermusthermophilusXM[J].Acta Bioch Bioph Sin, 2007,39(11):844-850.
[2] MITSUNORI I, SATOSHI K, ATSUSHI K,etal. Purification and characterization of the recombinantthermussp. strain T2α-galactosidase expressed inEscherichiacoli[J]. Appl Environ Microb, 2001,67(4):1601-1606.
[3] EIBEIN A D, PAN Y T, PASTUSZAK I,etal. New insights on trehalose: a multifuntional molecule[J].Glycobiol,2003, 60(13):17-27.
[4] TSUSAKI K, NISHIMOTO T, NAKADA T,etal. Cloning and sequencing of trehalose synthase gene fromthermusaquaticusATCC33923[J].Acta Bioch Bioph Sin, 1997,1334(45):28-32.
[5] ZDZIEBLO A, SYNOWIECKI J. Production of trehalose by intramolecular transglucosylation of maltose catalysed by a new enzyme fromThermusthermophilusHB-8[J]. Food Chem, 2006,96(6):8-13.
[6] WOESE C R, KANDLER O, WHEELIS M L. Towards a natural system of organisms: Proposal for the domains Archaea, Bacteria, and Eucarya[J]. Proc Natl Acad Sci USA,1990, 87:4576-4579.
[7] 阮繼生,劉志恒,梁麗糯,等. 放線菌研究及應(yīng)用[M]. 北京: 科學(xué)出版社, 1990.
[8] J.薩姆布魯克,E.F.弗里奇.分子克隆實(shí)驗(yàn)指南[M].金冬雁,黎孟楓,譯.2版.北京:科學(xué)出版社,2001:548.
[9] 王慧榮,韋宇拓,黃日波.一個(gè)新型嗜熱菌質(zhì)粒的測序鑒定及抗生素抗性研究[J].湖南師范大學(xué)自然科學(xué)學(xué)報(bào),2010,33(2):102-106.
[10] SANDRA T M, FRENANDA N, FRED A R,etal.Thermonemarossianumsp. Nov., a new thermophilic and slightly halophilic species from saline hot springs in Naples, Italy[J]. Int J Syst Bacterilol, 1997,47(1):122-126.
[11] FELIPE C, OLGA Z, AXEL M,etal. A new type of NADH dehydrogenase specific for nitrate respiration in the extreme thermophilethermusthermophilus[J]. J Biol Chem, 2004, 29(24):45369-45378.
[12] 韋傳東.海藻糖合成酶基因的融合表達(dá)、純化以及酶學(xué)性質(zhì)研究[D]. 南寧:廣西大學(xué),2006.
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(shù)(2020年3期)2020-12-06 07:32:54
現(xiàn)代裝飾(2020年11期)2020-11-27 01:47:48
中學(xué)生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(jí)(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時(shí)刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
