體外培養獼猴穆勒細胞經誘導可表現出視網膜前體細胞特征
2011-12-25 06:39:54郭祥玉金立方紀少琿魏強牛昱宇季維智
Zoological Research 2011年6期
郭祥玉, 金立方, 紀少琿, 魏強, 牛昱宇, 季維智
(中國科學院昆明動物研究所 靈長類研究中心, 云南省動物生殖生物學重點實驗室, 云南 昆明 650223)
體外培養獼猴穆勒細胞經誘導可表現出視網膜前體細胞特征
郭祥玉, 金立方, 紀少琿, 魏強, 牛昱宇, 季維智*
(中國科學院昆明動物研究所 靈長類研究中心,云南省動物生殖生物學重點實驗室,云南 昆明650223)
近年來的研究發現, 雞、大鼠和人視網膜中的穆勒膠質細胞(muller cell)在一定條件下,可顯示出視網膜干/前體細胞的功能。但是, 有關人類近親獼猴穆勒細胞的研究尚未見報道。該研究首先使用基礎培養基(DMEM+10%FBS)建立獼猴的穆勒細胞系, 其表達GS、vimentin、CRALBP和EGFR等穆勒細胞標記, 不表達GFAP,與人的穆勒細胞基因表達相似而與嚙齒類差異較大。穆勒細胞經神經干細胞培養基培養一周后可表達pax6、nestin、sox2、otx2、six6和six3等視網膜干細胞標記, 繼續使用視黃酸誘導, 部分細胞可分化為神經元細胞,這說明獼猴的穆勒細胞在體外誘導條件下,可去分化為視網膜前體細胞, 有望成為視網膜疾病細胞替代療法的一種細胞來源。
獼猴; 視網膜干細胞; 穆勒細胞; 視網膜疾病
神經退行性視網膜疾病,如青光眼、視網膜色素變性、老年性黃斑變性等是目前的高致盲性疾病(Wallace, 2007), 其共同病理特征是部分視網膜神經細胞的病變死亡, 臨床上尚無有效的治療手段。干細胞具有分化的多能性, 被認為可分化為與病變細胞相應的細胞類型并取而代之, 從而治愈疾病。因此, 使用視網膜干細胞治療上述眼疾是一種具有良好發展前景的治療策略。
穆勒細胞是視網膜中廣泛存在的一類放射狀膠質細胞, 主要起支撐視網膜結構, 為視網膜神經元和血管提供代謝支持的作用, 同時還具有防止感光細胞過分向視網膜下生長的作用(Limb et al, 2002)。在發育上, 穆勒細胞和視網膜其他神經細胞具有共同的祖細胞。近年來研究發現, 在成年的雞(Fischer & Reh, 2001)和大鼠(Ooto et al, 2004)中, 視網膜被NMDA藥物損傷后,穆勒細胞會顯著增殖,部分表達視網膜干細胞抗原, 并可遷移至損傷部位分化成相應的細胞。嚙齒類動物(Das et al, 2006; Wan et al, 2007)穆勒細胞已被成功分離在體外培養,并被證實其確實具有視網膜前體細胞的屬性, 具有一定自我更新能力且可分化成視網膜特異的神經元。人的穆勒細胞雖然在體外條件下也可被誘導去分化, 但是其去分化后更與神經干細胞相似(Lawrence et al, 2007)。
嚙齒類動物的視網膜的生理結構、功能與人類差異較大, 而非人靈長類動物的視網膜與人類極為相似, 是研究視網膜疾病的理想模型(West et al, 2009)。目前, 關于非人靈長類穆勒細胞的研究還未見報道。本研究首次建立了獼猴穆勒細胞系, 分析了獼猴穆勒細胞體外培養條件下的基因表達情況,使用神經干細胞培養體系誘導穆勒細胞去分化為視網膜前體細胞, 并進一步誘導分化出神經元細胞。
1 材料和方法
1.1 試驗動物
獼猴眼球來自于中國醫學科學院昆明生物醫學研究所用于生產疫苗的不同年齡段的成年健康獼猴(n=30), 這些獼猴從未用于其他的實驗。
1.2 穆勒細胞的分離培養
眼球先用含有青霉素和鏈霉素的PBS清洗并剪除周邊組織, 然后在角膜緣后1 cm左右處進行環切, 將眼球后緣部分浸入含有0.25%胰蛋白酶(sigma)+1%膠原酶的DMEM/F12溶液中, 放置在37 ℃ 1 h。最后用眼科鑷小心地將神經視網膜剝離出來(圖1A), 注意避免色素上皮的污染。剝離出來的視網膜先用剪刀剪碎, 然后用含有0.25%胰蛋白酶+1%dispase的DMEM/F12在37 ℃消化0.5 h。消化好的細胞用含10%胎牛血清+EGF(10 ng/mL)的DMEM培養基以105/mL濃度懸浮, 最后接種至明膠處理好的培養皿中。
1.3 誘導穆勒細胞去分化及再分化
將傳代培養貼壁后的穆勒細胞培養基更換為無血清培養基:DMEM/F12+N2 supplement+FGF2 (20 ng/mL)+EGF(20 ng/mL), 此后隔天更換半量培養基。細胞傳代使用胰蛋白酶消化, 按1∶3的比例傳至上代細胞的條件培養基中。在無血清培養條件下培養一周后進行視網膜干細胞標記檢測。使用無血清培養基培養一周后改換為神經分化培養基(DMEM/F12+1%FBS+50 mmol/L RA)進行誘導分化,培養4周后進行分化檢測。
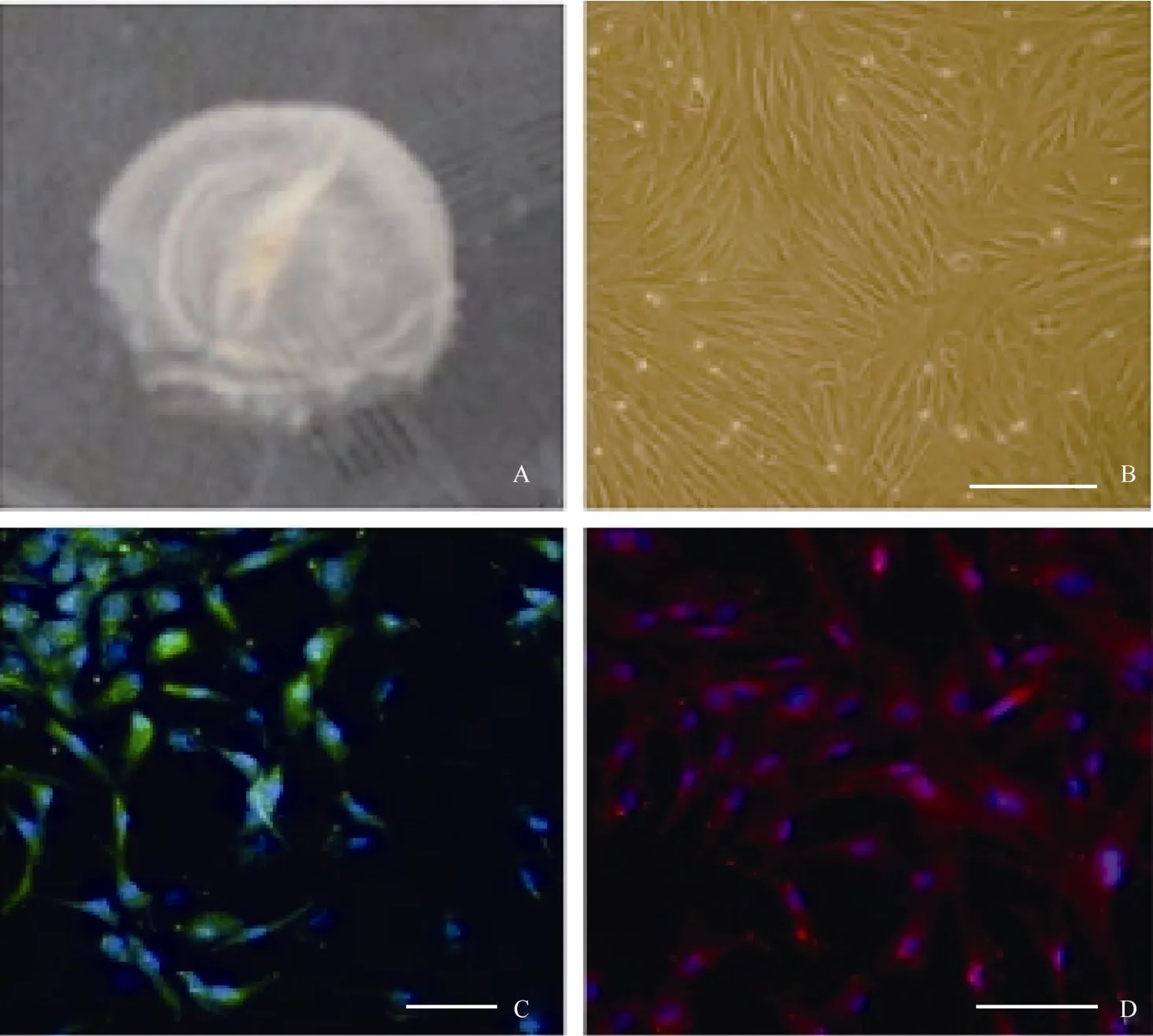
圖1 穆勒細胞體外培養及免疫熒光檢測Fig.1 Immunofluorescence test for in vitro cultured muller cell
1.4 細胞免疫熒光染色
PBS沖洗細胞3次,4%多聚甲醛室溫下固定15~20 min;PBS沖洗細胞3次, 0.4%Triton X-100透膜10 min(僅對細胞內蛋白);隨后再次用PBS 沖洗細胞3次,5%~10%羊血清室溫封閉30 min。細胞在一抗中室溫孵育2 h或4 ℃過夜。PBS洗3次后加入FITC-或PE/Texas Red(Santa Cruz)連接的二抗,室溫孵育1 h。用Hochest 33258進行細胞核染色。以添加二抗, 而不添加一抗的細胞作為陰性對照組,檢測二抗的假陽性情況。使用的一抗包括:Glutamate synthetase(1∶600, sigma)、vimentin(1∶400, Dako)、Pax6(1∶600, Abcam)、nestin(1∶600, chemicon)、otx2(1∶600, chemicon)免疫標記的細胞在激光共聚焦掃描顯微鏡(Zeiss, LSM 510 META)下進行檢測。
1.5 逆轉錄-多聚酶鏈式反應(reverse transcriptionpolymerase chain reaction, RT-PCR)
根據廠家提供的實驗步驟, 用Trizol試劑(Invitrogen, Calsbad, CA)從培養的細胞中直接提取總RNA。用DNaseI消化總RNA中可能含有的基因組DNA, 去除DNA的污染。
逆轉錄反應:在20 μL體系(1× reverse transcription buffer, 0.5 mol/L dNTPs, 50 pmol oligo dT primer, 20 U RNase inhibitor, 5 U reverse transcriptase)中加入1 μg總RNA, 在PCR儀(MJ Research, Watertown, MA)上42 ℃保溫1.5 h后65 ℃繼續保溫 10 min。PCR反應:在25 μL的PCR體系(10× PCR reaction buffer, 1.5 mmol/L MgCl2, 0.5 mmol/L dNTPs, 0.4 μmol/L 正反向引物, 1.25 U rTaq DNA聚合酶)中加入1 μL RT產物(表1)擴增25~35個PCR循環(95 ℃,30 s, 52~60 ℃,30 s, 72℃,30 s)以保證PCR反應仍然處于線形擴增階段,隨后在72 ℃延伸5 min。PCR反應所用引物見表2, 所有的RT-PCR試劑均購于Takara (Takara, Dalian, China)。PCR產物在2% 瓊脂糖膠上電泳后,用溴化乙錠(ethidium bromide)染色, 以管家基因GAPDH為標準, 用Quantity-One軟件(Bio-Rad)通過電泳條帶的強弱程度, 評價基因表達量的高低。

表1 PCR反應體系Tab. 1 PCR reaction system
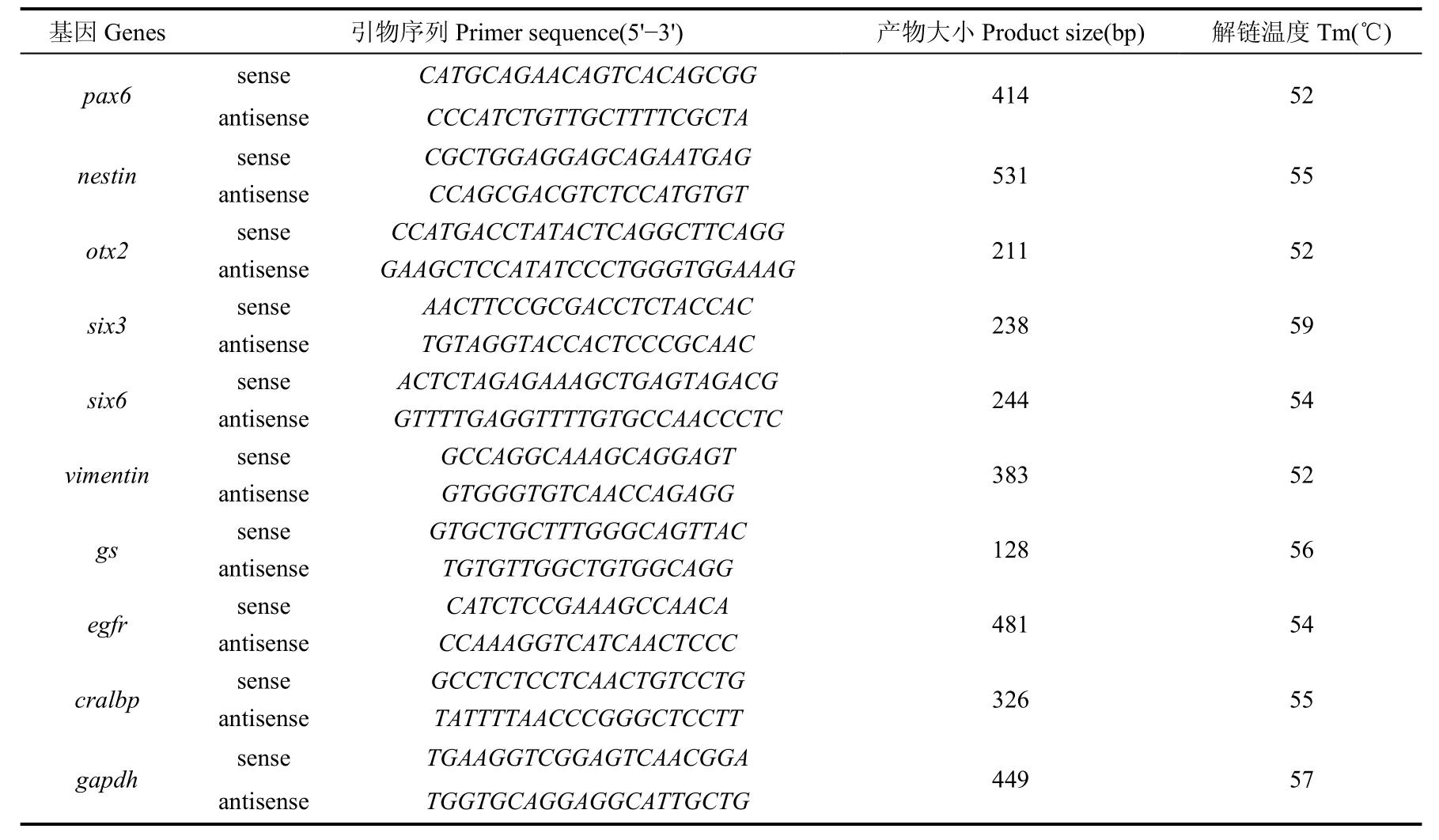
表2 PCR引物信息Tab. 2 Primer information for PCR
1.6 統計分析
統計數據以平均值±標準誤 (Mean±SD) 進行表示。統計分析采用SPSS 10.0軟件進行One-Way ANOVA分析。P<0.05限定為差異顯著。
2 結 果
2.1 獼猴穆勒細胞系的建立及基因表達特征
在含有10%FBS的培養條件下, 接種48 h細胞開始貼壁生長, 穆勒細胞單層貼壁生長, 呈成纖維細胞樣, 分裂周期約為24 h。體外傳代可達30次以上。我們從13個猴子中建立12株細胞系(92.3%)。使用免疫熒光和RT-PCR的方法分析第3、15和30代細胞, 發現穆勒細胞穩定表達GS[(99.3±0.5) %], vimentin[(97.7±2.1)%), EGFR和CRALBP等穆勒細胞特征基因(圖1C, D;圖2)而少有GFAP的表達。同時, 檢測視網膜干細胞標記基因發現, 在含10%FBS的培養條件下, 穆勒細胞不表達pax6、nestin,、sox2、otx2、six6、six3等視網膜干細胞標記(圖3; 圖4A, B, C)。我們的結果說明,獼猴穆勒細胞可在體外大量擴增且屬性較為穩定。
2.2 穆勒細胞去分化為視網膜前體細胞
據研究報道, 成年哺乳動物, 包括人的穆勒細胞一定條件下在體內外可去分化視網膜干細胞。第3、15、30代培養的獼猴穆勒細胞貼壁后,我們將培養基更換為含有FGF2、EGF、N2 supplemnt的無血清培養基。培養一周后, 免疫熒光染色檢測發現穆勒細胞開始表達pax6、nestin、otx2等視網膜干細胞標記基因(圖4 A, B, C), RT-PCR的檢測同時也驗證了上述視網膜干細胞基因的表達, 而且six3和six6兩個視網膜發育相關基因也有表達, 而穆勒細胞相關基因GS、vimentin、EGFR和CRALBP的表達出現了顯著的下降(圖3)。但是, 隨著細胞的傳代,視網膜干細胞標記基因的表達會出現明顯下降(數據未顯示)。該結果證明,體外培養的獼猴穆勒細胞可誘導去分化為視網膜干細胞, 但較難長時間維持在視網膜干細胞的狀態。
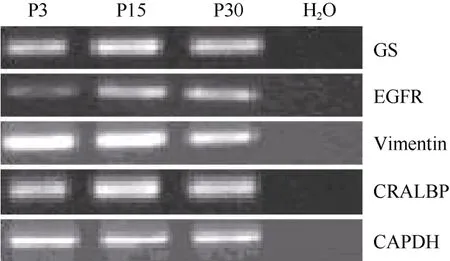
圖2 不同代數穆勒細胞基因表達的RT-PCR分析Fig.2 RT-PCR analysis of gene expression of muller cells at different passage
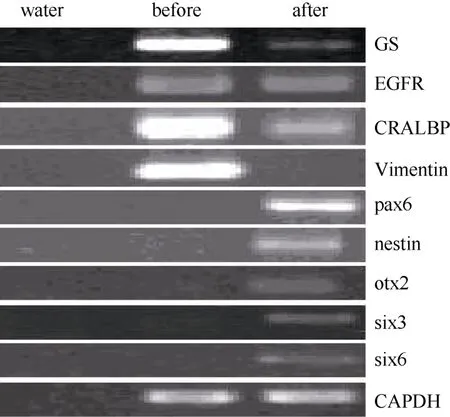
圖3 穆勒細胞特征基因及視網膜干細胞基因在神經干細胞培養基誘導前后表達差異Fig.3 Expression switch of muller cell specific genes and retinal stem cell genes after induction
2.3 去分化后的穆勒細胞向神經細胞分化
干細胞的一個重要標準是具有分化的多能性(West et al, 2009)。獼猴穆勒細胞經神經干細胞培養基培養一周后, 使用含有1%FBS和30 mmol/L視黃酸的DMEM/F12培養基誘導培養3周后發現, 有部分細胞分化成神經元 (圖4D), 但是使用免疫熒光染色法未檢測到有視網膜特異類型的神經細胞形成, 該結果說明獼猴的穆勒細胞可以跨譜系分化為神經元細胞, 但是能否分化成特異的視網膜神經元需要進一步研究。
3 討 論
本研究成功建立了獼猴穆勒細胞系, 該系可在體外長期傳代并保持屬性的穩定。表達GS、vimintin、EGFR、CRALBP; 而不表達GFAP, 這與人的穆勒細胞基因表達特征相似(Limb et al, 2002),而與大鼠(Birnbach et al, 1994)和貓(Lewis et al, 1988)的有所不同。另外, 穆勒細胞與視網膜中的星形膠質細胞在外形和基因表達方面都非常相似, 而且我們的細胞分離方法使得培養的細胞有可能混雜有星形膠質細胞。星形膠質細胞與穆勒細胞的主要區別是星形膠質細胞表達GFAP且不表達CRALBP, 我們建立的細胞系表達CRALB;而不表達GFAP。 因此, 我們建立的細胞系是穆勒細胞而不是星形膠質細胞。
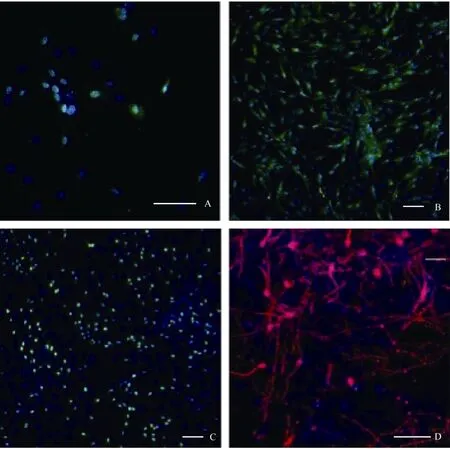
圖4 誘導去分化及再分化后相關基因的免疫熒光檢測Fig.4 Immunofluorescence test for gene expression after induction and redifferentiation
穆勒細胞是視網膜中最后分化出的細胞類型,有學者認為穆勒膠質細胞是視網膜干細胞進入靜息狀態的一種存在形式, 當有生理需要的時候穆勒細胞就會又返回到視網膜干細胞的狀態(Agathocleous & Harris, 2009)。在雞(Fischer & Reh, 2001)和大鼠的試驗中視網膜在受損傷的情況下, 穆勒細胞可起到視網膜干細胞的作用。體外分離培養的嚙齒動物(Das et al, 2006; Wan et al, 2007)和人(Lawrence et al, 2007)的穆勒細胞也具有一定視網膜干細胞特征。我們分離培養的獼猴穆勒細胞同樣在誘導條件下可去分化, 與人的穆勒不同的是獼猴的更多的表達視網膜干細胞的基因, 而人的則更傾向于去分化為神經干細胞。
雖然獼猴的穆勒細胞在體外去分化的過程中可表達多種視網膜干細胞基因, 但是神經干細胞培養基并不能長期維持穆勒細胞去分化為視網膜干細胞的狀態。這可能是由于培養條件不夠理想, 僅能誘導其去分化而不能抑制其再分化造成的, 同時說明獼猴的穆勒膠質細胞與其他物種的穆勒細胞一樣具有可塑性, 研究優化誘導和培養條件可為發育生物學研究及臨床應用提供良好的平臺。
Agathocleous M, Harris WA. 2009. From progenitors to differentiated cells in the vertebrate retina[J]. Ann Rev Cell Dev Biol,25: 45-69.
Birnbach CD, Jarvelainen M, Possin DE, Milam AH. 1994. Histopathology and immunocytochemistry of the neurosensory retina in fundus flavimaculatus[J]. Ophthalmology,101: 1211-1219.
Das AV, Mallya KB, Zhao X, Ahmad F, Bhattacharya S, Thoreson WB, Hegde GV, Ahmad I. 2006. Neural stem cell properties of Muller glia in the mammalian retina: regulation by Notch and Wnt signaling[J]. Dev Biol,299: 283-302.
Fischer AJ, Reh TA. 2001. Muller glia are a potential source of neural regeneration in the postnatal chicken retina[J]. Nat Neurosci,4: 247-252.
Lawrence JM, Singhal S, Bhatia B, Keegan DJ, Reh TA, Luthert PJ, Khaw PT, Limb GA. 2007. MIO-M1 cells and similar muller glial cell lines derived from adult human retina exhibit neural stem cell characteristics[J]. Stem Cells,25: 2033-2043.
Lewis GP, Kaska DD, Vaughan DK, Fisher SK. 1988. An immunocytochemical study of cat retinal Muller cells in culture[J]. Exp Eye Res,47:855-868.
Limb GA, Salt TE, Munro PM, Moss SE, Khaw PT. 2002. In vitro characterization of a spontaneously immortalized human Muller cell line (MIO-M1) [J]. Invest Ophthalmol Vis Sci,43: 864-869.
Ooto S, Akagi T, Kageyama R, Akita J, Mandai M, Honda Y, Takahashi M. 2004. Potential for neural regeneration after neurotoxic injury in the adult mammalian retina[J]. Proc Natl Acad Sci USA,101: 13654-13659.
Wallace VA. 2007. Stem cells: a source for neuron repair in retinal disease[J]. Can J Ophthalmol,42: 442-446.
Wan J, Zheng H, Xiao HL, She ZJ, Zhou GM. 2007. Sonic hedgehog promotes stem-cell potential of Muller glia in the mammalian retina[J]. Biochem Biophys Res Commun,363: 347-54.
West EL, Pearson RA, MacLaren RE, Sowden JC, Ali RR. 2009. Cell transplantation strategies for retinal repair[J]. Prog Brain Res,175: 3-21.
Rhesus monkey (Macaca mulatta) muller cells exhibit retinal stem/progenitor cell featuresin vitro
GUO Xiang-Yu, JIN Li-Fang, JI Shao-Hui, JI Wei-Zhi*
(Yunnan Key Laboratory for Animal Reproductive Biology, Kunming Institute of Zoology, the Chinese Academy of Sciences, Kunming 650223, China)
Recent evidences indicate that retinal muller cells exhibit retinal progenitor characteristics under certain condition in chick, rat and human. However, there is no report on nonhuman primate, a close relative to human. In this study, we first established a muller cell line of rhesus monkey expressing GS, vimentin, CRALBP, EGFR, barely GFAP, which resemble the expression profile of human muller cells, differ from that of rodent. Expression of pax6, nestin, sox2, otx2, six6, and six3 was detected after one week culture in neural stem cell medium. Further culture with retinoic acid induced some cells differentiate toward neuron. These results suggest that primate muller cell is capable of dedifferentiating to retinal progenitors, which may serve as a potential cell source for cell therapy to treat retinal degenerative diseases.
Rhesus monkey; Retinal stem cells; Muller cell; Retinal degenerative diseases
Q813.11; Q959.848; R774.1
A
0254-5853-(2011)06-0611-06
10.3724/SP.J.1141.2011.06611
2011-05-10;接受日期:2011-10-20
科技部重大專項(2009ZX09501-028);云南社會科技發展計劃(2007GH);中科院知識創新工程重大項目(KSCX1-YW-22)
?通訊作者(Corresponding author),Tel/fax: 86-0871-5139413, E-mail: wji@mail.kiz.ac.cn

- Zoological Research的其它文章
- New details indicated by different stainings during conjugation of ciliated protozoa Paramecium
- Histological and immunocytochemical study of deferens ducts in the Chinese rat snake (Zaocys dhumnades)
- PTEN在小鼠卵細胞和早期胚胎發育中的功能研究
- A phylogeny of the Tylototriton asperrimus group (Caudata: Salamandridae) based on a mitochondrial study: suggestions for a taxonomic revision
- 云南新平哀牢山西黑冠長臂猿分布與群體數量
- 杭州灣及錢塘江河口水鳥群落組成、季節動態及種間相關性分析