華山新麥草葉片的光合生理特性
2011-12-31 13:50:20柴永福張曉飛
植物資源與環境學報 2011年3期
柴永福,李 倩,張曉飛,岳 明
(西北大學西部資源生物與現代生物技術教育部重點實驗室,陜西 西安 710069)
華山新麥草(Psathyrostachys huashanica Keng ex P.C.Kuo)為多年生草本植物,隸屬于禾本科(Poaceae)新麥草屬(Psathyrostachys Nevski),是中國特有種,主要分布在陜西華山的華山峪、黃浦峪和仙峪[1]。華山新麥草是小麥屬(Triticum L.)植物的野生近緣種,具有抗病、抗旱和早熟等特點,在探索小麥屬植物的起源、進化及遺傳育種等方面具有重要價值[2],已被列為國家一級珍稀瀕危植物和急需保護的農作物野生親緣種[3]。
近年來,對華山新麥草的研究主要集中在系統進化、雜交育種和生殖生物學等方面[4],而關于華山新麥草生理生態學方面的研究尚未見報道。為了有效保護華山新麥草這一野生種質資源、探討其致瀕機制,作者對華山新麥草的光合生理特性進行了分析。
1 自然分布現狀和研究方法
1.1 自然分布現狀
目前,華山新麥草僅在陜西省華陰市境內的華山有分布。華山是秦嶺支脈華山山脈的一部分,地理坐標為北緯34°25′~34°30′、東經109°57′~110°05′;該區域屬大陸性季風氣候,四季分明,季節氣溫差別較大,冬季寒冷干燥、夏季炎熱多雨;年均降雨量902.7 mm,年均氣溫5.9℃,最高溫度27.7℃,最低溫度-24.1℃。華山新麥草分布的海拔范圍較廣,從山麓平原(海拔351 m)到山頂(海拔2 153 m)均有分布,海拔高差1 800 m[5];該種在向陽地段分布較密集,在背光處分布較稀疏,在林下或陰濕處未見分布;喜干旱較溫暖氣候,在降雨較少的中低山地區及山麓地帶分布數量相對較多,而在高山地帶分布數量相對較少[6];多生長在巖石凹陷或空隙處;生長的土壤主要是微生物分解土層表面腐敗的植物脫落或死亡器官形成的腐殖質,土壤中的水分主要依靠降水,因此,降水的不確定性導致了華山新麥草分布區域土壤含水量的不規律性[5]。
1.2 材料和方法
供試華山新麥草采自華山黃浦峪并引種栽培于西安市西北大學生物系果園內,株高約40~50 cm。栽培所用土壤均來源于華山新麥草原生長地,栽培期間每隔3 d澆水1次。
于2010年4月上旬在晴朗無云的天氣條件下隨機選擇健康、正常的華山新麥草營養葉5片,用美國CID公司生產的CI-301PS便攜式光合測定儀測定凈光合速率、蒸騰速率、葉溫、氣孔阻力、光合有效輻射強度和氣溫等相關指標。測定時系統處于開放狀態,測定時間為每天7:30至19:30,1 h測量1次,重復測量5 d,結果取平均值。
1.3 數據處理與分析
運用Statistic 6.0軟件對實驗數據進行統計分析,采用最小二乘法對光響應方程進行擬合,日變化曲線和光響應曲線均使用Origin 7.5軟件完成。
2 結果和分析
2.1 凈光合速率的日變化
華山新麥草凈光合速率日變化曲線見圖1。與大多數植物凈光合速率的日變化曲線呈“雙峰”型的趨勢有明顯不同[7-11],華山新麥草凈光合速率的日變化曲線呈“三峰”型,共包含有3個峰值,3個峰值出現的時間分別為9:30、11:30和16:30,對應的凈光合速率分別為6.5、6.2和9.0μmol·m-2·s-1,第2個峰和第3個峰之間出現“午降”現象,“午降”時(中午12:30左右)華山新麥草葉片的凈光合速率較低,僅為2.5μmol·m-2·s-1。
2.2 蒸騰速率的日變化
蒸騰作用的強弱是衡量植物水分代謝能力的重要生理指標[12]。華山新麥草蒸騰速率的日變化曲線見圖2。結果表明:華山新麥草蒸騰速率的日變化曲線呈“單峰”型,峰值為1.7 mmol·m-2·s-1,出現在13:30;在11:30至13:30,葉片蒸騰速率的上升速率最快;13:30后葉片的蒸騰速率逐漸下降,在19:30下降至0.3 mmol·m-2·s-1。
2.3 葉溫日變化及其對凈光合速率的影響
華山新麥草葉溫的日變化曲線以及葉溫與其凈光合速率的關系圖分別見圖3和圖4。華山新麥草葉溫的日變化曲線呈“單峰”型(圖3),在12:30達到最大值(39.8℃)。根據葉溫與凈光合速率的變化擬合得到華山新麥草凈光合速率對葉溫的響應方程y= -0.023 78x2+1.371 18x-13.123 50(r2=0.353 9)。由圖4可見:在一定葉溫范圍內,凈光合速率與葉溫呈正相關,隨著葉溫的升高凈光合速率逐漸增大,當葉溫達到28℃時凈光合速率最大,但葉溫超過28℃時凈光合速率開始逐漸下降。
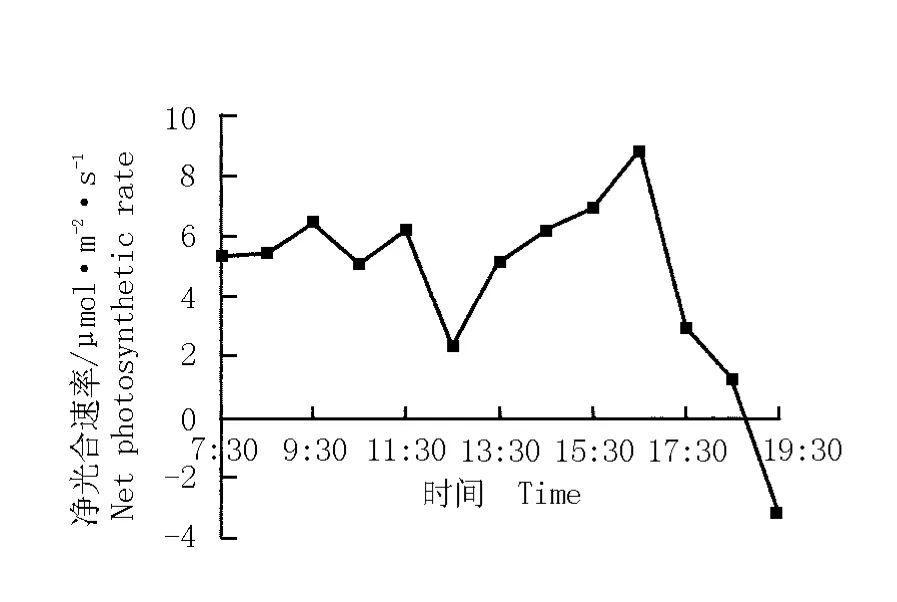
圖1 華山新麥草凈光合速率的日變化曲線Fig.1 Daily change curve of net photosynthetic rate of Psathyrostachys huashanica Keng ex P.C.Kuo
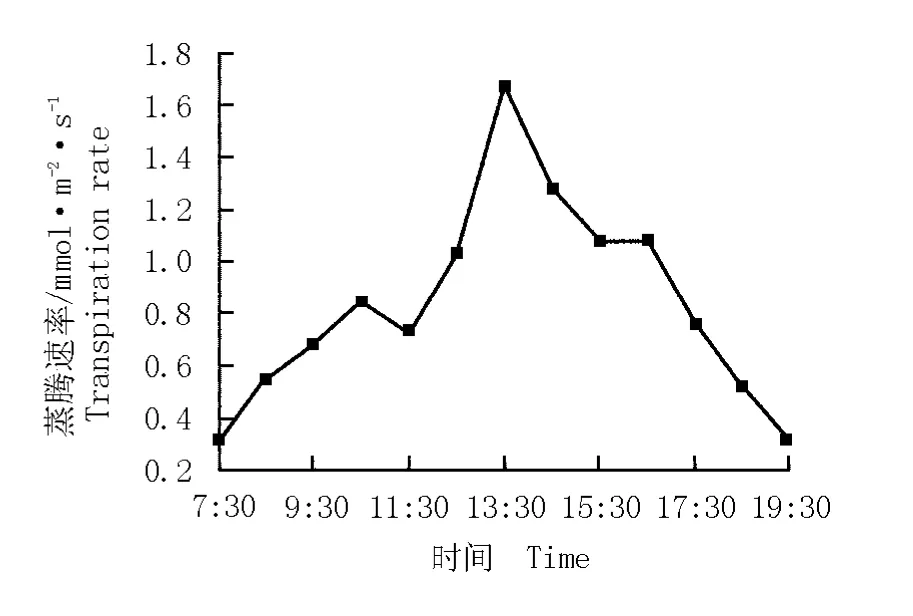
圖2 華山新麥草蒸騰速率的日變化曲線Fig.2 Daily change curve of transpiration rate of Psathyrostachys huashanica Keng ex P.C.Kuo
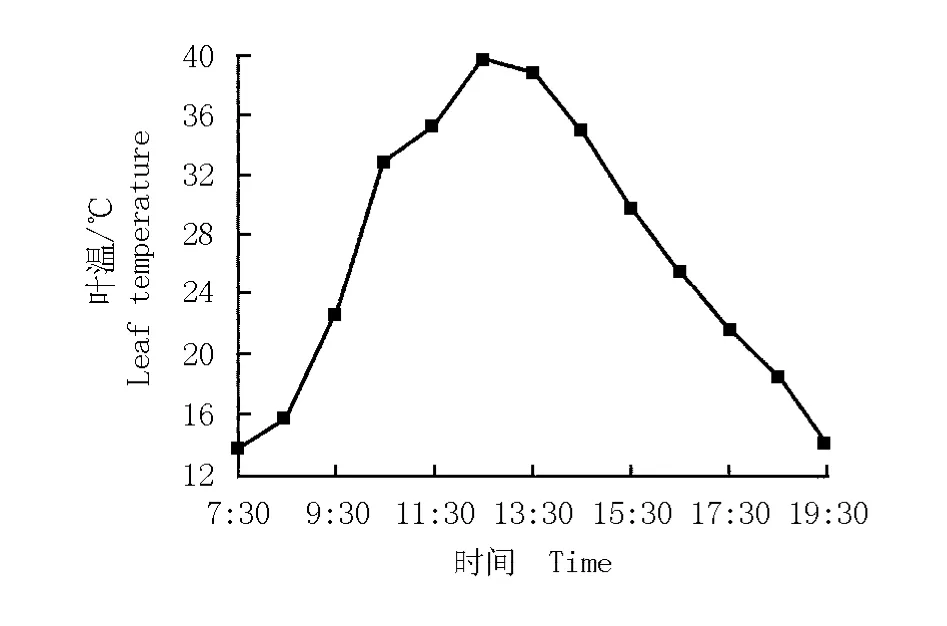
圖3 華山新麥草葉溫的日變化曲線Fig.3 Daily change curve of leaf tem perature of Psathyrostachys huashanica Keng ex P.C.Kuo
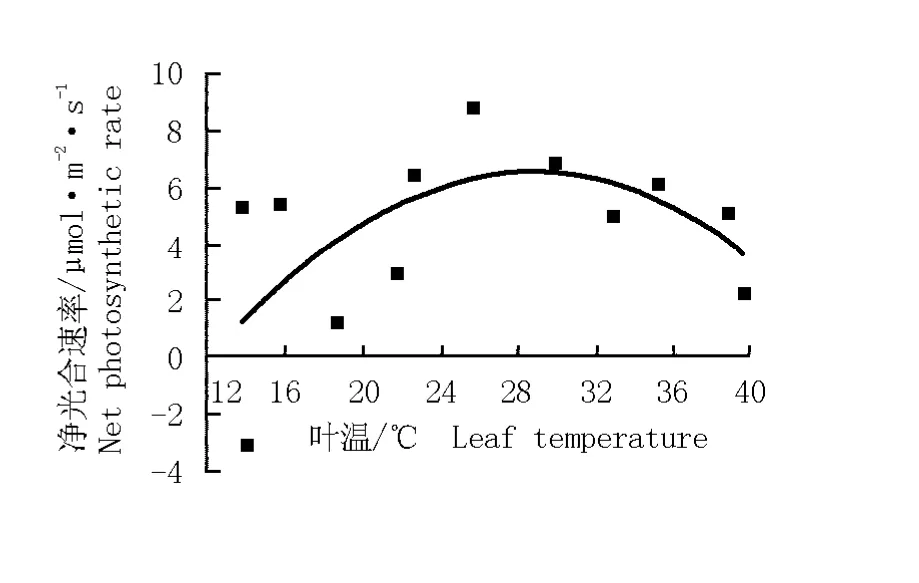
圖4 華山新麥草凈光合速率對葉溫的響應曲線Fig.4 Response curve of net photosynthetic rate of Psathyrostachys huashanica Keng ex P.C.Kuo to leaf tem perature
2.4 氣孔阻力日變化及其對凈光合速率的影響
華山新麥草氣孔阻力的日變化曲線以及氣孔阻力與其凈光合速率的關系圖分別見圖5和圖6。結果表明:華山新麥草氣孔阻力的日變化曲線呈“單峰”型,在11:30達到最大值,為74 m2·s·mol-1。根據氣孔阻力與凈光合速率的變化擬合得到華山新麥草凈光合速率對氣孔阻力的響應方程y=0.000 37x2-0.104 19x+7.895 66(r2=0.510 6)。由圖6可見:華山新麥草的凈光合速率與氣孔阻力呈負相關關系,隨著氣孔阻力的增大,凈光合速率急速下降。
2.5 光合有效輻射強度日變化及其對凈光合速率的影響
華山新麥草生境中光合有效輻射強度的日變化曲線及其與凈光合速率的關系圖分別見圖7和圖8。由圖7可看出:華山新麥草生境中光合有效輻射強度的日變化曲線呈“單峰”型。從7:30開始,光合有效輻射強度迅速增大,10:30達到600μmol·m-2·s-1,12:30達到最大值(900μmol·m-2·s-1);隨后,光合有效輻射強度逐漸降低,至19:30幾乎接近于0μmol·m-2·s-1。
根據光合有效輻射強度與凈光合速率的變化擬合得到華山新麥草凈光合速率對光合有效輻射強度的響應方程y=-0.000 03x2+0.031 89x-0.035 49 (r2=0.669 0)。由圖8可見:華山新麥草的凈光合速率隨光合有效輻射強度增大而呈“拋物線”型變化,光合有效輻射強度在300~700μmol·m-2·s-1范圍內凈光合速率較高;凈光合速率達到最高時的光合有效輻射強度為500μmol·m-2·s-1。將葉片凈光合速率為0μmol·m-2·s-1時的光照強度作為光補償點(light compensation point,LCP),將凈光合速率最高時的拐點作為光飽和點(light saturation point,LSP),根據擬合方程計算得出華山新麥草的LCP和LSP分別為1.1和531.5μmol·m-2·s-1。
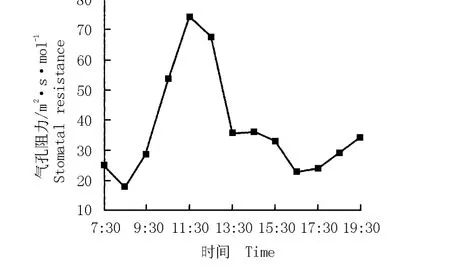
圖5 華山新麥草氣孔阻力的日變化曲線Fig.5 Daily change curve of stomatal resistance of Psathyrostachys huashanica Keng ex P.C.Kuo
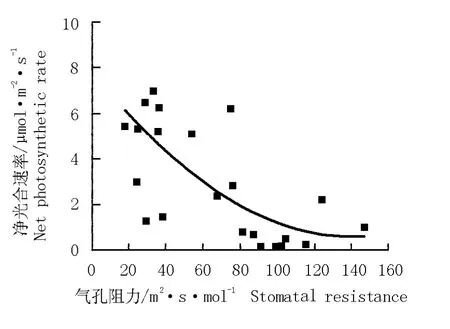
圖6 華山新麥草凈光合速率對氣孔阻力的響應曲線Fig.6 Response curve of net photosynthetic rate of Psathyrostachys huashanica Keng ex P.C.Kuo to stomatal resistance
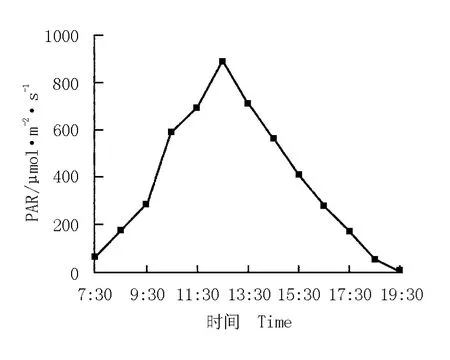
圖7 華山新麥草生境中光合有效輻射強度(PAR)的日變化曲線Fig.7 Daily change curve of photosynthetically active radiation intensity(PAR)in habitat of Psathyrostachys huashanica Keng ex P.C.Kuo
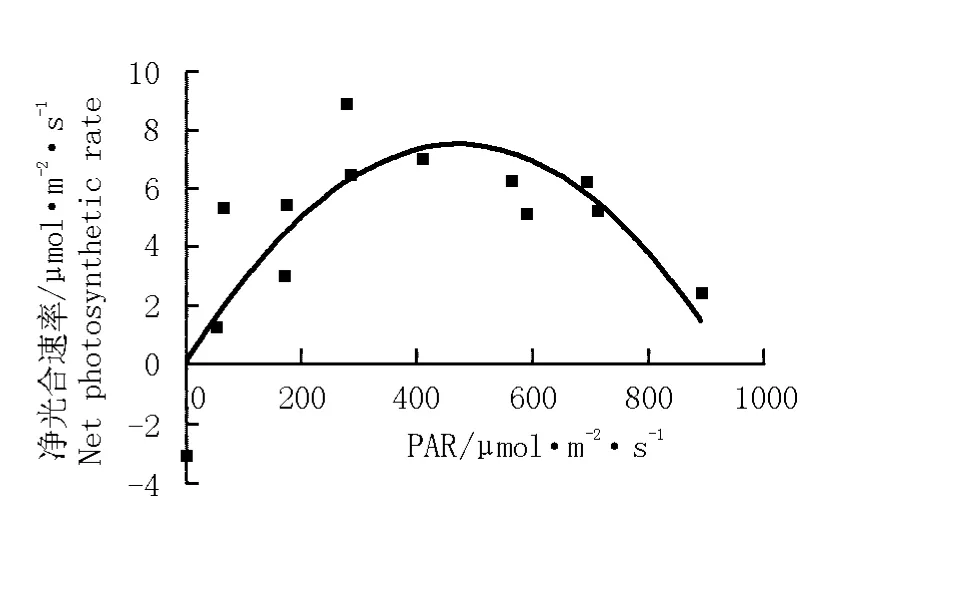
圖8 華山新麥草凈光合速率對光合有效輻射強度(PAR)的響應曲線Fig.8 Response curve of net photosynthetic rate of Psathyrostachys huashanica Keng ex P.C.Kuo to photosynthetically active radiation intensity(PAR)
2.6 氣溫日變化及其對凈光合速率的影響
氣溫與光合有效輻射強度常常是密切相關的,光合有效輻射強度大,氣溫也相應較高,因而,氣溫與植物的凈光合速率也具有相關性。華山新麥草生境中氣溫的日變化曲線及其與凈光合速率的關系圖分別見圖9和圖10。圖9顯示:華山新麥草生長環境的氣溫日變化曲線也呈“單峰”型,從7:30開始氣溫持續升高;10:30氣溫已達到26.8℃;12:30至14:30間的氣溫較高,保持在32℃以上;13:30氣溫達到峰值,隨后逐漸下降。
根據華山新麥草凈光合速率與氣溫的變化擬合得到華山新麥草凈光合速率對氣溫的響應方程y= -0.138 89x2+7.162 14x-85.225 39(r2=0.394 0)。由圖10可見:華山新麥草凈光合速率對氣溫的響應曲線也呈“拋物線”型,與其對光合有效輻射強度的響應曲線相似。在一定范圍內(低于26℃),隨著氣溫的升高,凈光合速率增大;氣溫超過26℃,凈光合速率逐漸下降。對于華山新麥草而言,適宜的氣溫為20℃~30℃,最適氣溫為26℃,在此氣溫條件下華山新麥草的凈光合速率可達到最高。
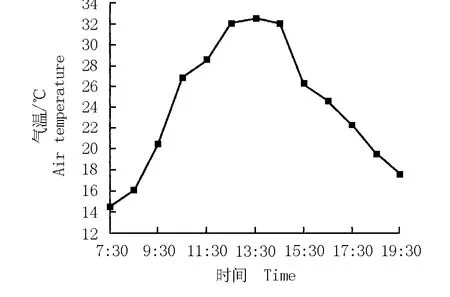
圖9 華山新麥草生境中氣溫的日變化曲線Fig.9 Daily change curve of air tem perature in habitat of Psathyrostachys huashanica Keng ex P.C.Kuo
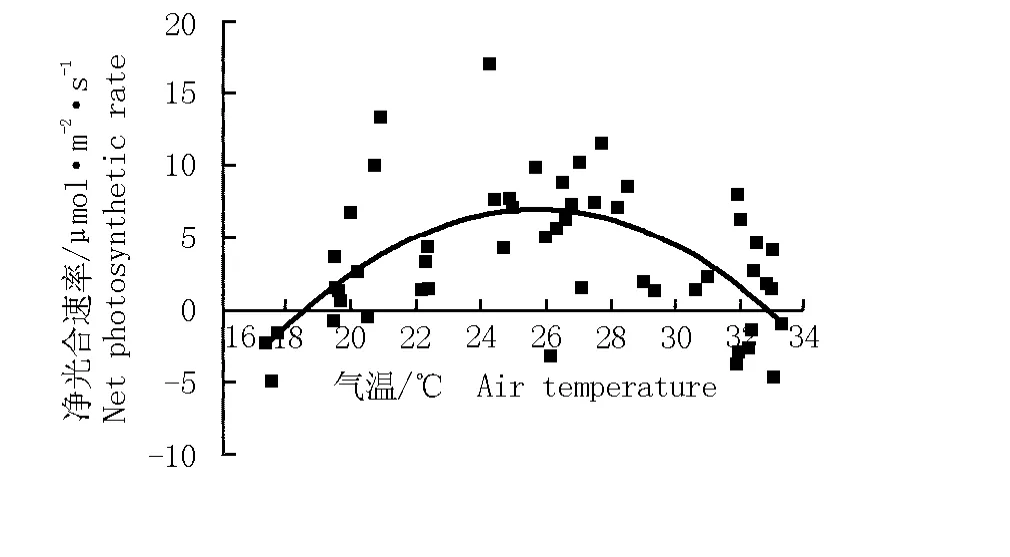
圖10 華山新麥草凈光合速率對氣溫的響應曲線Fig.10 Response curve of net photosynthetic rate of Psathyrostachys huashanica Keng ex P.C.Kuo to air temperature
3 討論和結論
研究結果表明:華山新麥草凈光合速率的日變化曲線共存在3個峰值且數值均較小,分別為6.5、6.2和9.0μmol·m-2·s-1。灰綠型羊草〔Aneurolepidium chinense(Trin.)Kitag.〕的凈光合速率日變化曲線有2個峰值,分別為23.2和17.3μmol·m-2·s-1[13];小麥(Triticum aestivum L.)的最大凈光合速率為27.7 μmol·m-2·s-1[14];水稻(Oryza sativa L.)的最大凈光合速率為23.6μmol·m-2·s-1[15],均高于華山新麥草的凈光合速率。產生這一現象的原因有二:一是實驗過程中氣候干旱,華山新麥草植株處于水分脅迫狀態,因而凈光合速率的測定值均偏低;二是與華山新麥草本身的生理代謝特性有關。岳明等[5]的研究結果表明:華山新麥草的生存競爭能力較弱。本研究結果從凈光合速率角度也印證了這個結論。
華山新麥草的凈光合速率具有“午降”現象,“午降”時凈光合速率的最低值為2.5μmol·m-2·s-1,明顯低于灰綠型羊草“午降”時凈光合速率的最低值(6.0μmol·m-2·s-1)[13]。推測華山新麥草的凈光合速率在12:30左右產生“午降”現象,可能的原因為:此時的光合有效輻射強度達到全天的最大值,空氣和葉片溫度均較高,致使氣孔關閉,進而抑制植株的光合作用;此時植株體內的光合產物已積累到一定程度,更重要的是葉內水分含量在中午最少,使植株的光合作用受到水分脅迫的影響;另外,CO2自根向地上部分的運輸量在中午增加、過度光照引起光氧化反應以及呼吸作用的日變化所引發的光合作用的變動[16]等都可能與華山新麥草凈光合速率的“午降”現象有關。
研究結果表明:華山新麥草葉片的蒸騰速率一直在1.7 mmol·m-2·s-1以下,明顯低于灰綠型羊草蒸騰速率的“雙峰”值(6.1和7.2 mmol·m-2·s-1)[13],這可能與華山新麥草葉片氣孔阻力較大有關。
根據華山新麥草凈光合速率對光合有效輻射強度的擬合方程,計算得出華山新麥草的光補償點和光飽和點分別為1.1和531.5μmol·m-2·s-1。在光飽和點以下,光合有效輻射強度是華山新麥草光合作用的主要限制因素,其凈光合速率隨著光合有效輻射強度的增強而提高;但光合有效輻射強度達到光飽和點后,光合有效輻射強度就不再是華山新麥草光合作用的主要限制因素。植物葉片的光飽和點與光補償點反映了植物對光照條件的要求,分別體現了對強光和弱光的利用能力。光補償點較低、光飽和點較高的植物對光照強度的適應范圍較廣;而光補償點較高、光飽和點較低的植物則對光照強度的適應范圍較窄[17]。華山新麥草的光補償點較低而光飽和點較高,顯示其具有很強的喜光性,且對光照強度的適應范圍較廣。
氣孔阻力能夠反映氣孔開啟變化的情況,直接影響植物的光合作用和蒸騰作用[18]。華山新麥草的氣孔阻力變化非常大,從早上8:30的15 m2·s·mol-1迅速增大到11:30的74 m2·s·mol-1,嚴重阻礙了光合作用的順利進行。禾本科植物的氣孔與其他植物不同,不但在白天仍處于半開狀態,而且對光的反應很敏感。由此可以認為:在陰天和雨天或是早晨、晚上,禾本科植物的光合作用不僅受到光照條件的限制,同時還受到氣孔開啟程度的制約[19]。因此,氣孔阻力是對華山新麥草凈光合速率和蒸騰速率具有負效應的重要因子。
綜合分析結果表明:華山新麥草的凈光合速率與氣孔阻力、光合有效輻射強度、葉溫和氣溫等因素均有很大的關系,其中,葉溫、光合有效輻射強度和氣溫對凈光合速率的影響曲線基本為“拋物線”型,氣孔阻力對凈光合速率的影響則表現為“隨氣孔阻力的增大凈光合速率持續降低”的趨勢。較大的氣孔阻力阻礙了華山新麥草的光合作用,使其不能有效合成代謝所需的大量營養物質,以維持自身的正常生命活動,這可能是華山新麥草瀕臨滅絕的主要生理原因之一。
[1]中國科學院西北植物研究所.秦嶺植物志:第1卷第1分冊[M].北京:科學出版社,1976:99.
[2]孫根樓,顏 濟,楊俊良.普通小麥和新麥草屬間雜種的產生及細胞遺傳學研究[J].遺傳學報,1992,19(4):322-326.
[3]盧寶榮.小麥族遺傳資源的多樣性及其保護[J].生物多樣性,1995,3(2):63-68.
[4]王 麗,趙桂仿.華山新麥草胚和胚乳的發育研究[J].西北植物學報,2002,22(4):786-790,T005-T006.
[5]岳 明,張林靜,馬 凱,等.華山新麥草瀕危原因及種群繁殖對策[J].生態學報,2001,21(8):1314-1320.
[6]趙利鋒,李 珊,潘 瑩,等.華山新麥草自然居群沿海拔梯度的遺傳分化[J].西北植物學報,2001,21(3):391-400.
[7]陳德祥,李意德,駱土壽,等.海南島尖峰嶺熱帶山地雨林下層喬木中華厚殼桂光合生理生態特性的研究[J].林業科學研究,2003,16(5):540-547.
[8]劉志龍,方建民,虞木奎,等.三種林-茶復合林分中環境因子和茶的光合特征參數的日變化規律[J].植物資源與環境學報,2009,18(2):62-67.
[9]譚會娟,周海燕,李新榮,等.珍稀瀕危植物半日花光合作用日動態變化的初步研究[J].中國沙漠,2005,25(2):262-267.
[10]王建麗,張永亮,朱占林,等.雜花苜蓿葉片光合生理生態特性[J].草地學報,2006,14(2):138-141.
[11]賈宏濤,趙成義,盛 鈺,等.干旱地區紫花苜蓿光合日變化規律研究[J].草業科學,2009,26(7):56-60.
[12]鄒學校,馬艷青,張竹青,等.環境因子對辣椒光合與蒸騰特性的影響[J].植物資源與環境學報,2005,14(4):15-20.
[13]王德利,王正文,張喜軍.羊草兩個趨異類型的光合生理生態特性比較的初步研究[J].生態學報,1999,19(6):837-843.
[14]王煥忠,李雁鳴,張建平,等.不同熟期小麥品種光合性能的初步研究[J].河北農業大學學報,1998,21(2):1-5.
[15]曹樹青,翟虎渠,張紅生,等.不同類型水稻品種葉源量及有關光合生理指標的研究[J].中國水稻科學,1999,13(2): 91-94.
[16]徐 江,朱建雯,張巨松,等.田間棉花葉片呼吸作用對光合日變化的影響[J].新疆農業大學學報,1997,20(4):1-4.
[17]武維華.植物生理學[M].1版.北京:科學出版社,2003: 168-170.
[18]劉貞琦,伍賢進,劉振業.土壤水分對煙草光合生理特性影響的研究[J].中國煙草學報,1995,2(3):44-49.
[19]戶刈義次,山田登,杉山直儀,等.作物的生理生態[M].崔繼林,劉中宣,鄭丕堯,等,譯.北京:科學出版社,1962:27-56.
