新蚜蟲癘霉感染致病蚜蟲的空間分布特征及其理論抽樣技術
2012-02-28 07:47:56和淑琪杜廣祖李正躍
植物保護 2012年2期
關鍵詞:理論
陳 斌, 和淑琪, 杜廣祖, 李正躍
(云南農業大學植物保護學院,生物多樣性控制病蟲害教育部重點實驗室,生物多樣性控制作物病蟲害應用技術國家工程研究中心,昆明 650201)
新蚜蟲癘霉[Pandor a neoaphidis (Remaudière& Hennebert)Hu mber](蟲霉目:蟲霉科)是分布范圍廣、流行性強的蚜科專化性蟲生真菌[1-2],常引發桃蚜的流行病,在桃蚜種群自然控制中具有重要作用[2-3],也是云南省昆明地區十字花科蔬菜蚜蟲病原真菌的主要種類[4-5]。國內外已對新蚜蟲癘霉的分布與生物學[5-7]及其應用[8-9]等方面進行了研究報道。邱麗絢等[10]對小白菜田蘿卜蚜(Lipaphis er ysi mi)(原誤定為菜縊管蚜Rhopal osiphu m pseudobr assicae)新蚜蟲疫霉的空間分布進行了研究,而對于發生流行普遍的新蚜蟲癘霉在甘藍田桃蚜種群的空間分布型及田間調查理論抽樣技術尚未見報道。生物種群的空間分布型是生物在特定空間中存在特征,研究蟲生真菌在甘藍田桃蚜種群中的空間分布特征,對深入研究其流行規律、準確預測預報及合理利用具有重要意義。因此,本研究立足于云南特殊的地理氣候環境條件下桃蚜及新蚜蟲癘霉在十字花科蔬菜中發生流行普遍的實際,系統調查,采用空間分布特征指標分析其空間分布格局,并對其抽樣技術進行了擬合,為深入研究和利用該蟲生真菌提供理論依據。
1 材料和方法
1.1 調查地點及調查方法
調查地點:設在云南中部昆明市嵩明縣蔬菜基地,N25°05′~25°28′,E102°40′~103°20′。屬典型的溫帶、暖溫帶和北亞熱帶混合型氣候,年均氣溫14℃,全年分為雨季和旱季,每年6-8月份為雨季,降水充沛,濕度較大,9月到第2年5月為旱季,濕度較低。主要種植蔬菜、玉米、馬鈴薯和水稻等作物。
調查方法:2009年1-12月,選擇種植管理水平基本一致的甘藍田,每塊田隨機抽樣10株,檢查計數每株被感染桃蚜數量。每月上、中、下旬各調查一次,每次調查10塊田。統計每塊田桃蚜被新蚜蟲癘霉感染的數量,用Taylor冪法則測定其聚集度,用Iwao回歸方程測定其在不同時間的空間分布型,分析其空間分布特征。
1.2 空間分布型測定
1.2.1 聚集度指標檢驗
按照 叢 生 指 標[11]、m*/m 指 標[12]、擴 散 系 數C[13]、Ca指標[14]、負二項式分布 K 指標進行檢驗,各指標的計算公式如下:
叢生指標I:I=S2/m-1,當I>0時為聚集分布;當I=0時為隨機分布;當I<0時為均勻分布。
m*/m指標:即平均擁擠度與其平均密度之比,當m*/m>1時為聚集分布;當m*/m=1時為隨機分布;當m*/m <1時為均勻分布。
擴散系數C:C=S2/m,當C>1時為聚集分布;當C=1時為隨機分布;當C<1時為均勻分布。
Ca指標:Ca=(S2/m-1)/m,當Ca>0時為聚集分布;Ca=0時為隨機分布;Ca<0時為均勻分布。
Water負二項式分布K 指標:K=m2/(S2-1)。當K<0時為均勻分布;當K→+∞時為隨機分布;當0<K<8時為聚集分布。
1.2.2 線性回歸方程檢驗
Iwao m*-m 檢驗[15]:m*=α+βm,其中,α為分布的基本成分按大小分布的平均擁擠度,β為基本成分的空間分布圖式。當α>0時,個體間相互吸引;當α=0時,分布的基本成分是單個個體;當α<0時,個體間相互排斥。當β>1時為聚集分布;β=1時為隨機分布;β<1時為均勻分布。
Tayl or冪法則檢驗[16]:lg S2=l g(α)+b l g(m)。當b>1時呈聚集分布;b=1時呈隨機分布;b<1時呈均勻分布。
1.2.3 聚集原因分析
用Blackit h聚集均數λ[13]:λ=×γ,其中γ為自由度為2 Kc、概率為0.5時的χ2分布值。當λ>2時,其聚集原因是由病原體本身的習性和環境條件或由其中一個因素所引起;當λ<2時,其聚集原因則是由于環境條件所引起。
1.3 抽樣技術
2+β-1),式中:n為理論抽樣數;x為平均密度(頭/樣方);t為一定置信度下t分布值。取概率保證值t=1.96,允許誤差 D=0.1、0.2、0.3,0.4,0.5;α、β為Iwao回歸式中的參數。建立理論抽樣模型,計算新蚜蟲癘霉感染桃蚜不同蟲口密度下所需的理論抽樣數。
1.4 數據統計方法
叢生指標、m*/m指標、擴散系數C、Ca指標、負二項式分布K指標均在DPS統計軟件[17]進行,被感染蚜量間的方差值采用LSD極小顯著差異法進行統計。
2 結果與分析
2.1 新蚜蟲癘霉空間分布型
2.1.1 新蚜蟲癘霉空間分布的聚集度指標
根據1-12月各調查田塊新蚜蟲癘霉感染桃蚜的平均感染蚜量(X),方差(V)及平均擁擠度()分析結果,其平均擁擠度為(3.889±0.685)~(58.654±4.560),I指標為0.643~17.978,/m 指標為1.429~2.746 7,Ca指標為0.399~1.738,擴散系數C為1.633~18.968,K 指標為1.242~8.297。根據種群聚集分布的指標值I>0,/m>1,Ca>0,C>1,0<K<8的要求,該研究中新蚜蟲癘霉空間分布的/m指標值、Ca指標、擴散系數C和K 指標值均符合聚集分布特征參數值,表明新蚜蟲癘霉的空間分布型呈聚集分布格局。
2.1.2 新蚜蟲癘霉空間分布特征
將新蚜蟲癘霉感染桃蚜的平均密度與方差間的關系用上述冪函數公式進行擬合,結果為α=0.12,b=1.36,經卡方(χ2)檢驗,χ2=76.85<(359)=124.34,故擬合有效。說明新蚜蟲癘霉在甘藍上桃蚜種群中的分布為聚集分布。
2.1.3 新蚜蟲癘霉負二項分布的公共K值
由于α=0.12>0,b=1.36>1,因此新蚜蟲癘霉在甘藍田間的分布型為具公共K值的負二項分布,其個體間互相吸引,負二項分布的公共K值為2.87,卡方值=124.34(df=359),在空間以個體群結構的形式存在,且個體群間也呈聚集分布,個體群平均大小指數(即m+1)也與平均密度呈極顯著正相關。
2.2 不同季節新蚜蟲癘霉空間分布特征
2.2.1 聚集度指標
不同季節新蚜蟲癘霉空間分布的結果表明,1-12月其平均擁擠度為(3.889±0.685)~(58.654±4.56),均滿足聚集分布特征值。根據種群聚集分布的指標值I>0/m>1,Ca>0,C>1,0<K<8的要求,本研究中/m指標值、Ca指標、擴散系數C和K指標值均符合聚集分布特征參數值(表1)。表明新蚜蟲癘霉在甘藍田呈聚集分布格局。
2.2.2 空間分布型特征

表1 不同季節新蚜蟲癘霉的聚集度指標

表2 不同時間新蚜蟲癘霉空間分布型特征1)
2.3 聚集原因分析
Blackith(1961)[13]提出,昆蟲種群空間聚集原因可通過公式λ=×γ(式中:γ為自由度為2 Kc,概率為0.05時的χ2分布值)進行計算和評價。即當λ<2時,種群聚集的原因可能是由于某些環境因素作用的結果;當λ>2時,其聚集原因可能由于病原菌本身的特性所引起,也可能由于昆蟲本身的特性與環境的異質性兩個因素所引起。

依據上式,當λ=2時,m=0.94。因此,當新蚜蟲癘霉感染后罹病桃蚜的平均密度<0.94頭/株時,其在甘藍上的聚集與當時病原菌自身的特性有關;而當平均密度≥0.94頭/株時,新蚜蟲癘霉在甘藍田內的聚集分布除與當時病原菌的特性有關外,還與其寄主蚜蟲本身的聚集行為有關。
2.4 新蚜蟲癘霉空間抽樣技術
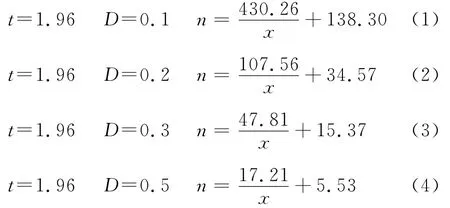
通過以上理論模型,計算出甘藍田新蚜蟲癘霉在不同罹病桃蚜種群密度下所需的理論抽樣數(表4)。
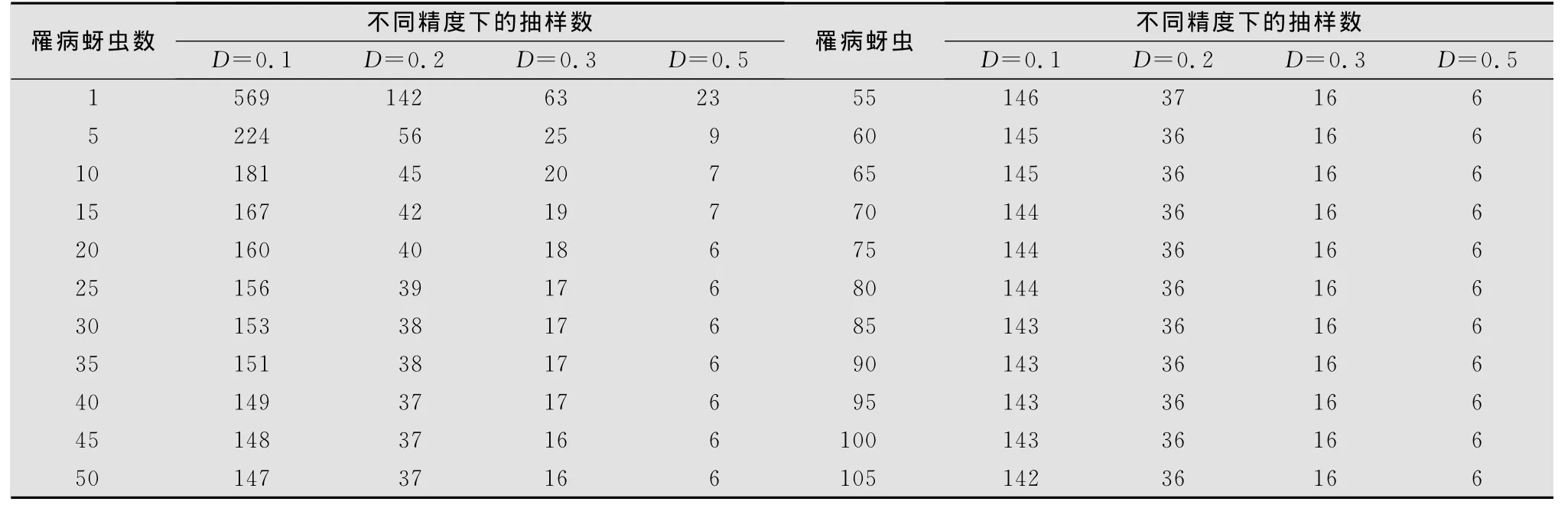
表4 不同罹病桃蚜密度下的理論抽樣數
從表4結果看出,當新蚜蟲癘霉感染的桃蚜平均密度≤1頭/株時,取概率保證值t=1.96,允許誤差D=0.1、0.2、0.3和0.5時,其理論抽樣數依次為569、142、63株和23株。當平均密度在5頭時,所需抽樣數分別為224、56、25、9株。而當平均密度在10頭以上時,其理論抽樣數隨蟲口密度的升高而平穩下降,蟲口密度越高,理論抽樣數越少。綜合比較4個模型,在新蚜蟲癘霉密度較低,即低于每樣方5頭時,以模型(4)較為適宜,在蟲口密度較高,即每樣方10頭以上時,以模型(3)較宜。
綜合近年來在云南菜區新蚜蟲癘霉病害流行情況,在6-8月份,對該病害的總體理論抽樣以模型(3)理論抽樣為宜,即抽查20株左右樣方;而在其他季節,每樣方蟲口密度5頭以下時,抽查30株左右的樣方。
3 討論
根據昆蟲空間分布評價的5項聚集度指標及Taylor冪法則與Iwao回歸方程,新蚜蟲癘霉在甘藍田內桃蚜種群中呈聚集分布中的負二項分布,該結果與邱麗絢等[10]報道的蘿卜蚜種群中的新蚜疫霉的空間分布研究結果一致。此外,根據桃蚜在甘藍田的空間分布特點[18],新蚜蟲癘霉的分布型與寄主昆蟲的分布型一致,該結果也與Royal et al.[8]提出的新蚜蟲癘霉為密度依賴型病原菌結果及關雄等[19]報道的柑橘葉片上粉虱座殼孢菌在柑橘樹冠上的空間分布格局與柑橘粉虱在柑橘上的空間分布格局一致的結論相似,這也許正是新蚜蟲癘霉病害發生流行性強的重要原因之一。另外,根據一年中新蚜蟲癘霉感染桃蚜的聚集強度的季節性變化規律,6-10月份平均擁擠度為全年內最高時段,這與劉樹生等[18]報道的桃蚜在一年內聚集高峰期一致,同時也與昆明地區近年來桃蚜種群中新蚜蟲癘霉病害的發生流行實際一致[4-5]。
寄主昆蟲的習性也是影響昆蟲病原微生物聚集特性的重要因素之一[10],新蚜蟲癘霉能寄生桃蚜等多種蚜蟲[2],在昆明地區多種蚜蟲中均有發現[3,5],而甘藍上除桃蚜外,還有大量的甘藍蚜,但在平時的調查中發現甘藍蚜感染率很低[5],因此本研究中僅采用甘藍上被新蚜蟲癘霉感染的桃蚜的密度來分析新蚜蟲癘霉的空間分布及理論抽樣技術,該結果與關雄等[19]根據柑橘葉片上粉虱座殼孢菌子座數進行取樣、分析粉虱座殼孢菌在柑橘樹冠上的空間分布格局的結果相似,由此本研究取樣方法及其結果可以反映該菌在甘藍田的空間分布特征。
新蚜蟲癘霉的空間聚集分布的基本單位為個體群,當平均感染蚜量≥0.94頭/株時,聚集分布的原因除與當時病原菌的特性有關外,還與其寄主蚜蟲本身的聚集行為有關,而當新蚜蟲癘霉對桃蚜的平均感染蚜量<0.94頭/株時,聚集與當時病原菌自身的特性有關,這種特性可能是由于新蚜蟲癘霉可通過有翅蚜的遷飛擴散傳播[5-7]、捕食性瓢蟲的活動取食來擴散和傳播[7]特性及該菌分生孢子較強的彈射能力,使該菌在呈聚集分布的桃蚜種群中呈現典型的聚集分布。
[1] Hu mber R A.Fungi:Identification[M]∥Lacey L A.Mannual of Techniques in Insect Pat hology.New Yor k:Academic Press,1997:153-185.
[2] 李增智主編.中國真菌志(第十三卷:蟲霉目)[M].北京:科學出版社,2000.
[3] 蒲蟄龍,李增智.昆蟲真菌學[M].合肥:安徽科學技術出版社,1996.
[4] 陳斌,李正躍,桂富榮,等.云南省昆明地區菜蚜新蚜蟲癘霉發生流行的規律[J].華中農業大學學報,2008,27(2):204-207.
[5] Chen B,Li Z Y,Feng M G.Occurrence of ento mopat hogenic f ungi on migratory alate aphids in Yunnan[J].Biocontrol,2008,53(2):317-326.
[6] Feng M G,Chen C,Chen B.Wide dispersal of aphid-pathogenic entomophthorales among aphids relies upon migratory alates[J].Envir on mental Ento mology,2004,6(5):510-516.
[7] Roy H E,Pell J K,Alderson P G.Targeted dispersal of the aphid pat hogenic f ungus Er ynia neoaphidis by the aphid predator Coccinell a septempunctata [J].Biocontrol Science and Technology,2001,11(1):99-110.
[8] Shah P A,Aebi M,Tuor U.Infection of Macrosiphum euphorbiae with mycelial preparations of Er ynia neoaphidis in a greenhouse trial[J].Mycological Research,2000,104(6):645-652.
[9] Wilding N,Mardell S K,Brobyn P J.Introducing Er ynia neoaphidis into a field population of Aphis f abae:for m of the inoculu m and effectof irrigation[J].Annals of Applied Biology,1990,108(1):373-385.
[10]邱麗絢,黃耀堅,鄭本暖.新蚜疫霉流行病的空間分布型[J].昆蟲天敵,1989,11(4):1-4.
[11]David F N,Moore P G.Notes on contagious distribution in plant population[J].Annals of Botany,1954,18(1):47-53.
[12]Lloyd M.Mean cro wding[J].Jour nal of Ani mal Ecology,1967,36(1):1-30.
[13]徐汝梅.昆蟲種群生態學[M].北京:北京師范大學出版社,1987.
[14]Kuno E.Use of the regression of mean crowing on mean density for esti mating sa mple size and the transfor mation of data for analysis of variance[J].Researches on Population Ecology,1968,10(2):210-214.
[15]Iwao S.A new regression met hod for analyzing the aggregation pattern of ani mal populations[J].Researches on Population Ecology,1968,10(1):1-20.
[16]Taylor L R.Aggregation variance and the mean[J].Nature,1961,189(4766):732-735.
[17]唐啟義,馮明光.實用計算機統計分析及其計算機處理平臺[M].北京:中國農業出版社,2002.
[18]劉樹生,汪信庚,吳曉晶,等.甘藍和白菜上桃蚜種群的空間格局及其時序動態[J].昆蟲學報,1996,39(2):158-164.
[19]關雄,高日霞.粉虱座殼孢菌的空間分布型及其抽樣技術[J].福建農學院學報,1991,20(1):29-33.
猜你喜歡
當代陜西(2022年5期)2022-04-19 12:10:18
新世紀智能(數學備考)(2021年9期)2021-11-24 01:14:28
湘潮(上半月)(2021年4期)2021-07-20 08:05:28
汕頭大學學報(自然科學版)(2020年4期)2020-12-14 07:05:00
讀與寫·教育教學版(2017年10期)2017-11-10 22:28:57
大電機技術(2017年3期)2017-06-05 09:36:02
廣州大學學報(社會科學版)(2016年1期)2016-06-24 09:46:02
區域經濟評論(2016年2期)2016-05-17 05:06:43
學習月刊(2015年21期)2015-07-11 01:51:44
社會生活探索(2013年0期)2013-10-24 03:44:40
