苦參堿對小鼠坐骨神經損傷后脊髓運動神經元的影響
2012-08-20 10:40:38王永安劉浩宇尹維田
中國實驗診斷學 2012年1期
曹 劍,王永安,魏 壯,劉浩宇,尹維田
(1.赤峰市醫院,內蒙古 赤峰024000;2.吉林大學中日聯誼醫院)
本文對坐骨神經損傷BALB/c小鼠應用苦參堿,通過對坐骨神經相應脊髓節段gap-43的測定和神經LFB染色的觀察,探討苦參堿對周圍神經損傷后的修復和再生方面的作用。
1 材料與方法
1.1 實驗動物和分組
健康成年雄性BALB/c小鼠160只(體重25±2g,由吉林大學基礎醫學院實驗動物中心提供)單側坐骨神經離斷后,常溫飼養,普通飲食,自由飲水。隨機分為生理鹽水空白對照組、高劑量給藥組、中劑量給藥組和低劑量給藥組,其中實驗組動物又根據術后存活時間隨機分為12h,24h,3d,5d、7d、2 w,4w,8w組,以上各組每組10只動物。
1.2 動物模型制備
坐骨神經損傷模型的制作:BALB/c小鼠經1%硫噴妥鈉100mg/kg腹腔麻醉,俯臥位固定,無菌條件下單側下肢股后部縱切口約2cm,梨狀肌下暴露坐骨神經,用玻璃分針鈍性小心分離坐骨神經主干和周圍組織,在坐骨結節下0.5cm處完全離斷坐骨神經,以11/0顯微縫線12倍顯微鏡下吻合坐骨神經,分層縫合肌肉和皮膚。
1.3 給藥方法和劑量
采取灌胃方法給藥。參照臨床應用苦參堿原藥劑量,等效換算為腹腔注射用劑量[1],將此數值作為給藥的中等劑量,并以等比數列量確定高低劑量組用量。最終各組給藥量為,高劑量組10mg/kg/d,中劑量組5mg/kg/d,低劑量組2.5mg/kg/d,取相應劑苦參堿以生理鹽水溶解后腹腔注射。對照組給予同等體積生理鹽水。連續給藥至處死。
1.5 取材及標本制備
取每組動物5只分別在相應的時間點用1%硫噴妥鈉100mg/kg腹腔麻醉后,采用脊柱后方正中切口咬骨鉗咬開椎管,切取損傷側L4-L6段的脊髓,標記后迅速浸置于液氮中備用。
取每組另外5只在相應時間點切取坐骨神經自吻合口(包括吻合口在內)向遠端0.6-0.7cm神經干,10%中性甲醛固定標本72小時以上,經酒精逐級脫水后石蠟包埋。
1.6 檢測方法
1.6.1 Real-time PCR 法檢測 首先進行引物的合成,用 Beacon designer 7軟件設計 gap-43、以GAPDH為引物(表1),經檢索Blast驗證特異性。分別取各時間點的L4-6脊髓節段組織,采用Trizol試劑提取總RNA,以提取的總RNA為模板進行反轉錄制備cDNA庫。以cDNA庫為模板,PCR擴增L4-6脊髓節段組織的gap-43,每個反應體系中均加入GAPDH的一對引物作為PCR擴增的內參,反應條件:95℃30s-58℃60s-72℃60s40個循環。
1.6.2 髓鞘固蘭染色 切取坐骨神經自吻合口(包括吻合口在內)向遠端0.6-0.7cm神經干,10%中性甲醛固定標本72小時以上,經酒精逐級脫水后石蠟包埋,制片行HE染色。觀察神經的基本結構及有無細胞增生、炎細胞浸潤等改變,初步篩選特染對象。切片脫蠟后,用LFB液60℃浸泡12小時;95%酒精浸泡5min;加入0.05%碳酸鋰15s;而后70%酒精洗滌完畢蒸餾水洗滌,脫水透明封片。

表1 Beacon designer 7軟件設計GAP43、GAPDH引物
2 結果
2.1 Realtime-PCR分析結果
Gap-43mRNA在正常大鼠坐骨神經組織少量表達,當坐骨神經損傷后,相應脊髓節段組織gap-43mRNA的含量上升,從損傷后12h開始到5d呈現逐漸增加的趨勢,各組間差異性明顯,其中高中劑量組的增多量明顯低于低劑量和對照組。見圖1。2w后gap-43在高中劑量組呈持續表達。其中,中劑量組明顯促進了gap-43的表達。
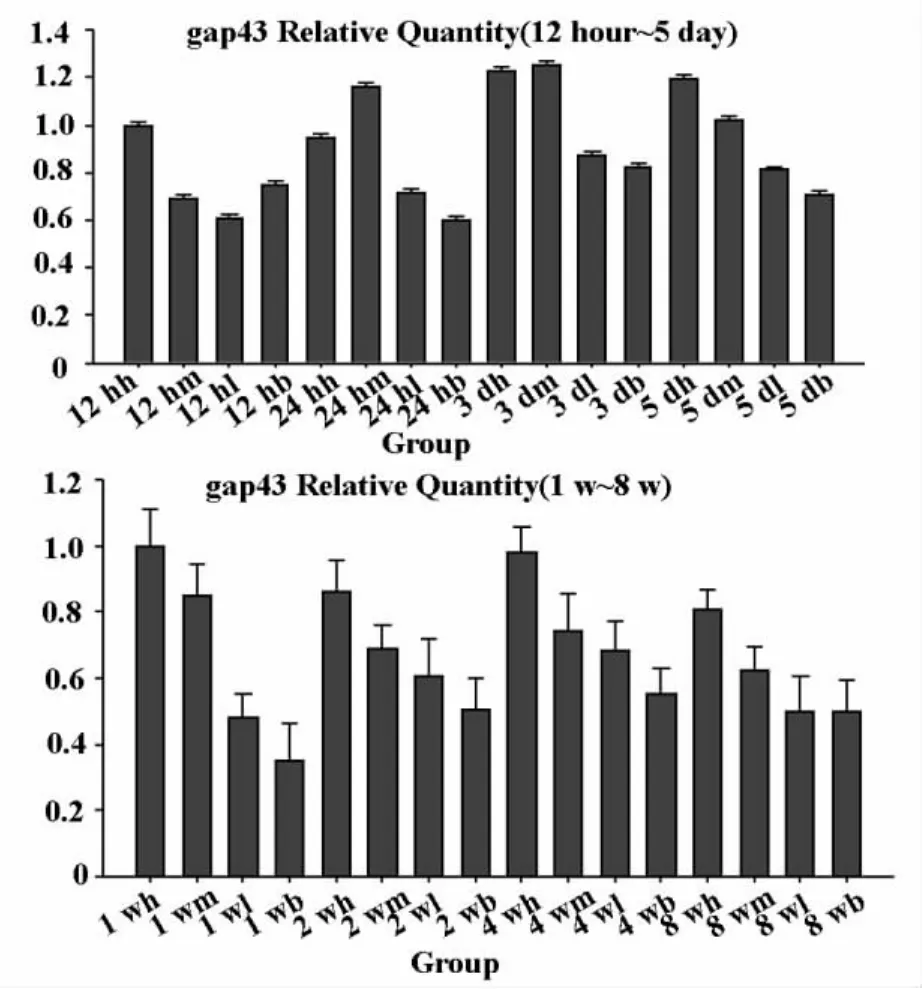
圖1 各時間點gap-43的mRNA表達
2.2 LFB染色結果
LFB染色后,髓鞘呈天藍色,軸突不著色,背景為白色。術后8周染色結果見圖2。高、中劑量組髓鞘形狀規則,厚度較為均一,輪廓明顯且周邊纖維組織增生不明顯;低劑量組髓鞘形狀及厚度較不規則,但輪廓尚明顯,束間可見纖維結締組織增生;對照組鞘形狀更加不規則,纖維結締組織增生明顯。
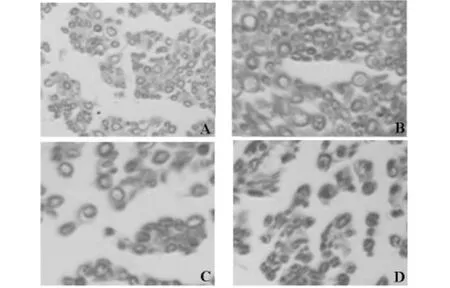
圖2 造模后各劑量組髓鞘固藍染色結果
綜合以上兩組指標的檢測,不同時相小鼠L4-6脊髓節段組織中gap-43mRNA的變化,以及髓鞘LFB染色的趨勢具有一致性。
3 討論
苦參堿是廣泛存在于豆科植物苦參(sophora flavescents Ait)、苦豆子(S.alopecuroides L)及廣豆根(S.subprostrate chunet T.Chenk)等中草藥中的有效成份。近年來對苦參堿的研究較為廣泛,它的化學式為C15H24N20,屬于四環的喹嗪啶類,分子骨架可看作2個喹嗪啶環的雜體。苦參堿具有多種藥理作用,對中樞神經系統可引發中樞神經麻痹,可防治腦水腫和鎮靜、降溫作用;對心血管系統,可增強心肌收縮力,抗心律失常、抗動脈粥樣硬化;對消化系統,可防治肝炎、抗肝纖維化,利膽、止瀉,保護胃腸黏膜;對呼吸系統,具有祛痰平喘作用。此外,還有抗病毒、抗寄生蟲、抗炎、免疫調節及抗瘢痕形成等作用[2-4]。而對其促進神經損傷后修復的研究較少。生長相關蛋白gap-43是一種胞膜磷酸蛋白質,是膜快速轉運蛋白,與神經元生長發育、突觸形成及神經可塑性密切相關[5]。在膜信號轉導系統中,G蛋白作為信號轉導因子和放大器處于核心地位,gap-43可調控G蛋白[6]。這種作用是通過gap-43的可結合到膜內表面的氨基末端完成的,作為純化的蛋白質,gap-43起一個鳥苷酸釋放蛋白的作用。它可提高從G0蛋白釋放GDP、提高GTP激酶活性及與GTP的結合[7]。我們的研究發現:Realtime PCR的檢驗結果表明坐骨神經損傷后,相應脊髓節段的gap-43被激活并大量表達,應用苦參堿后高中劑量組的gap-43表達明顯高于低劑量組及對照組,并與2周后持續表達。坐骨神經損傷后應用苦參堿可以對gap-43的活化起到持續促進作用,促進了神經的再生。
[1]de Oliveira Luiz FC,Edwards Howell GM,Velozo Eudes S,Nesbitt M.Vibrational spectroscopic study of brazilin and brazilein,the main constituents of brazilwood from Brazil[J].Vib Spectrosc,2002,28:243.
[2]Kim JK.Illustrated natural drugs encyclopedia[M].Seoul;Namsandang Publishers;1989:79.
[3]黃繼漢,黃曉暉,陳志揚,等.藥理試驗中動物間和動物與人體間的等效劑量換算[J].中國臨床藥理學與治療學,2004,9(9):1069.
[4]劉 梅,劉雪英,程建峰.苦參堿的藥理進展[J].中國中藥雜志,2003,28(9):801.
[5]Benowitz L I,Routtenberg A.GAP-43:an intrinsic determinant of neuronal development and plasticity[J].Trends Neurosci,1997,20 (2):84.
[6]Fishman MC.GAP243:Putting constraints on neuronal plasticity[J].Perspective in developmental neurobiology,1996,4:193.
[7]Dani JW,Armstrong DM,Benowitz L I.Mapping the development of the rat brain by GAP-43Immunocytochemistry[J].Neuroscience,1991,40(1):277.
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
課堂內外·初中版(科學少年)(2023年10期)2023-12-10 00:43:06
全科護理(2022年10期)2022-12-26 21:19:15
中國合理用藥探索(2022年1期)2022-11-26 00:22:32
鄉村科技(2021年33期)2021-03-16 02:26:54
國際放射醫學核醫學雜志(2021年10期)2021-02-28 08:41:58
學苑創造·A版(2020年9期)2020-10-13 09:41:02
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
中國合理用藥探索(2014年11期)2014-03-11 20:30:20
