日本七鰓鰻pcna基因克隆、生物信息學(xué)分析及真核表達(dá)
2012-09-18 09:16:10龔興旺黃金莎李慶偉
大連大學(xué)學(xué)報(bào) 2012年6期
逄 越,龔興旺,黃金莎,李慶偉,*
(1. 大連大學(xué) 生命科學(xué)與技術(shù)學(xué)院,遼寧 大連 116622;2. 遼寧師范大學(xué) 生命科學(xué)學(xué)院, 遼寧 大連 116029)
日本七鰓鰻(Lampetra japonica)是現(xiàn)存最古老的脊椎動(dòng)物代表。由于七鰓鰻是聯(lián)系無脊椎動(dòng)物與脊椎動(dòng)物之間的重要階元,因此在遺傳信息方面,它必定印記了無脊椎動(dòng)物的進(jìn)化歷史,同時(shí)作為脊椎動(dòng)物最直接的祖先,又為脊椎動(dòng)物的起源與進(jìn)化提供豐富的遺傳信息[1-3]。增殖細(xì)胞核抗原(proliferating cell nuclear antigen,PCNA)是一種相對(duì)分子質(zhì)量為36KD的酸性蛋白,已經(jīng)證實(shí)它的表達(dá)是同細(xì)胞周期中S期的細(xì)胞增殖狀態(tài)相關(guān)聯(lián),直接參與 DNA的合成和復(fù)制,是反映細(xì)胞增殖狀態(tài)的特異性核抗原[4.5]。1987年 Bravo等[6]指出PCNA是DNA多聚酶δ的輔助蛋白,DNA合成必不可少的因子,PCNA在細(xì)胞核內(nèi)合成,并存在于細(xì)胞核內(nèi)。PCNA在細(xì)胞周期的不同階段表達(dá)有所不同,在G0~G1期細(xì)胞中無明顯表達(dá),G1晚期,其表達(dá)大幅度增加,S期達(dá)到高峰,G2~M期明顯下降,其量的變化與DNA合成一致。因而檢測(cè)其在細(xì)胞中的表達(dá),可作為評(píng)價(jià)細(xì)胞增殖狀態(tài)的一個(gè)指標(biāo)[7-9]。本研究首次獲得七鰓鰻pcna基因并對(duì)其進(jìn)行生物信息學(xué)分析。除此之外,建立七鰓鰻PCNA與綠色熒光蛋白融合基因表達(dá)載體并成功表達(dá)融合蛋白。七鰓鰻pcna基因真核表達(dá)載體的成功構(gòu)建及表達(dá)為pcna基因在七鰓鰻中生物學(xué)功能研究奠定基礎(chǔ),也為七鰓鰻細(xì)胞株建立提供材料。
1 材料和方法
1.1 材料
實(shí)驗(yàn)材料:日本七鰓鰻2011年12月末采自黑龍江省松花江流域同江地區(qū)。選取健康、體表完整的個(gè)體,經(jīng)過 7d培養(yǎng)觀察,證實(shí)無病后用于實(shí)驗(yàn)。
菌株與質(zhì)粒:質(zhì)粒PGFP-N2、大腸桿菌DH5α菌株、Hela細(xì)胞系為本實(shí)驗(yàn)室保存。
主要試劑:限制性核酸內(nèi)切酶HindⅢ和BamHⅠ、T4DNA連接酶、pMD19-T Simple Vector、PrimeScript RT-PCR Kit、PrimeSTAR HS DNA聚合酶、瓊脂糖凝膠DNA純化試劑盒(Agarose GeI DNA Purification Kit)、質(zhì)粒小量提取試劑盒和DL2000、DL5000DNA 和 λ-HindⅢdigest DNA Marker均購自大連TaKaRa公司;Trizol試劑購自Invitrogen公司;Xfect Transfection Reagent購自Clontech公司;DMEM高糖、FBS、DPBS、卡那霉素、Typan blue購自于 Hyclone公司;Sodium Pyruvate購自 Invitrogen公司;Trypsin-EDTA Solution購自Sigma公司。
1.2 方法
1.2.1 功能基因的EST比對(duì)及序列進(jìn)化分析
從本實(shí)驗(yàn)前期構(gòu)建的七鰓鰻肝臟cDNA文庫(2.1×106pfu/mL)中隨機(jī)挑選cDNA單克隆進(jìn)行表達(dá)序列標(biāo)簽(EST)測(cè)序,建立EST數(shù)據(jù)庫(約10077條EST序列)[10]。將EST序列在NCBI蛋白質(zhì)數(shù)據(jù)庫(SwissProt, GenBank, PDB)中進(jìn)行BLAST比對(duì)發(fā)現(xiàn)一條與高等脊椎動(dòng)物 pcna基因同源的序列。利用CLUSTAL X軟件(version 1.81)將七鰓鰻PCNA氨基酸序列同其它不同物種的氨基酸序列進(jìn)行比對(duì),并用MEGA(version 4.0)軟件做進(jìn)化樹分析。
1.2.2 總RNA的提取和cDNA的合成
取 50~100 mg七鰓鰻肝臟組織按照 Trizol試劑盒說明書提取總RNA,置于-80℃冰箱保存。逆轉(zhuǎn)錄反應(yīng)按照PrimeScript RT-PCR Kit(Code No:DRR014S)說明書操作進(jìn)行,以適量總RNA為模板,以O(shè)ligo(dT)為引物,逆轉(zhuǎn)錄反應(yīng)體系于30℃溫育10 min;42℃溫預(yù)30 min;95℃溫育5 min。最終獲得cDNA第一鏈。
1.2.3 Lj-pcna基因片段擴(kuò)增
參考七鰓鰻肝臟cDNA文庫中的EST序列,利用Primer Premier 5.0軟件設(shè)計(jì)如下引物,P1F:5'-ACACACGACAAACGCACTCGCCAA-3',P1R:5'-TTATGCAGCCTCTTCATCCTC-3'。以含有pcna全長基因的日本七鰓鰻第一鏈cDNA為模板,使用 PrimeSTAR HS DNA聚合酶(Code No:DR010S)擴(kuò)增Lj-pcna基因。反應(yīng)條件:94℃變性30 s,55℃退火30 s,72℃延伸1 min,循環(huán)35次;最后72℃延伸10 min。取出PCR產(chǎn)物10 mg/L瓊脂糖凝膠電泳檢測(cè),凝膠成像分析系統(tǒng)記錄檢測(cè)結(jié)果。
1.2.4 Lj-pcna基因克隆
使用TaKaRa Agarose Gel DNA Purification Kit Ver.2.0(Code No:DV805A)切膠回收PCR產(chǎn)物。將目的片段 Lj-pcna與 pMD19-T Simple Vector(Code No:D104A)連接,并導(dǎo)入大腸埃希菌 DH5α感受態(tài)菌中,提取重組質(zhì)粒 T-Lj-pcna送到大連寶生物工程有限公司進(jìn)行測(cè)序及序列分析。
1.2.5 真核表達(dá)載體的構(gòu)建
以T-Lj-pcna重組質(zhì)粒為模板,根據(jù)1.2.4所測(cè)序列設(shè)計(jì)如下引物,P2F:5'-CCCAAGCTT ATGTTCGAGGCACGCATCCT G-3',P2R:5'-CGCGGATCCTGCAGCCTCTTCATC CTCAAT-3'。在上下游引物序列中分別添加了HindⅢ和BamHⅠ酶切位點(diǎn)(下劃線部分)。PCR擴(kuò)增步驟如1.2.3所示。回收的PCR產(chǎn)物和真核表達(dá)載體分別用HindⅢ和BamHⅠ進(jìn)行雙酶切,酶切的Lj-pcna基因片段和質(zhì)粒PGFP-N2片段通過T4DNA連接酶16℃連接過夜,轉(zhuǎn)化到大腸埃希菌 DH5a感受態(tài)中,在含有 kanar抗生素(20 ug/mL)的LB固體培養(yǎng)基上篩選確認(rèn)陽性克隆菌。挑取陽性克隆菌落進(jìn)行擴(kuò)大培養(yǎng),進(jìn)而提取PGFP-N2-Lj-pcna重組質(zhì)粒,送至寶生物工程有限公司進(jìn)行測(cè)序及序列分析。
1.2.6 重組質(zhì)粒pGFP-N2-Lj-pcna轉(zhuǎn)染Hela細(xì)胞
用高糖 DMEM(含10% FBS)培養(yǎng)基培養(yǎng)HeLa 細(xì)胞,轉(zhuǎn)染前24 h,選擇生長狀態(tài)良好、細(xì)胞密度約70%的細(xì)胞接種12-well plate,去掉細(xì)胞培養(yǎng)皿中的培養(yǎng)液,使用5 mL DPBS清洗,去掉培養(yǎng)基后添加 1 mL Trypsin-EDTA Solution,37℃放置,待細(xì)胞完全消化;再添加 9mL新鮮培養(yǎng)基混勻細(xì)胞,添加Typan blue計(jì)數(shù),調(diào)整細(xì)胞濃度為2×105cells/mL,0.5 mL/well 在12-well plate中接種細(xì)胞;37 5% CO2℃ 條件下培養(yǎng)24 h;24 h后,觀察細(xì)胞生長狀態(tài)良好,細(xì)胞密度約70%,按照轉(zhuǎn)染試劑說明書進(jìn)行轉(zhuǎn)染實(shí)驗(yàn):使用Xfect Transfection Reagent (Clontech cat#631317),準(zhǔn)備2個(gè)1.5 mL tube,分別命名為Polymer和DNA,按如下方法添加試劑:DNA tube: 5uL PGFP-N2-Lj-pcna Plasmid (500 ng/uL) + 45 uL Xfect Reaction Buffer Polymer tube: 0.75 uL Xfect Polymer + 49.25 uL Xfect Reaction Buffer 分別震蕩混勻,之后將 Polymer tube的混合液添加到DNA tube中,中速震蕩混勻10 s,混合液室溫下放置15 min,之后將100 uL的混合液全量添加到細(xì)胞孔中,100 uL/well,前后左右搖蕩 12-well plate混勻,37℃放置培養(yǎng)4 h后,細(xì)胞孔中使用1mL 新鮮培養(yǎng)基換液,繼續(xù) 37℃培養(yǎng)24 h后在熒光顯微鏡下觀察熒光表達(dá)情況。
2 結(jié)果
2.1 Lj-pcna片段擴(kuò)增結(jié)果
從日本七鰓鰻肝臟組織中獲得 pcna基因,PCR產(chǎn)物經(jīng) 10 mg/L瓊脂糖凝膠電泳,在大約800bp處可見特異性擴(kuò)增條帶,見圖1。
2.2 七鰓鰻PCNA生物信息學(xué)分析
通過NCBI網(wǎng)站ORF搜索軟件分析Lj-pcna cDNA序列786bp,編碼261個(gè)氨基酸,蛋白分子量為28713Da,見圖2所示。在NCBI數(shù)據(jù)庫中獲得多個(gè)物種的PCNA氨基酸序列,構(gòu)建進(jìn)化樹(如圖3),結(jié)果顯示,日本七鰓鰻PCNA基因位于脊椎動(dòng)物PCNA基因的外群,且處于節(jié)肢動(dòng)物門、尾索動(dòng)物亞門及脊椎動(dòng)物亞門之間,符合物種進(jìn)化規(guī)律。
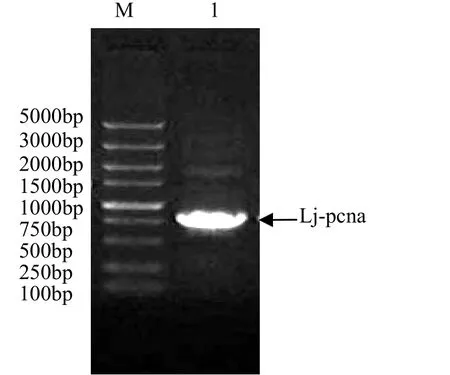
圖1 Lj-pcna目的基因

圖2 日本七鰓鰻 pcna的 ORF區(qū)及其編碼的氨基酸序列

圖3 PCNA系統(tǒng)發(fā)育樹
2.3 構(gòu)建pGFP-N2-Lj-pcna真核表達(dá)載體
將帶酶切位點(diǎn)的 Lj-pcna目的片段和pGFP-N2質(zhì)粒均用HindⅢ和BamHⅠ雙酶切,切膠回收后分別用10 mg/L的瓊脂糖凝膠鑒定,見圖 4。用 T4連接酶將切膠回收后的目的片段Lj-pcna與 pGFP-N2載體連接,轉(zhuǎn)化,提取pGFP-N2-Lj-pcna重組質(zhì)粒。
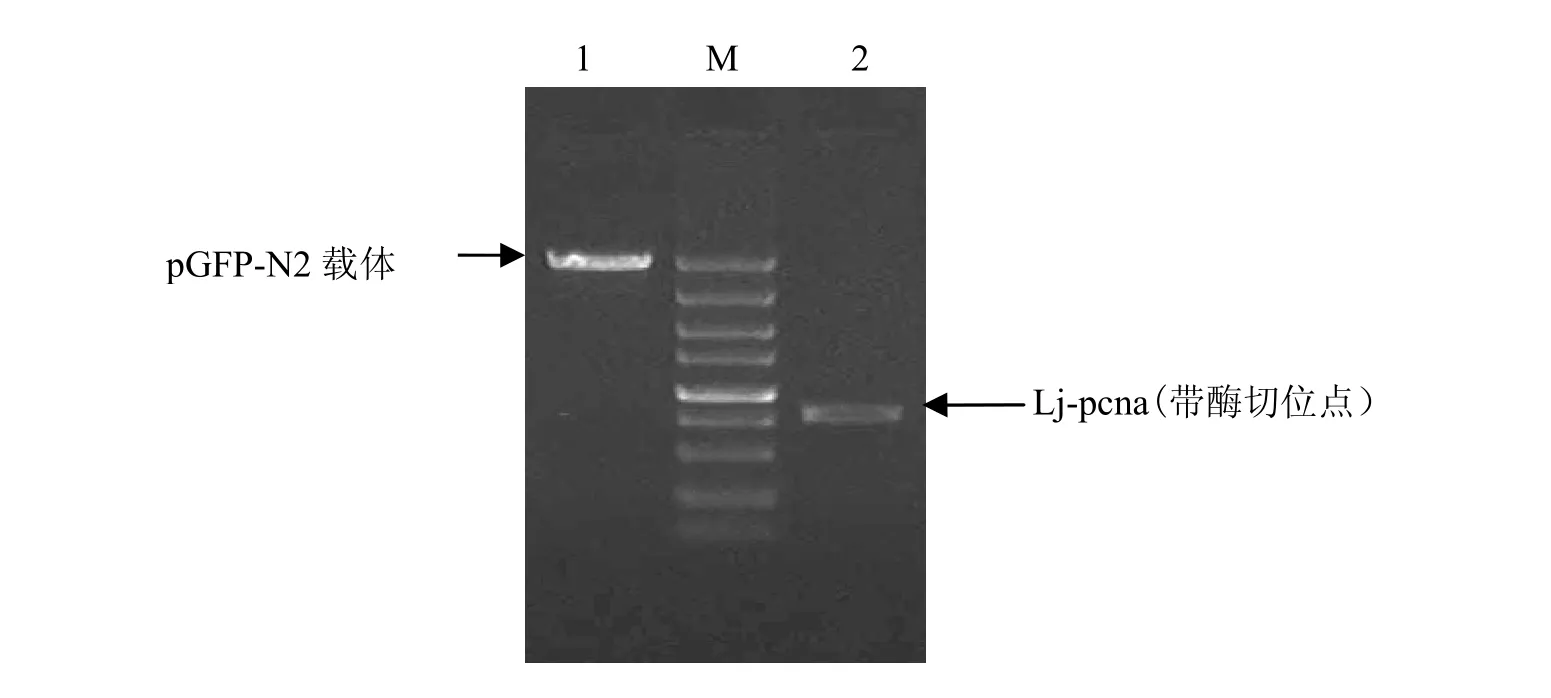
圖4 pGFP-N2載體及Lj-pcna酶切片段回收
2.4 真核表達(dá)載體鑒定
pGFP-N2-Lj-pcna重組質(zhì)粒經(jīng)HindⅢ和BamHⅠ雙酶切及PCR擴(kuò)增均得到約780bp的目的片段,見圖5,測(cè)序結(jié)果顯示插入載體中的pcna核苷酸序列沒有發(fā)生突變,與cDNA文庫中EST序列完全一致,說明成功將 pcna基因構(gòu)建到pGFP-N2真核表達(dá)載體中。

圖5 真核表達(dá)載體pGFP-N2-Lj-pcna鑒定
2.5 pGFP-N2-Lj-pcna在Hela細(xì)胞中表達(dá)
在12-well plate 中接種 Hela 細(xì)胞,細(xì)胞接種后 24 h,使用 Xfect Transfection Reagent將pGFP-N2-Lj-pcna質(zhì)粒轉(zhuǎn)染到Hela細(xì)胞中,細(xì)胞轉(zhuǎn)染24 h后,熒光顯微鏡下觀察有綠色熒光蛋白表達(dá),見圖6。

圖6 融合蛋白(七鰓鰻PCNA和綠色熒光蛋白)在Hela細(xì)胞中表達(dá)(×100)
3 討論
pcna又稱為周期蛋白(Cyclin),是DNA多聚酶 δ 的一種輔助因子。靜止細(xì)胞含量很少,G1晚期開始增多,S期達(dá)到高峰,G2期、M期下降,其量的變化與DNA合成一致,腫瘤細(xì)胞增殖越活躍,pcna表達(dá)越高,是評(píng)價(jià)細(xì)胞增殖狀態(tài)的重要指標(biāo)[11-14]。七鰓鰻是現(xiàn)存最原始的無頜類脊椎動(dòng)物,它印記了無脊椎動(dòng)物的進(jìn)化歷史,同時(shí)作為脊椎動(dòng)物最直接的祖先,又為脊椎動(dòng)物的起源與進(jìn)化提供了豐富的遺傳信息。七鰓鰻作為脊椎動(dòng)物發(fā)育早期和原始特征的模型,是研究哺乳動(dòng)物基因起源和進(jìn)化十分難得的實(shí)驗(yàn)材料。前期研究發(fā)現(xiàn),體外培養(yǎng)的七鰓鰻細(xì)胞分裂緩慢,細(xì)胞生長周期長,不利于體外七鰓鰻組織細(xì)胞的原代培養(yǎng),更難以建立七鰓鰻細(xì)胞系[15]。因此促使我們從cDNA文庫的EST序列中尋找促進(jìn)細(xì)胞生長的一些周期蛋白,其中pcna基因就是一個(gè)重要的細(xì)胞周期蛋白。目前,還沒有關(guān)于七鰓鰻pcna基因序列與功能的相關(guān)報(bào)道。本課題組首次從日本七鰓鰻肝臟組織中克隆了與高等脊椎動(dòng)物 pcna同源的cDNA序列。NCBI數(shù)據(jù)庫的BLAST搜索分析表明,七鰓鰻 pcna的核苷酸及推導(dǎo)的氨基酸序列和其他物種的 pcna有很高的同源性。從進(jìn)化樹結(jié)果分析,Lj-pcna位于高等脊椎動(dòng)物pcna基因的外群。因此可通過對(duì)日本七鰓鰻(現(xiàn)存最古老的脊椎動(dòng)物)pcna基因的研究,從而確定七鰓鰻與其它物種的進(jìn)化地位及聯(lián)系。
目前用于表達(dá)外源性基因的真核表達(dá)載體主要分為病毒載體和非病毒載體兩大類。一般而言,病毒載體相對(duì)表達(dá)穩(wěn)定,轉(zhuǎn)染效率高,不過易隨機(jī)整合到靶細(xì)胞的染色體上,故難以控制且安全性差。非病毒載體主要是質(zhì)粒載體,質(zhì)粒載體被轉(zhuǎn)染試劑轉(zhuǎn)染入靶細(xì)胞后,獨(dú)立存在于靶細(xì)胞中,對(duì)靶細(xì)胞自身傷害小,安全性較好,因此本實(shí)驗(yàn)采用質(zhì)粒載體作為目的基因的攜帶者,將目的基因轉(zhuǎn)入靶細(xì)胞中。近幾年來,報(bào)告基因已普遍應(yīng)用在細(xì)胞生物學(xué)和分子生物學(xué)等研究領(lǐng)域[16],現(xiàn)有報(bào)告基因種類很多,如半乳糖苷酶(LacZ)、葡萄糖苷酸酶(GUS)、螢火蟲熒光素酶(LUC)等,此外,綠色熒光蛋白(Green Fluorescent Protein,GFP)是一類較理想的報(bào)告基因,其在基礎(chǔ)研究領(lǐng)域及臨床檢測(cè)中都發(fā)揮著重要作用,將帶有GFP報(bào)告基因的載體轉(zhuǎn)入靶細(xì)胞后,可以很方便地觀察目的蛋白的定位及確定目的蛋白在細(xì)胞或體內(nèi)的表達(dá)情況,并且在動(dòng)物實(shí)驗(yàn)中通過熒光顯微鏡可以對(duì)表達(dá) GFP的細(xì)胞進(jìn)行定位觀察。目前科研領(lǐng)域應(yīng)用最廣泛的是將GFP同目的蛋白形成融合蛋白,通過GFP熒光顯示位置從而監(jiān)測(cè)目的蛋白的定位[17],由于GFP作為報(bào)告基因具有方便、及時(shí)、直觀、應(yīng)用廣泛等特點(diǎn),因而在基因表達(dá)、基因整合、細(xì)胞分選及轉(zhuǎn)基因動(dòng)物研究領(lǐng)域的應(yīng)用是其它報(bào)告基因所無法取代的[18.19]。因此本實(shí)驗(yàn)采用分子克隆技術(shù),將七鰓鰻pcna基因構(gòu)建到pGFP-N2真核表達(dá)載體上,經(jīng)限制性酶切,PCR擴(kuò)增及DNA測(cè)序等證明載體構(gòu)建成功。另外應(yīng)用一種新型的轉(zhuǎn)染試劑——Xfect,一種用于胚胎干細(xì)胞的轉(zhuǎn)染試劑對(duì)重組質(zhì)粒進(jìn)行轉(zhuǎn)染。Xfect轉(zhuǎn)染試劑兼容性很好,無論質(zhì)粒質(zhì)量好壞,也無論質(zhì)粒是環(huán)狀還是線性,均能獲得較高的轉(zhuǎn)染效率。本實(shí)驗(yàn)結(jié)果表明pcna蛋白與綠色熒光蛋白以融合蛋白形式表達(dá)在Hela細(xì)胞內(nèi),且表達(dá)效率較高。
本研究成功將七鰓鰻 pcna基因構(gòu)建到真核表達(dá)載體 pGFP-N2上,并在 Hela細(xì)胞中成功地表達(dá)了PCNA和GFP融合蛋白,說明我們構(gòu)建的針對(duì) pcna的真核表達(dá)載體是正確有效的,為后續(xù)研究 pcna基因在七鰓鰻中功能研究以及同其它基因之間的相互關(guān)系提供實(shí)驗(yàn)工具,并為今后能夠建立穩(wěn)定的七鰓鰻細(xì)胞株奠定實(shí)驗(yàn)基礎(chǔ)。
[1]HERRIN B R, COOPER M D. Alternative adaptive immunity in jawless vertebrates [J]. J Immunol, 2010,185(3): 1367-1374.
[2]GUO P, HIRANO M, HERRIN B R, et al. Dual nature of the adaptive immune system in lampreys[J]. Nature,2009, 459(7248): 796-801.
[3]YUE Pang, RONG Xiao, XIN Liu, et al. Identification and characterization of the lamprey high-mobility group box 1 gene[J]. Plos one, 2012, 7(4): e35755.
[4]MAGA G, HUBSCHER U. Proliferating cell nuclear antigen (PCNA): a dancer with many partners[J]. J Cell Sci, 2003, 116(15): 3051-3060.
[5]OSADA S, SAJI S, KUNO T. Clinical significance of combination study of apoptotic factors and proliferating cell nuclear antigen in estimating the prognosis of hepatocellular carcinoma [J]. J Surg Oncol, 2004, 85(1):48-54.
[6]BRAVO R, FRANK R, BLUNDELL P A,Macdonald-Bravo H. Cyclin/PCNA is the auxiliary protein of DNA polymerase delta[J]. Nature, 1987,326(6112):515-517.
[7]STENNER M, DEMGENSKY A, MOLLS C, et al.Prognostic value of proliferating cell nuclear antigen in parotid gland cancer [J]. Eur Arch Otorhinolaryngol,2012, 269(4): 1225-1232.
[8]ACHARYA N, KLASSEN R, JOHNSON R E, et al.PCNA binding domains in all three subunits of yeast DNA polymerase δ modulate its function in DNA replication[J]. Proc Natl Acad Sci USA, 2011,8(44):17927-17932.
[9]STRZALKA W, ZIEMIENOWICZ A. Proliferating cell nuclear antigen(PCNA):a key factor in DNA replication and cell cycle regulation[J]. Ann Bot, 2011, 107(7):1127-1140.
[10]朱麗娜, 戴雅麗, 馬飛, 等. 日本七鰓鰻(Lampetra japonica)肝臟 ESTs分析與比較轉(zhuǎn)錄組研究[J]. 中國科學(xué)C輯:生命科學(xué), 2007(6): 609-619.
[11]ZHAO H, LO YH, MA L, et al. Targeting tyrosine phosphorylation of PCNA inhibits prostate cancer growth[J]. Mol Cancer Ther, 2011, 10(1):29-36.
[12]CHEN J, AI Y, WANG J, et al. Chemically ubiquitylated PCNA as a probe for eukaryotic translesion DNA synthesis[J]. Nature Chemical Biology,2010, 6(4): 270-272.
[13]MAYANAGIA K,KIYONARIC S, SAITOD M, et al.Mechanism of replication machinery as sembly as revealed by the DNA ligase-PCNA-DNA complex architecture[J]. PNAS, 2009, 106(12): 4647-4652.
[14]ROSENTAL B, HADAD U, BRUSILOVSKY M, et al.A novel mechanism for cancer cells to evade immune attack by NK cells: The interaction between NKp44 and proliferating cell nuclear antigen [J]. Oncoimmunology,2012, 1(4): 572-574.
[15]逄越, 李慶偉. 日本七鰓鰻外周血細(xì)胞顯微結(jié)構(gòu)及類淋巴細(xì)胞體外培養(yǎng)[J]. 海洋科學(xué), 2012, 36(1):23-29.
[16]張峰, 任燕, 陸長得. 綠色熒光蛋白及其應(yīng)用[J]. 生命科學(xué), 1999, 11(2): 61-65.
[17]MARX J C, ALLAY J A, PERSONS D A, et al. High efficiency transduction and long-term gene ex pr ession with a murine stem cell retroviral vector encoding the green fluorescent protein in human marrow stromal cells [J]. Hum Gene Ther, 1999, 10(7): 1163-1173.
[18]OHMIYA N, EMI N, NIWA Y, et al. Insulin-enhanced liposome-mediated gene transfer into a gastric carcinoma cell line [J]. Clin Exp Pharmacol Physiol,2002, 29(7): 544- 548.
[19]PERSONS D A, ALLAY J A, ALLAY E R, et al.Retroviral mediated transfer of the green fluorescent protein gene into murine hematopoietic cells facilitates scoring and selection of transduced progenitors in vitro and identification of genetically modified cells in vivo[J]. Blood, 1997, 90(5): 1777-86.
