青花菜非對稱體細胞雜交研究
2012-10-10 06:47:02戴永娟和兆榮胡靖鋒楊紅麗刁玉華李春慧和江明
湖南農業科學 2012年5期
關鍵詞:融合
戴永娟 ,和兆榮 ,胡靖鋒 ,汪 騫 ,楊紅麗 ,刁玉華 ,李春慧 ,和江明
(1.云南大學生命科學學院,云南 昆明 650091;2.云南省農業科學院,云南 昆明 650205;3.通海縣經濟作物工作站,云南 通海 652700)
青花菜(Brassica oleracea var.italica),又稱西蘭花,屬十字花科(Cruciferae)蕓薹屬(Brassica L.)植物,食用部分為花球[1]。由于其柔嫩多汁、味道鮮美以及具有防癌抗癌等功效深受廣大消費者的喜愛。但青花菜在生產中常常遭受病蟲害的威脅,嚴重影響了產量和品質。青花菜新品種“優秀”是日本坂田公司育成的春秋兩用的早熟青花菜新品種,具有早熟、長勢旺、抗性強、花蕾細小濃綠、花球平滑、出口合格率高等特點,是目前比較理想的早熟型青花菜品種。但“優秀”是一個Ogura蘿卜胞質雄性不育品種,不育性、不育率和不育度均為100%,利用常規雜交育種難以實現其核基因所控制的優異性狀的轉移。然而體細胞融合技術是實現這一轉移的有效途徑[2]。
自1960年Cocking[3]首創原生質體技術,20世紀70年代中期Kartha等[4]通過葉肉細胞形成愈傷組織和再生植株,Schenck[5]第一次報道成功獲得B.oleracea和B.napa的體細胞雜種植株至今,蕓薹屬作物幾乎所有種都可由原生質體培養獲得再生植株[6]。在蕓薹屬內、蕓薹屬與近緣種之間有不少獲得體細胞雜種的報道,同時非對稱融合技術在蕓薹屬內也得到了快速發展及應用。非對稱體細胞融合能夠克服有性雜交的障礙,解決同源染色體的配對問題,實現核質和胞質的重新組合[7-8],而且形成的雜種通常只含有供體親本的部分染色體,因此在提高雜種的育性及縮短育種年限等方面具有一定的優勢。近年來國內外不少專家學者通過非對稱體細胞雜交已經成功獲得雜種植株[9-11]。筆者旨在通過非對稱體細胞雜交途徑,將“玉皇”可育的細胞質轉移到“優秀”中,以期探索青花菜進行性狀改良的途徑,為實現品種改良提供可用的育種材料。
1 材料和方法
1.1 材料
“玉皇”為供體,“優秀”為受體(日本坂田公司)。
1.2 方法
1.2.1 原生質體制備 青花菜原生質體分離及純化:分別取接種于MS培養基中光照培養5~6 d的“玉皇”、暗培養的“優秀”無菌苗下胚軸,碎解成1 mm左右的小段,按50粒種子1 mL酶液,于25℃黑暗條件下,搖床上30 r/min酶解12 h,酶液的配制參考Kao等[12]。酶解后用 45 μm孔尼龍網過濾酶解分離原生質體。將濾液移至離心管中,1 000 r/min離心5 min,棄上清,加入CPW-10洗液(27.2 mg/L KH2PO4、101 mg/L KNO3、246 mg/L MgSO4·7H2O、0.16 mg/L KI、0.025 mg/L CuSO4·5H2O、1 480 mg/L CaCl2·2H2O、10% 甘露醇,pH 5.8),離心清洗5 min,棄上清,重復2次。最后用原生質體培養基(PJ1)離心清洗1次,即可獲得純凈的青花菜原生質體。將收集到的“優秀”,“玉皇”原生質體調濃度至(1~2)×106個/mL,分別用 0.05、0.075、0.10、0.15、0.20、0.25 J/cm劑量的UV射線照射“玉皇”下胚軸原生質體,然后分別用 10、20、30、40、50、60 μg/mL的羅丹明(R-6G)處理“優秀”下胚軸原生質體。處理方法參考侯喜林等[13]。
1.2.2 原生質體活力測定 伊文思藍不為活原生質體吸收,而為死原生質體吸收,顯藍色的原生質體為無活力的。取0.5 mL純化的原生質體,加入1~2滴0.3%的伊文思藍染色液,染色2~3 min后,顯微鏡下觀察,統計其存活率。重復3次,取其平均值進行統計。
存活率=(未染色的原生質體/觀察的原生質體總數)×100%
1.2.3 原生質體的融合及培養 (1)正交試驗優化培養基激素水平:選用6-BA、NAA、2,4-D激素濃度3個因素進行L9(34)正交試驗,優化原生質體培養基激素水平(表1)。
(2)供、受體原生質體融合:將經過處理的供、受體原生質體,按1:1的比例混合(各約0.5 mL)。融合方法采用聚乙二醇(PEG)融合法[14],略作修改。加入PEG溶液0.425 mL(PEG-6000 250 g/L、Ca-Cl2·2H2O 1.025 g/L、甘露醇 45.5 g/L、山梨醇 45.5 g/L,pH 6.0)。然后再加入15%二甲基亞砜(DMSO)0.075 mL,室溫下靜置融合10 min。加入PEG洗液(PEG洗液A:甘氨酸7.5 g/L,pH 10.5;PEG洗液B:CaCl2·2H2O 14.7 g/L、甘露醇 91 g/L、山梨醇 91 g/L,pH 10.5),PEG 洗液 A、B 使用前按 1∶1 立即混勻。置30℃培養箱靜置30 min,然后小心移除上清,加入 50 mmol/L CaCl2·2H2O,再次清洗 PEG,移除上清,用培養基(PJ1)清洗1次,最后每個培養皿加入2 mL(PJ1)進行融合原生質體的培養。
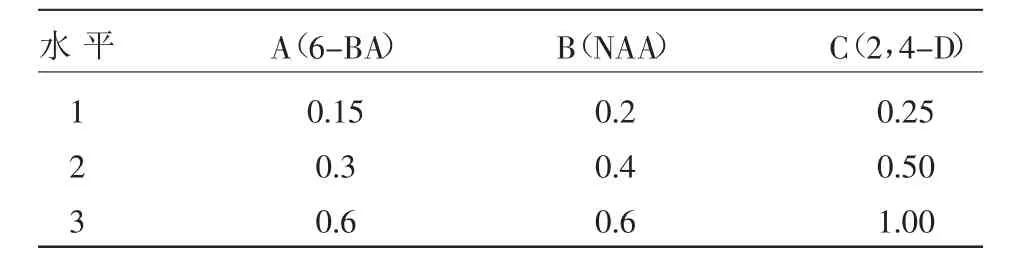
表1 原生質體培養基因素水平 (mg/L)
(3)融合體培養:25℃暗培養5~7 d,當觀察到好的細胞分裂時,加入 0.5 mL 培養基(PJ2),3~5 d后,當觀察到有小細胞團形成時,再加入0.5 mL培養基(PJ3),同時光照轉為正常光照。此后每隔3 d加入1次培養基(PJ3),促使愈傷組織形成。同時以相同激素水平的培養基培養經UV處理過的“玉皇”原生質體,和經R-6G處理過的“優秀”原生質體作為對照。
1.2.4 愈傷組織分化與植株再生 當愈傷組織長到約5 mm大小時,將其轉移至擴增培養基(PJ4),弱散射光下培養,促進芽生長。當幼苗的真葉形成時轉移到無激素MS培養基中使再生植株正常生長。培養20 d左右,將正常生長植株轉入生根培養基(PJ5)。原生質體培養、稀釋、擴增和生根培養基的激素配比見表2。
2 結果與分析
2.1 UV射線對“玉皇”原生質體核失活的影響
從圖1中可以看出,所有UV射線都會使原生質體的活力下降,并隨著輻射劑量的加大,活力下降增多。與對照相比,除0.050 J/cm的處理外,其他處理劑量,原生質體活力下降都超過了50%,說明青花菜原生質體對UV射線處理較為敏感。對上述所設UV射線劑量處理的原生質體進行培養,培養2 d后,0.050~0.075 J/cm輻射的處理,原生質體均在培養中有細胞分裂,3~5 d開始第一次細胞分裂,培養13 d左右,倒置顯微鏡下可觀察到細胞團,培養25 d左右可以發現肉眼可見愈傷組織或胚狀體,但隨著輻射劑量的加大,植板率明顯下降。輻射劑量為0.15 J/cm,僅在培養中發現了少數幾次細胞分裂,未見有細胞團形成。高于0.15 J/cm輻射處理的均未發現細胞分裂以及細胞團和胚狀體的形成,試驗采用劑量為0.10 J/cm。
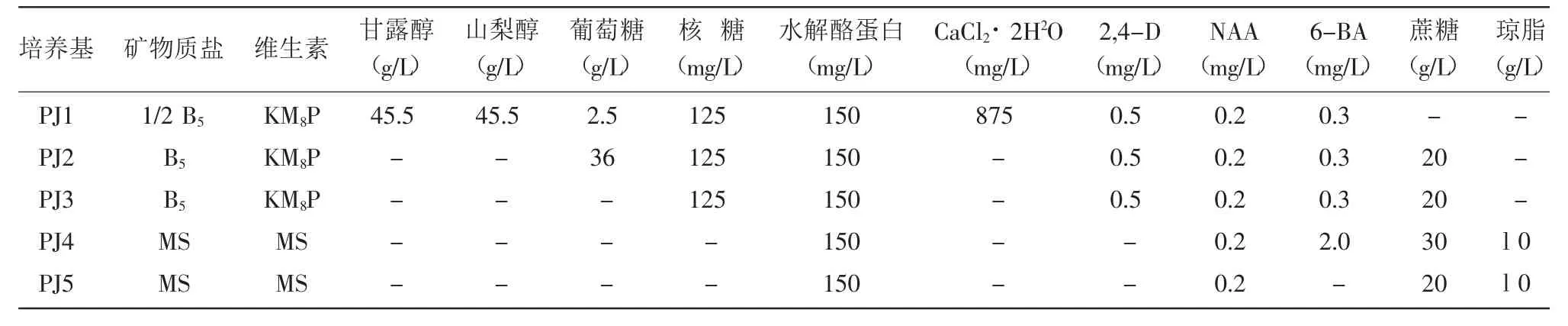
表2 培養基的激素配比
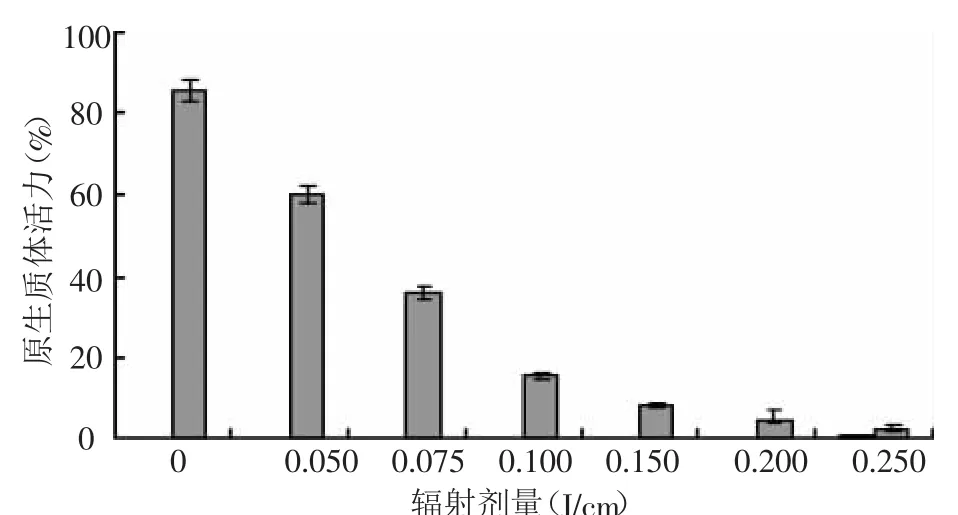
圖1 UV射線對受體原生質體活力的影響
2.2 羅丹明(R-6G)對“優秀”原生質體失活的影響
如圖2所示,羅丹明(R-6G)在10 μg/mL濃度下處理30 min(包括離心所用時間,25℃),“優秀”原生質體的植板率為14.6%;當濃度加倍為20 μg/mL時,植版率下降為6.7%;當濃度為50 μg/mL時,植板率為0。試驗采用濃度為40 μg/mL,處理30 min。
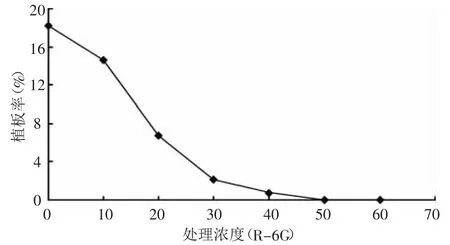
圖2 羅丹明(R-6G)對受體原生質體活力的影響
2.3 融合方法對原生質體融合率的影響
對比了姜淑慧等[14](方法1)、Ryshka[15](方法2)以及兩種方法結和(方法3)對融合率的影響。方法1、2、3原生質體的融合率分別為 9%、12%、14%,方法3的融合效果要高于方法1和方法2。方法3的洗滌液中,不僅有Ca2+能夠保證原生質體的質膜穩定,而且添加甘露醇能夠保證穩定的原生質體滲透壓。30℃培養箱中洗滌30 min,可以使原生質體膜融合更加充分。
2.4 不同激素組合對青花菜下胚軸原生質體培養的影響
采用正交試驗,研究了不同激素組合對青花菜下胚軸原生質體培養的影響,結果如表3所示。對9個試驗結果進行比較可以看出,組合4分裂頻率最高,為 18.9%,為 A2、B1、C2。從表 3 中可以看出,極差值RA>RC>RB,說明6-BA激素水平變化對分裂頻率的影響高于NAA、2,4-D。
表3 激素組合對原生質體分裂頻率的影響
2.5 不同培養基對融合原生質體培養的影響
利用Km8p[16]改良的B5[17]以及筆者綜合改良的培養基(PJ),3種培養基進行融合體培養的比較。結果表明同一激素水平下,筆者所采用的培養基融合體分裂頻率相對較高,細胞團出現的時間相對較早,在進一步培養的過程中褐化程度相對較低(表4)。

表4 基本培養基對融合體的影響
2.6 融合體愈傷組織、胚狀體的形成及植株再生
由于供體“玉皇”采用的是正常光照下培養的幼苗下胚軸作為原生質體分離材料,“優秀”采用的是暗培養的幼苗下胚軸,原生質體中不含有葉綠體。通過顯微鏡觀察,結果如圖3所示。融合體中含有不含葉綠體的融合,圖3A為分離、純化的原生質體;圖3B說明供受體之間已發生融合;在融合體培養3~4 d發現細胞第一次分裂,6~7 d第二次分裂,12 d左右就可以觀察到分裂形成的細胞團(圖3C);18 d左右可以觀察到胚體雛形(圖 3D);25~30 d形成肉眼可見的愈傷組織或胚狀體(圖3E);當愈傷組織長到約5 mm時,轉移愈傷組織到(PJ4)培養基促進芽生長;當幼苗的真葉形成時轉移到無激素MS培養基中使再生植株正常生長,再將正常生長植株轉入生根培養基中,促進再生植株生根(圖3F、圖 3G)。
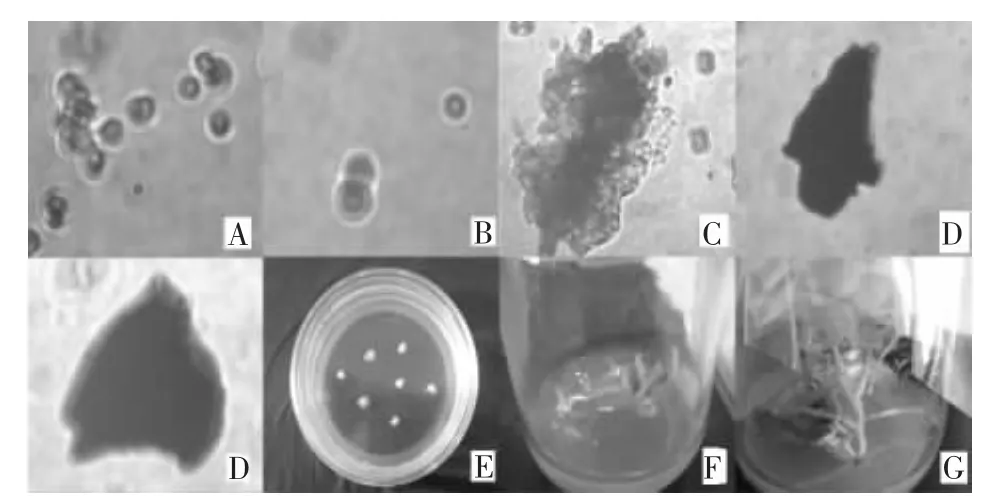
圖3 “優秀”、“玉皇”原生質體融合及再生植株
3 討 論
筆者利用綜合改進的PEG融合法將青花菜“玉皇”、“優秀”的原生質體進行融合,該方法明顯提高了融合率。因為2種原生質體加入PEG融合液后,只發生粘連,在洗滌過程中才發生膜融合,核融合通常在第一次有絲分裂過程中發生[18],所以在加入PEG溶液使原生質體發生粘連后,洗滌過程才是是否發生融合的關鍵。
Km8p、B5是廣泛運用于原生質體培養的培養基。近年來,在原生質體培養中也有很多在Km8p、B5的基礎上通過改良得來的培養基[19]。筆者結合了Km8p的有機成分、B5的礦物質鹽,利用甘露醇和山梨醇作為滲透壓調節劑,以綜合改進的培養基進行體細胞融合培養,進一步減少了培養過程中的褐化程度,提高分裂頻率。在融合體培養過程中采用不斷減小培養基滲透壓的方法,減少了酚類物質對細胞的毒害作用。供受體材料分別用UV射線和羅丹明(R-6G)進行預處理,但處理劑量及濃度均小于原生質體失活的量,因為在處理過程中,細胞活力均會受到很大的影響,若均采用鈍化劑量進行融合及培養,將會極大程度降低植板率。經過預處理作為對照培養的供、受體原生質體,在培養過程中很少發現有細胞分裂,未見有細胞團形成,繼續培養1個月以后也未見愈傷組織或胚狀體。由此再次說明,UV射線和羅丹明(R-6G)的處理抑制了供、受體原生質體的分裂和生長,融合后培養再生獲得的愈傷組織和胚狀體來源于融合細胞,再生植株能初步判定為雜種植株。
筆者在建立青花菜原生質體融合及培養的新體系的同時,還探究了原生質體融合、培養影響因素,獲得了再生植株,并在原生質體融合初期及愈傷組織和胚狀體形成時期進行了初步的雜種鑒定,為進一步分子細胞遺傳研究,生物學鑒定工作提供了科學依據。
[1]陸信娟,楊 峰,樊繼德.青花菜主要農藝性狀的相關分析[J].江西農業學報,2011,23(1):49-51.
[2]惠志明.原生質體非對稱融合向花椰菜轉移Ougar蘿卜胞質雄性不育性的研究[D].保定:河北農業大學,2002.
[3]Cocking E C.A method for the isolation of plant protoplast and Vacuoles[J].Nature,1960,187:962-963.
[4]Kartha K K,Michayluk M R,Kao K N,et al.Callus formation and plant regeneration form mesophyll protoplasts of rape plants(Brassica napus L.CV.Zphyr)[J].Plant science,1974,3(4):265-271.
[5]Schenck H R,Rbbelen G.Somatic hybrids by fusion protoplast form Brassica Oleracea and B.campestris [J].Can J Genet planzenz,1982,89:278-288.
[6]俞志華.蕓薹屬作物體細胞雜交研究[J].世界農業,1992,(5):17-19.
[7]叢建民,沈海龍,李玉花,等.植物體與植物體細胞無性系發育中的DNA甲基化及其表達調控研究進展 [J].安徽農業科學,2011,39(17):10170-10174,10196.
[8]Xie R,Yang Z L,Liu C Y,et al.RITSD analysis of the agronomic traits of somaclonal in rice[J].Agricultural Science&Technology,2010,11(1):36-41.
[9]周愛芬,夏光敏,陳惠民.普通小麥與簇毛麥不對稱體細胞雜交的研究[J].中國科學,1996,26(1):31-37.
[10]Sjodin C,Glimelius K.Transfer of disease resistance within the genus Brassica through asymmetric somatic hybridization[J].Theor Appl Genet,1989,78:513-520.
[11]姚星偉,劉 凡,云興福,等.非對稱體細胞融合獲得花椰菜與Brassica spinescens的種間雜種 [J].園藝學報,2005,32:1039-1044.
[12]Kao C M,.Keller W A,Gleddie S,et al.Efficent plant regeneration form hypocotyls protoplast of broccoli(Brassica oleracea L.ssp.italica Plenk)[J].Plant Cell Reports,1990,(9):311-315.
[13]侯喜林,曹壽椿,佘建明,等.碘乙酰胺和羅丹明對不接球白菜子葉原生質體線粒體失活效果的影響 [J].中國蔬菜,2002,(4):18-19.
[14]姜淑慧,管榮展,董海濱,等.播娘篙與甘藍型油菜原生質體融合與植株再生[J].中國油料作物學報,2005,27(4):1-6.
[15]Ryshka U,Schumann G,Klocke E,et al.Somatic hybridization in Brassicaceae[J].Acta Hortic,1996,407:201-208.
[16]Kao K N,Michayluk M R.Nutrtional requirements for growth Vica hajastana cells and protoplast at a very low population density in liquid media[J].Planta,1975,126(2):105-110.
[17]Gamborg O L,Miuer R A,Ojima K.Naturient requirements of suspension cultures of soybean root cells[J].Exp cell Res,1968,50:151-158.
[18]梁 丹,丁 丹,王火旭.甘藍與大白菜的原生質體融合[J].安徽農業科學,2009,37(8):3448-3449.
[19]侯喜林,曹壽椿,佘建明,等.不接球白菜原生質體培養再生植株[J].南京農業大學學報,2000,23(4):17-20.
猜你喜歡
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
數學年刊A輯(中文版)(2022年4期)2022-02-16 08:17:34
今日農業(2021年19期)2022-01-12 06:16:36
中老年保健(2021年11期)2021-08-22 03:15:44
無線電通信技術(2021年4期)2021-07-13 08:58:28
無線電通信技術(2021年3期)2021-06-08 03:33:48
中學生數理化(高中版.高考數學)(2021年1期)2021-03-19 08:28:38
無線電工程(2020年11期)2020-10-29 01:25:46
現代出版(2020年3期)2020-06-20 07:10:34
福利中國(2015年4期)2015-01-03 08:03:38
